Рыбья чешуя - Википедия - Fish scale

А рыбья чешуя представляет собой небольшую жесткую пластину, которая растет из кожа рыбы. Кожа большинства рыб покрыта этими защитными напольные весы, который также может обеспечить эффективную камуфляж за счет использования отражение и окраска, а также возможные гидродинамические преимущества. Период, термин шкала происходит от Старофранцузский «escale», что означает стручок или шелуха.[1]
Чешуя сильно различается по размеру, форме, структуре и размеру, начиная от прочных и жестких пластин брони у таких рыб, как креветки и самшит, микроскопическим или отсутствует у рыб, таких как угри и удильщик. В морфология шкалы можно использовать для определения вида рыб, из которых она произошла.
Наиболее костлявые рыбы покрыты циклоидной чешуей лосось и карп, или ктеноидные чешуи окунь, или чешуйки ганоидов осетровые и Гарс. Хрящевые рыбы (акулы и лучи ) покрыты плакоидной чешуей. Вместо этого некоторые виды покрываются щитки, и другие не имеют внешнего покрытия на части или на всей коже.
Рыбья чешуя является частью рыбной покровная система, и производятся из мезодерма слой дерма, что отличает их от чешуя рептилий.[2][3] Такой же гены участвует в развитии зубов и волос в млекопитающие также участвуют в масштабном развитии. Плакоидные чешуйки хрящевых рыб также называются дермальными зубчиками и структурно являются гомологичный с зубами позвоночных. Было высказано предположение, что чешуя костистых рыб похожа по структуре на зубы, но, вероятно, происходит из другой ткани.[4] Большинство рыб также покрыто слоем слизь или слизь, которая может защитить от патогенов, таких как бактерии, грибки и вирусы, и снизить поверхностное сопротивление, когда рыба плавает.
Весы телодонта

Костные чешуи телодонты, самая распространенная форма ископаемая рыба, хорошо поняты. Чешуя формировалась и сбрасывалась на протяжении всей жизни организмов и быстро отделялась после их смерти.[5]
Кость - ткань, которая устойчива к механическим повреждениям и относительно склонна к окаменению, часто сохраняет внутренние детали, что позволяет гистология и рост чешуек подлежит детальному изучению. Чешуя представляет собой нерастущую «корону», состоящую из дентин, с иногда украшенным эмелоид верхняя поверхность и аспидиновая основа.[6] Его растущая основа сделана из бесклеточной кости, которая иногда развивает структуры крепления, чтобы зафиксировать ее на боку рыбы.[7] Помимо этого, по-видимому, существует пять типов роста костей, которые могут представлять пять естественных групп внутри телодонтов - или спектр, варьирующийся между конечными членами мета- (или орто) дентина и мезодентиновыми тканями.[8] Кажется, что каждая из пяти морфов чешуи напоминает чешую более производных групп рыб, что позволяет предположить, что группы телодонтов могли быть стволовыми группами для последующих кладов рыб.[7]
Однако использование одной лишь морфологии чешуи для различения видов имеет некоторые подводные камни. Внутри каждого организма форма чешуи сильно различается в зависимости от площади тела.[9] с промежуточными формами, появляющимися между различными областями - и, что еще хуже, морфология чешуи может даже не быть постоянной в пределах одной области. Чтобы еще больше запутать ситуацию, морфология чешуи не уникальна для таксонов и может быть неотличима на одной территории у двух разных видов.[10]
Морфология и гистология телодонтов являются основным инструментом для количественной оценки их разнообразия и различения видов, хотя в конечном итоге с использованием таких сходящийся traits подвержен ошибкам. Тем не менее, структура, состоящая из трех групп, была предложена на основе морфологии и гистологии чешуи.[8] Сравнение с современными видами акул показало, что чешуя телодонта была функционально схожа с чешуей современных хрящевых рыб, а также позволило провести обширное сравнение экологических ниш.[11]
Космоидные весы

Космоидные чешуи встречаются только на древних рыба с лопастными плавниками, в том числе некоторые из самых ранних двоякодышащие (подкласс Двоякодышащие ), И в Кроссоптеригии, в том числе живые латимерия в модифицированном виде (см. эластоидные шкалы ниже). Вероятно, они произошли от слияния чешуек плакоидов. Внутренняя часть чешуи выполнена из плотной пластинчатый кость называется изопедином. Сверху лежит слой губчатой или сосудистый кость снабжена кровеносными сосудами, за которой следует комплекс дентин -подобный слой, называемый космин с поверхностным внешним покрытием из витродентин. Верхняя поверхность кератин. Космоидные чешуйки увеличиваются в размерах за счет роста пластинчатого слоя кости.[12]
Элазмоидная чешуя

Элазмоидные чешуйки тонкие, черепичный чешуйки, состоящие из слоя плотной пластинчатой коллагеновой кости, называемой изопедином, над которой находится слой бугорков, обычно состоящих из кости, как в Евстеноптерон. Слой дентина, который присутствовал у рыб с первыми плавниками, обычно редуцирован, как и у современных латимерия, или полностью отсутствует, как в двоякодышащая рыба и в девоне Евстеноптерон.[13] Элазмоидная чешуя на протяжении эволюции рыб появлялась несколько раз. Они присутствуют в некоторых рыба с лопастными плавниками, такие как все существующие и некоторые вымершие двоякодышащие, так же хорошо как латимерия которые имеют видоизмененные космоидные чешуйки, в которых отсутствует космин, и которые тоньше истинных космоидных чешуек. Они также присутствуют у некоторых тетраподоморфов, таких как Евстеноптерон, амииды и костистые кости, циклоидная и ктеноидная чешуйки которых представляют наименее минерализованные эластоидные чешуйки.
В данио Элазмоидные чешуйки используются в лаборатории для изучения процесса минерализации костей и могут культивироваться (храниться) вне организма.[14][15]
Ганоидная чешуя


Чешуя ганоида встречается в осетровые, веслонос, Гарс, Bowfin, и бихиры. Они происходят от космоидных чешуек и часто имеют зазубренные края. Они покрыты слоем твердой эмали. дентин в месте космин, и слой неорганической костной соли, называемый ганоин на месте витродентин.
Ганоин - характерный компонент чешуи ганоида. Это стекловидная, часто многослойная минерализованная ткань покрывающий весы, а также черепной кости и плавниковые лучи в некоторых нетипичных рыбы с луговыми плавниками,[16] Такие как Гарс, бихиры, и латимерия.[17][18] Он состоит из стержневидных апатит кристаллиты.[19] Ганойн - это древняя особенность рыб с лучевыми плавниками, встречающаяся, например, на чешуе стволовая группа актиноптеригиан Хейролепис.[18] Хотя часто считается синапоморфный характер лучеплавниковых рыб, ганоин или ганоиноподобные ткани также встречаются на вымерших акантодии.[18] Было высказано предположение, что ганоин гомологичный к эмаль зубов у позвоночных[16] или даже считается разновидностью эмали.[19]
 Стриатус амблиптер | Ганоид чешуи вымерших Каменноугольный рыбы, Стриатус амблиптер. (а) показывает внешнюю поверхность четырех чешуек, а (б) показывает внутреннюю поверхность двух чешуек. Каждая из ганоидных чешуек ромбовидной формы у Amblypterus имеет гребень на внутренней поверхности, который на одном конце превращается в выступающий штифт, который входит в выемку в следующем масштабе, подобно тому, как черепица соединяется вместе на крыше. дома. |  |
Большинство чешуек ганоидов ромбовидный (ромбовидные) и соединяются шпильками. Обычно они толстые и сложены друг с другом, как мозаика, а не накладываются друг на друга, как другие чешуйки.[20] Таким образом, чешуя ганоида почти непроницаема и является отличной защитой от хищников.

В аллигатор гар имеет жесткую броню ромбовидный -образная чешуя ганоида.[20]

В осетр имеет ряды ганоидных чешуек, увеличенных в щитообразный броневые листы.

Ганоид чешуйки на Bowfin уменьшены в размерах и напоминают циклоидные весы.




У осетровых рыб чешуя сильно увеличена в броневые пластины по бокам и спине, в то время как у лукового плавника чешуя значительно уменьшена по толщине, чтобы напоминать циклоидные весы.

Серьги из чешуи ганоида аллигатора.

Ископаемое примитивное скатное плавник с ганоидной чешуей



Коренные американцы и люди Карибского моря использовали жесткие ганоидные весы аллигатор гар для наконечников стрел, нагрудников и в качестве защиты от плугов. В настоящее время из этих чешуек делают украшения.[21]
Лептоидные чешуи
Лептоидная (костно-гребенчатая) чешуя встречается у костистых рыб более высокого порядка, костистые кости (чем больше полученный клады лучеконечных рыб). Наружная часть этих чешуек образует веерообразные костные гребни, а внутренняя часть пересечена волокнистой соединительной тканью. Чешуйки лептоидов более тонкие и полупрозрачные, чем чешуйки других типов, и в них отсутствуют затвердевшие эмалевидные или дентинные слои. В отличие от чешуи ганоида, дополнительные чешуи добавляются концентрическими слоями по мере роста рыбы.[22]
Чешуя лептоидов перекрывается в конфигурации голова к хвосту, как черепица, что делает их более гибкими, чем чешуйки космоидов и ганоидов. Такое расположение обеспечивает более плавный поток воды по телу и снижает тащить.[23] Чешуя некоторых видов демонстрирует полосы неравномерного сезонного роста, называемые аннулировать (единственное число кольцо). Эти полосы можно использовать для состарить рыбу.
Чешуя лептоида бывает двух видов: циклоидной и ктеноидной.
Циклоидные весы
Циклоидные (круглые) чешуи имеют гладкую текстуру и однородные, с гладким внешним краем или краем. Чаще всего они встречаются у рыб с мягкими лучами плавников, таких как лосось и карп.
 |  |
Азиатская арована имеют большие циклоидные чешуи, расположенные на рыбе в мозаика приподнятых ребер (слева). Сами чешуйки покрыты ажурным сетчатым узором (справа).[24][25] | |


Циклоидная (круглая) чешуя обычно встречается у карпообразных или лососевых рыб. |



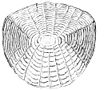



Ктеноидная чешуя
Ктеноидная (зубчатая) чешуя похожа на циклоидную чешую, за исключением того, что у нее маленькие зубцы или шипики называется ctenii по их внешнему или заднему краю. Из-за этих зубов чешуя имеет грубую текстуру. Обычно они встречаются у рыб с лучами колючих плавников, таких как окунь Рыбы. Эти чешуйки почти не содержат кости, поскольку состоят из поверхностного слоя, содержащего гидроксиапатит и карбонат кальция и более глубокий слой, состоящий в основном из коллаген. Эмаль других типов чешуек редуцируется до поверхностных гребней и ктений.


 |  |
Размер зубов на ктеноидных чешуях может варьироваться в зависимости от положения, так как эти чешуйки от крысиный хвост Cetonurus crassiceps Показать. | |
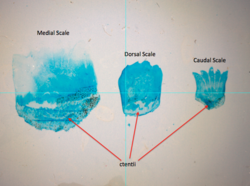

Ктеноидная (зубчатая) чешуя обычно встречается у окуневидных рыб. |





Ктеноидные чешуйки, как и другие эпидермальные структуры, происходят от плакоды и отличительная клеточная дифференциация делает их исключительными из других структур, возникающих из покров.[27] Развитие начинается рядом с хвостовой плавник, вдоль боковая линия рыбы.[28] Процесс разработки начинается с накопления фибробласты между эпидермис и дерма.[27] Коллагеновые фибриллы начинают организовываться в дермальном слое, что приводит к инициированию минерализация.[27] Сначала увеличивается окружность чешуек, а затем толщина, когда перекрывающиеся слои минерализуются вместе.[27]
Чешуйки гребневидной кости можно разделить на три типа:
- Crenate шкалы, где на краю шкалы имеются углубления и выступы.
- Спиноид чешуйки, где чешуя несет шипы, которые продолжаются с самой чешуей.
- Истинный ктеноид чешуйки, где иглы на чешуе представляют собой отдельные структуры.
У большинства рыб с лучевыми плавниками есть гребневидная чешуя. Некоторые виды камбалы имеют ктеноидные чешуйки на глазной стороне и циклоидные чешуйки на слепой стороне, в то время как другие виды имеют ктеноидные чешуйки у мужчин и циклоидные чешуи у женщин.
Отражение



Многие костистые рыбы покрыты чешуей с высокой отражающей способностью, которые действуют как маленькие зеркала и создают вид посеребренного стекла. Отражение через серебрение широко распространено или преобладает у рыб открытого моря, особенно у тех, которые живут в верхних 100 метрах. А прозрачность Эффект может быть достигнут с помощью серебрения, чтобы сделать тело животного сильно отражающим. На средних глубинах в море свет идет сверху, поэтому вертикально ориентированное зеркало делает животных, например рыб, невидимыми сбоку.[29]
В морской топорик чрезвычайно сплюснутый по бокам (из стороны в сторону), оставляя тело толщиной всего миллиметры, а тело настолько серебристое, что напоминает алюминиевая фольга. Зеркала состоят из микроскопических структур, подобных тем, которые используются для структурная окраска: стопки от 5 до 10 кристаллов гуанин расположены на расстоянии около ¼ длины волны друг от друга, чтобы конструктивно интерферировать и обеспечивать почти 100-процентное отражение. В глубоких водах, в которых обитает топорик, только синий свет с длиной волны 500 нанометров проникает вниз и должен отражаться, поэтому зеркала на расстоянии 125 нанометров обеспечивают хорошую маскировку.[29]
Большинство рыб в верхнем слое океана замаскированы серебрением. В рыбе, такой как сельдь, который обитает на более мелкой воде, зеркала должны отражать смесь длин волн, и, соответственно, у рыбы есть стопки кристаллов с диапазоном различных расстояний. Еще одна сложность для рыб с закругленными в поперечном сечении телами заключается в том, что зеркала будут неэффективными, если положить их на кожу, поскольку они не будут отражать горизонтально. Общий зеркальный эффект достигается с помощью множества небольших отражателей, все ориентированных вертикально.[29]
Рыбья чешуя с этими свойствами используется в некоторых косметических средствах, поскольку они могут придавать мерцающий эффект макияжу и помаде.[30]
Плакоидные чешуи

Плакоид (заостренные, зубчатые) чешуйки встречаются в хрящевые рыбы: акулы, лучи. Их еще называют дермальные зубчики. Чешуйки плакоидов структурно гомологичный с позвоночное животное зубы («зубчик» переводится как «маленький зуб»), имеющий центральный полость пульпы снабжен чем то кровеносный сосуд, окруженный коническим слоем дентин, все они расположены на прямоугольной базальной пластине, которая опирается на дерма. Самый внешний слой состоит из витродентин, в основном неорганический эмаль -подобное вещество. Чешуя плакоида не может увеличиваться в размерах, но по мере увеличения размера рыбы добавляется больше чешуи.
Подобные шкалы также можно найти под заголовком зубчатая сельдь. В лучах масштаб покрытия намного меньше.
Акулья кожа

Кожа акулы почти полностью покрыта мелкой плакоидной чешуей. Чешуя поддерживается шипами, которые кажутся шершавыми при движении в обратном направлении, но когда они сглаживаются движением воды вперед, образуются крошечные вихри которые уменьшают гидродинамический тащить и уменьшить турбулентность, что делает плавание более эффективным и тихим по сравнению с плаванием костистых рыб.[31] Он также играет роль в предотвращении обрастания, демонстрируя эффект лотоса.[32]
Все зубчики состоят из внутренней полости пульпы с нервным и артериальным питанием, уходящим корнями в дерма для снабжения зубного камня слизью.[33] Зубцы содержат ребристые структуры, которые выступают за поверхность чешуи; Под микроскопом эта риблочка может выглядеть как крючок или выступающие из чешуи выступы. Общая форма выступа зубца зависит от типа акулы и может быть описана двумя способами.[34] Первая - это шкала, в которой гребни расположены сбоку от акулы и параллельны потоку воды. Вторая форма представляет собой гладкую чешуйку с чем-то вроде крючковатой гребешки, вьющейся из поверхности и направленной к задняя сторона акулы.[34] Обе формы гребня помогают создать турбулентный пограничный слой принуждение ламинарный поток подальше от акульей кожи.[35]
В отличие от костистых рыб, у акул сложная кожная корсет из гибкого коллагеновый волокна устроен как спиральный сеть, окружающая их тело. Корсет работает как внешний скелет, обеспечивая прикрепление плавательных мышц и тем самым экономя энергию.[36] В зависимости от положения этих плакоидных чешуек на теле они могут быть гибкими и пассивно возведенными, что позволяет им изменять угол атаки. Эти чешуйки также имеют выступы, которые выровнены по направлению потока, эти гребни уменьшают силу сопротивления, действующую на кожу акулы, отталкивая вихрь дальше от поверхности кожи, препятствуя любому высокоскоростному поперечному потоку.[37]
Морфология чешуи
Общая анатомия чешуек различна, но все их можно разделить на три части: макушка, шейка и основание. Податливость шкалы связана с размером основания шкалы. Весы с большей гибкостью имеют меньшее основание и поэтому менее жестко прикреплены к stratum laxum. На макушке быстро плавающих акул есть ряд параллельных гребней или гребней, идущих от переднего к заднему направлению.[38]
Анализируя три составляющих шкалы, можно сделать вывод, что основание зубного камня не контактирует ни с какой частью потока жидкости.[39] Однако коронка и шейка зубцов играют ключевую роль и отвечают за создание турбулентных вихрей и водовороты обнаружен у поверхности кожи.[39] Из-за того, что зубчики бывают разных форм и размеров, можно ожидать, что не все формы будут давать одинаковый тип турбулентный поток. Во время недавнего исследовательского эксперимента биомиметик образцы зубчиков акулы с микроструктурой в виде полумесяца, испытанные в резервуаре для воды с использованием вытяжного стола в качестве слайда. Эксперимент показал, что поверхность с зубчиками в целом уменьшила сопротивление на 10% по сравнению с гладким образцом. Причина этого снижения сопротивления была связана с тем, что турбулентные вихри застревали между зубчиками, создавая «подушкообразный» барьер против ламинарного потока.[40] Тот же тип эксперимента был проведен другой исследовательской группой, которая внесла больше изменений в свой биомиметический образец. Вторая группа пришла к такому же выводу, что и первая, однако, поскольку их эксперимент содержал больше вариаций в образцах, они смогли достичь высокой степени экспериментальной точности. В заключение они заявили, что более практичные формы более долговечны, чем формы с замысловатыми гребнями. Практичные формы были низкопрофильными и имели трапециевидное или полукруглое поперечное сечение, похожее на желоб, и были менее эффективными, но, тем не менее, снижали сопротивление на 6 или 7%.[41]
Снижение сопротивления

Акулы уменьшают сопротивление и общее стоимость транспорта (COT) несколькими способами. Сопротивление давления создается из-за разницы давлений между передней и задней стороной акулы из-за объема, который проходит мимо акулы, чтобы продвинуться вперед.[42] Этот тип сопротивления также прямо пропорционален ламинарный поток. Когда ламинарный поток вокруг рыбы увеличивается, сопротивление давления увеличивается.[43] Сопротивление трения является результатом взаимодействия жидкости с кожей акулы и может варьироваться в зависимости от того, как изменяется пограничный слой относительно поверхности рыбы.[42]
Рифлеты препятствуют продольному перемещению продольных вихрей в вязком подслое. Механизм сложен и еще не полностью изучен. По сути, гребни препятствуют образованию вихрей у поверхности, потому что вихрь не может поместиться в впадинах, образованных гребнями. Это выталкивает вихрь дальше от поверхности, взаимодействуя только с кончиками гребней, не вызывая высокоскоростного потока в долинах. Поскольку этот высокоскоростной поток теперь взаимодействует только с кончиком риблета, который представляет собой очень маленькую площадь поверхности, передача импульса, вызывающая сопротивление, теперь намного ниже, чем раньше, что эффективно снижает сопротивление. Кроме того, это уменьшает колебания скорости поперечного потока, что также способствует передаче импульса.[38]
Недавние исследования показали, что в пристеночном пограничном слое существует режим до и после пробоя, где подслой сгущается с убывающей скоростью, а затем резко распадается на турбулентные вихри, прежде чем окончательно схлопнуться. Эта система полностью саморегулируется и опосредует цикл роста и распада; вихри накапливаются в период роста и резко ликвидируются в Массивы Струхаля шпилечных вихрей, отрывающихся от стены. Подъемные вихри - это то, что выталкивает пограничный слой с поверхности акулы, что приводит к снижению общего сопротивления, испытываемого рыбой.[44]
Грубо, наждачная бумага -подобная текстура кожи акулы и ската, вкупе с прочностью, позволила ценить ее как источник сыромятной кожи натуральная кожа, называется шагрень. Одним из многих исторических применений шагрени из акулы было изготовление рукояток для мечи. Шероховатая текстура кожи также используется в Японская кухня сделать терки называется орошики, прикрепив кусочки акульей кожи к деревянным доскам. Небольшой размер чешуек очень мелко натирает пищу.
Техническое приложение

В морской индустрии существует чрезвычайно большой рынок и потребность в противообрастающие поверхности. С точки зрения непрофессионала, обрастание известно как процесс, при котором что-то покрывается материалом из окружающей среды, например ракушки, водоросли, и зеленый ил. Кожные зубчики являются чрезвычайно многообещающей областью исследований для этого типа применения в связи с тем, что акулы - одна из единственных рыб, у которых чешуя не нарастает и не накапливается. Исследования ВМС США показали, что если биомиметический материал может быть разработан, он потенциально может привести к увеличению экономии топлива на военных судах до 45%.[45]
Есть много примеров биомиметические материалы и поверхности, основанные на строении водных организмов, включая акул. Такие приложения предназначены для обеспечения более эффективного движения в текучих средах, таких как воздух, вода и масло.
Поверхности, имитирующие кожу акулы, также использовались для удержания микроорганизмов и водоросли от покрытия корпусов подводных лодок и кораблей. Один сорт продается как "акула ".[46][47]
Многие новые методы воспроизведения кожи акулы включают использование полидиметилсилоксан (PDMS) для создания пресс-формы. Обычно процесс включает взятие плоского куска акульей кожи, покрытие его PDMS, чтобы сформировать форму, и повторная заливка PDMS в эту форму, чтобы получить точную копию кожи акулы. Этот метод был использован для создания биомиметической поверхности, которая имеет супергидрофобный свойства, демонстрирующие эффект лотоса.[46] Одно исследование показало, что эти биомиметические поверхности снижают сопротивление до 9%,[37] при этом при маховом движении снижение сопротивления достигло 12,3%.[48]
Наряду с морскими приложениями, аэрокосмическая промышленность также может извлечь выгоду из этих биомиметических конструкций. Параметрическое моделирование было сделано на зубцах акул с широким диапазоном конструктивных вариаций, таких как низкопрофильные и высокопрофильные генераторы вихрей.[49] Эти биомиметические модели были разработаны и проанализированы, чтобы увидеть эффекты применения зубчатых структур на крыльях различных самолетов. Во время моделирования было отмечено, что образец изменил то, как низкие и высокие углы атаки отреагировал. Из испытанных образцов с низким и высоким профилем низкопрофильные вихревые генераторы превзошли существующие конструкции с гладким крылом на 323%. Это повышение производительности происходит из-за разделительного пузыря в следе от зубцов и потоковых вихрей, которые восполняют потерю импульса в пограничном слое из-за поверхностного трения.[49]
Щитки

Щитки похожи на весы и выполняют ту же функцию. В отличие от чешуи рыб, которые образуются из эпидермиса, щитки образуются в нижнем сосудистом слое кожи, а эпидермальный элемент - только на верхней поверхности. Формируясь в живой дерме, щитки образуют роговой внешний слой, внешне похожий на чешуйки.
Scute происходит от латинского для щит, и может принимать форму:
- внешняя щитообразная костная пластина, или
- видоизмененная утолщенная чешуя, часто килеватая или колючая, или
- выступающая, видоизмененная (грубая и сильно гребенчатая) чешуя, обычно связанная с боковой линией или на хвостовом стебле, образующем хвостовые кили, или вдоль вентрального профиля.
Некоторые рыбы, например сосновая шишка, полностью или частично покрыты щитками. Речная сельдь и плавники имеют брюшной ряд щитков, которые представляют собой чешуйки с приподнятыми острыми концами, которые используются для защиты. Немного домкраты иметь ряд щитков, следующих за боковая линия с обеих сторон.
Масштаб развития
Чешуя обычно появляется на поздних стадиях развития рыб. В случае данио, требуется 30 дней после оплодотворения, прежде чем разные слои, необходимые для начала формирования чешуек, дифференцируются и организуются. Для этого необходимо, чтобы консолидация мезенхима происходит тогда морфогенез индуцируется, и, наконец, процесс дифференциации или поздний метаморфоза происходит.[50][51]
- Консолидация мезенхимы: консолидация или структурирование мезенхимы происходит во время развития дерма. Этот процесс зависит от того, является ли рыба хрящевой или костной. У хрящевых рыб структурирование происходит за счет образования двух слоев. Первый поверхностный и широкий, а второй тонкий и компактный. Эти два слоя разделены мезенхимальные клетки. Костные рыбы производят бесклеточный субстрат организован перпендикулярно коллагеновые волокна. Впоследствии для обеих рыб фибробласты удлиненный.Они проникают в плотный слой мезенхимы, который консолидируется до образования чешуи, чтобы инициировать дермальную пластинку.[50][51][52]
- Индукция морфогенеза: морфогенез обусловлен формированием эпидермального сосочек, который генерируется присоединением эпидермис и дерма через процесс инвагинация. Морфогенез начинается в тот момент, когда фибробласты перемещаются в верхнюю часть компактной мезенхимы. На протяжении всего этого процесса базальные клетки из эпителий образуют разграничивающий слой, который располагается в верхней части мезенхимы. Впоследствии эти клетки будут дифференцироваться в области, где чешуя зачаток возникнет.[50][51][52]
- Дифференциация или поздняя метаморфоза: эта дифференциация порождается двумя разными формами в соответствии с типом формируемой шкалы. Формирование эластоидных чешуек (циклоидов и гребневиков) происходит за счет образования пространства между матрицей эпидермального сосочка. Это пространство содержит волокна коллагена. Вокруг этого пространства дифференцируются эластобласты, которые отвечают за выработку материала, необходимого для образования чешуи. Впоследствии матричная минерализация происходит, позволяя шкале приобрести жесткую характеристику, которая их идентифицирует.[50][51][52]
В отличие от эластоидных чешуек, ганоидная чешуя состоят из минерализованного и неминерализованного коллагена в разных регионах. Их образование происходит за счет проникновения поверхностных клеток мезенхимы в матрикс, последний состоит из коллагеновых волокон и располагается вокруг сосудистых капилляров, что приводит к возникновению сосудистых полостей. На этом этапе эластобласты заменяются на остеобласты, образуя кость. Патчи матрицы шкалы не окостеневший состоят из уплотненного коллагена, что позволяет ему поддерживать соединение с мезенхимой. Это известно как Волокна Шарпея.[50][51][52]
Одним из генов, регулирующих развитие образования чешуи у рыб, является звуковой еж (shh) ген, который посредством белка (shh) участвует в органогенез и в процессе сотовая связь, разрешите формирование чешуек.[53][54] В аполипопротеин E (ApoE), что позволяет транспортировать и метаболизировать триглицериды и холестерин, взаимодействует с shh, потому что ApoE обеспечивает холестерином shh сигнальный путь. Было показано, что в процессе дифференциация клеток и взаимодействие уровень транскрипции ApoE высокий, что позволило сделать вывод о важности этого белка для позднего развития чешуек.[53][54]
Модифицированные весы




Различные группы рыб имеют развился ряд модифицированных весов для выполнения различных функций.
- Почти у всех рыб есть боковая линия, система механорецепторы которые обнаруживают движение воды. У костистых рыб чешуя вдоль боковой линии имеет центральные поры, которые позволяют воде контактировать с сенсорными клетками.
- Шипы спинного плавника морские акулы и химеры, жалящие хвостовые шипы скаты, а «пильные» зубы пилильщики и акулы сросшиеся и видоизмененные чешуйки плакоида.
- Рыба-хирург иметь лезвие, похожее на скальпель, которое представляет собой модифицированную шкалу, по обе стороны от хвостовой стебель.[55]
- Немного селедка, анчоусы, и полуклювы имеют лиственные чешуи, которые легко сбрасываются и помогают убежать от хищников.
- Мужской Percina у стрелков есть ряд увеличенных кадукозная чешуя между тазовые плавники и анус.
- Рыбы-дикобразы имеют шкалы, преобразованные в большие внешние шипы.
- Напротив, рыба фугу у них более тонкие и скрытые шипы, чем у рыб-дикобразов, которые становятся видимыми только тогда, когда рыба надувается. В отличие от рыбы-дикобраза, эти шипы не являются модифицированной чешуей, а развиваются под контролем той же сети генов, которые производят перья и волосы у других позвоночных.[56][57]

Рыба-дикобраз изменить шкалы на шипы.
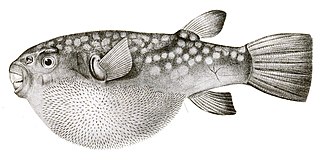
Рыба фугу шипы не являются модифицированными чешуйками, а развиваются независимой генной сетью.

Рыба без чешуи

Мандаринка не имеют чешуи и защищаются слоем вонючей и горькой слизи.

У рыб без чешуи обычно появляются альтернативы защитной чешуе, такие как жесткая кожистая кожа или костные пластины.
- Рыба без челюсти (миноги и микробы ) имеют гладкую кожу без чешуек и без кожной кости.[58] Миноги получают некоторую защиту благодаря жесткой кожистой шкуре. Хагфиш выделяет обильное количество слизи или слизь если им угрожают.[59] Они могут связать себя узел сверху, соскребая слизь на ходу и освобождаясь от хищника.[60]
- Наиболее угри безчешуйчатые, хотя некоторые виды покрыты крошечной гладкой циклоидной чешуей
- Наиболее сом отсутствуют чешуйки, хотя у некоторых семейств есть нательные доспехи в виде кожных пластинок или какого-либо щитка.[61]
- Мандаринка не имеют чешуек и имеют слой вонючей и горькой слизи, которая блокирует болезнь и, вероятно, отпугивает хищников, подразумевая, что их яркая окраска апосематический.[62]
- Удильщик иметь дряблую, тонкую кожу, часто покрытую тонкими раздвоенными кожными колючками или бугорки, но у них нет штатных весов. Они полагаются на камуфляж, чтобы избежать внимания хищников, а их дряблая кожа мешает хищникам схватить их.
Многие группы костистых рыб, в том числе игла, морские коньки, самшит, браконьеры, и несколько семейств колюшки, развили внешние костные пластинки, структурно напоминающие чешуйки плакоида, как защитную броню от хищников.
- У морских коньков нет чешуи, но они имеют тонкую кожу, натянутую на костяные пластинчатые доспехи, расположенные кольцами по всей длине тела.
- У самшита пластины сливаются вместе, образуя твердый панцирь или экзоскелет охватывая все тело. Эти костные пластинки представляют собой не модифицированные чешуйки, а окостеневшую кожу. Из-за этой тяжелой брони боксфиш могут двигаться медленно, но немногие другие рыбы способны есть взрослых особей.
 | |
Угри кажутся бесчешуйчатыми, но некоторые виды покрыты крошечной гладкой циклоидной чешуей. | |
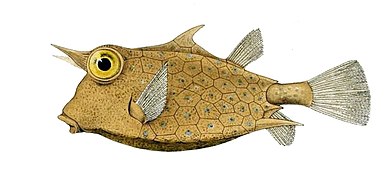
Boxfish имеют пластины окостеневшей кожи, слитые вместе, образуя твердую оболочку.

Морские коньки имеют тонкую кожу, натянутую на костные пластинки, расположенные кольцами.

Некоторые рыбы, такие как хоки и рыба-меч, рождаются с чешуей, но сбрасывают ее по мере роста.
Файфиш имеет грубую неперекрывающуюся чешую с небольшими шипами, поэтому их называют рыб-филлфиш. Некоторые рыбки кажутся безчешуйными, потому что их чешуя очень мала.
Заметное масштабирование появляется на тунец только по боковой линии и в корсет, защитная полоса из утолщенных и увеличенных чешуек в области плеча. На большей части тела у тунца чешуя настолько мала, что на первый взгляд кажется без чешуи.[63]

Немного рыба кажутся безмасштабными, потому что их чешуя такая маленькая

К случайному осмотру тунец кажутся в значительной степени свободными от чешуи, но это не так.


Левит
Знаменитый отрывок в Левит заявляет, что «из всех, что в водах ... в морях и реках», те, у которых нет и плавников, и чешуи, «будут мерзостью для вас» и не могут быть съедены.[64] Это устраняет все водные беспозвоночные в качестве мерзости и нечистый, а также любую рыбу без чешуи (вроде бы не бывает рыб без плавников).
Согласно подавиться или божественные указы Тора и Талмуд, для объявления рыбы кошерный, у него должны быть чешуя и плавники.[65] Определение «чешуя» отличается от определений, представленных в биологии, тем, что чешуя кошерной рыбы должна быть видна глазу, присутствовать во взрослой форме и может быть легко удалена с кожи вручную или ножом для чистки.[65] Согласно агентство кошерной сертификации из Православный союз, рыба считается кошерной, если чешую можно удалить, не повредив кожу.[66] Таким образом, карп и лосось кошерные, в то время как акула с микроскопической чешуей, осетр, чьи щитки нельзя легко удалить, не вырезая их из тела, и рыба-меч, теряющая чешую во взрослом возрасте, не кошерные. К другим некошерным рыбам относятся сом, угри, тихоокеанская треска, змеиная скумбрия и рыба фугу.[65]
Лепидофагия

Лепидофагия (Древнегреческий для щитовка) - это особый способ кормления рыб, связанный с поеданием чешуи других рыб.[68] Лепидофагия имеет самостоятельно развился по крайней мере в пяти пресноводных семьях и семи морских семьях.[69]
Рыбья чешуя может быть питательной, содержать кожную часть и слой богатой белком слизи помимо слоев кератин и эмаль. Они являются богатым источником фосфат кальция.[69] Однако энергия, затрачиваемая на нанесение удара по сравнению с количеством чешуи, потребляемой на удар, накладывает ограничение на размер чешуекрылых рыб, и обычно они намного меньше их добычи.[69] Пищевое поведение из-за чешуек обычно развивается из-за недостатка пищи и экстремальных условий окружающей среды. Поедание чешуи и кожи, окружающей чешуйки, обеспечивает богатые белком питательные вещества, которые могут быть недоступны в других местах этой ниши.[70]
Рыбные челюсти обычно показывают двусторонняя симметрия. Исключение составляет щитовка. цихлида Perissodus microlepis. Челюсти этой рыбы представлены двумя разными морфологический формы. У одного морфа челюсть вывернута влево, что позволяет ему с большей готовностью поедать чешую на правом боку жертвы. У другого морфа челюсть повернута вправо, что облегчает поедание чешуи с левого бока жертвы. Относительная численность двух морф в популяциях регулируется частотно-зависимый выбор.[67][71][72]
Смотрите также
- Определение возраста у рыб
- Окраска животных
- Отражатели для животных
- Фотонные кристаллы
- Чешуя рептилий
- Масштаб (зоология)
- Чешуйчатая броня
- Змеиная чешуя
- Урокотори - Японская рыбочистка
Рекомендации
- ^ Шкала Etymonline. Проверено 28 апреля 2019.
- ^ Mongera, A .; Нюссляйн-Фольхард, К. (2013). «Чешуя рыбы возникает из мезодермы». Текущая биология. 23 (9): R338 – R339. Дои:10.1016 / j.cub.2013.02.056. PMID 23660349.
- ^ Шарп, П. Т. (2001). «Развитие рыбьей чешуи: волосы сегодня, зубы и чешуя вчера?». Текущая биология. 11 (18): R751 – R752. Дои:10.1016 / S0960-9822 (01) 00438-9. PMID 11566120. S2CID 18868124.
- ^ Перкинс, Сид (16 октября 2013 г.). "Первые ложные зубы". Наука. Получено 2 марта 2018.
- ^ Тернер, С .; Тарлинг, Д. Х. (1982). «Распространение телодонта и других бесчеловечных галактик как тесты континентальных реконструкций нижнего палеозоя». Палеогеография, палеоклиматология, палеоэкология. 39 (3–4): 295–311. Bibcode:1982ППП .... 39..295Т. Дои:10.1016 / 0031-0182 (82) 90027-X.
- ^ Мерсс, Т. (2006). «Ультраскелетная скульптура ранних позвоночных». Журнал палеонтологии позвоночных. 26 (2): 235–252. Дои:10.1671 / 0272-4634 (2006) 26 [235: EUOEV] 2.0.CO; 2.
- ^ а б Жанвье, Филипп (1998). «Ранние позвоночные и их современные родственники». Ранние позвоночные. Oxford University Press. С. 123–127. ISBN 978-0-19-854047-2.
- ^ а б Тернер, С. (1991). «Монофилия и взаимоотношения телодонтов». В М. М. Чанг; Ю. Х. Лю; Г. Р. Чжан (ред.). Ранние позвоночные и связанные с ними проблемы эволюционной биологии. Science Press, Пекин. С. 87–119.
- ^ Мерсс, Т. (1986). "Сквамация телодонта агнатана" Флеболепис". Журнал палеонтологии позвоночных. 6 (1): 1–11. Дои:10.1080/02724634.1986.10011593.
- ^ Botella, H .; Х. И. Валенсуэла-Риос; П. Карлс (2006). «Новый раннедевонский телодонт из Кельтиберии (Испания) с переработкой испанских телодонтов». Палеонтология. 49 (1): 141–154. Дои:10.1111 / j.1475-4983.2005.00534.x.
- ^ Ferrón, Humberto G .; Ботелла, Эктор (2017). «Сквамация и экология телодонтов». PLOS ONE. 12 (2): e0172781. Bibcode:2017PLoSO..1272781F. Дои:10.1371 / journal.pone.0172781. ЧВК 5328365. PMID 28241029.
- ^ МАЙКЛ ОЛЛАБИ «Космоидная шкала». Словарь зоологии. . Encyclopedia.com. 29 октября 2019 <https://www.encyclopedia.com >
- ^ Зильберберг, Л., Менье, Ф.Дж., Лорин, М. (2010). Микроанатомическое и гистологическое исследование посткраниального дермального скелета девонского саркоптерига. Eusthenopteron foordi, Acta Palaeontologica Polonica 55: 459–470.
- ^ Берген, Дилан Дж. М .; Каге, Эрика; Хаммонд, Крисси Л. (2019). «Рыбы данио как новая модель остеопороза: платформа для первичного тестирования новых остеоактивных соединений». Границы эндокринологии. 10: 6. Дои:10.3389 / fendo.2019.00006. ISSN 1664-2392. ЧВК 6361756. PMID 30761080.
- ^ de Vrieze, E .; van Kessel, M.A.H.J .; Peters, H.M .; Spanings, F.A.T .; Flik, G .; Мец, Дж. Р. (01.02.2014). «Преднизолон вызывает фенотип, подобный остеопорозу, в регенерирующих чешуях рыбок данио». Остеопороз Интернэшнл. 25 (2): 567–578. Дои:10.1007 / s00198-013-2441-3. ISSN 1433-2965. PMID 23903952. S2CID 21829206.
- ^ а б Zylberberg, L .; Отец, J. -Y .; Нанси, А. (1997). «Иммунодетекция амелогенин-подобных белков в ганоине экспериментально регенерирующих чешуек Calamoichthys calabaricus, примитивных актиноптеригийских рыб». Анатомический рекорд. 249 (1): 86–95. Дои:10.1002 / (SICI) 1097-0185 (199709) 249: 1 <86 :: AID-AR11> 3.0.CO; 2-X. PMID 9294653.
- ^ Отец, Жан-Ив; Донохью, Филип С. Дж .; Викариус, Мэтьюз К. (2009). «Происхождение и эволюция покровного скелета у позвоночных, не являющихся четвероногими». Журнал анатомии. 214 (4): 409–440. Дои:10.1111 / j.1469-7580.2009.01046.x. ISSN 0021-8782. ЧВК 2736117. PMID 19422423.
- ^ а б c Рихтер, М. (1995). «Микроструктурное исследование ткани ганоина отобранных низших позвоночных». Зоологический журнал Линнеевского общества. 114 (2): 173–212. Дои:10.1006 / zjls.1995.0023.
- ^ а б Bruet, B.J. F .; Song, J .; Boyce, M.C .; Ортис, К. (2008). «Принципы проектирования материалов древних рыбных доспехов». Материалы Природы. 7 (9): 748–756. Bibcode:2008НатМа ... 7..748Б. Дои:10.1038 / nmat2231. PMID 18660814.
- ^ а б Шерман, Винсент Р .; Яраги, Николай А .; Кисаил, Давид; Мейерс, Марк А. (2016-12-01). «Микроструктурные и геометрические влияния в защитных чешуях Atractosteus spatula». Журнал интерфейса Королевского общества. 13 (125): 20160595. Дои:10.1098 / rsif.2016.0595. ISSN 1742-5689. ЧВК 5221522. PMID 27974575.
- ^ «План управления и восстановления Миссурийского аллигатора» (PDF). Департамент природоохранного рыболовства штата Миссури. 22 января 2013 г. Архивировано с оригинал (PDF) 6 мая 2016 г.. Получено 12 апреля, 2019.
- ^ Лаглер, К. Ф., Дж. Э. Бардах и Р. Р. Миллер (1962) Ихтиология. Нью-Йорк: Джон Вили и сыновья.
- ^ Баллард, Бонни; Чик, Райан (2 июля 2016 г.). Лекарство от экзотических животных для ветеринарного врача. Джон Вили и сыновья. ISBN 978-1-118-92421-1.
- ^ Pouyaud, L .; Сударто, Гай Г. Тойгелс (2003). «Разные по окраске разновидности азиатской arowana Scleropages formosus (Osteoglossidae) являются разными видами: морфологические и генетические свидетельства». Cybium. 27 (4): 287–305.
- ^ Исмаил, М. (1989). Систематика, зоогеография и сохранение пресноводных рыб полуострова Малайзия (Докторская диссертация под ред.). Государственный университет Колорадо.
- ^ E.J. Брилл (1953). Рыбы Индо-Австралийского архипелага. E.J. Брилл. С. 306–307.
- ^ а б c d Кавасаки, Кента С., «Генетический анализ морфологии чешуи цихлид» (2016). Магистерские диссертации май 2014 г. - действ. 425. http://scholarworks.umass.edu/masters_theses_2/425
- ^ Хельфман, Джин (2009). Разнообразие биологии, эволюции и экологии рыб. Вили-Блэквелл.
- ^ а б c Селедка, Питер (2002). Биология глубокого океана. Оксфорд: Издательство Оксфордского университета. С. 193–195. ISBN 9780198549567.
- ^ "Вероятно, в вашей помаде есть рыбья чешуя". HuffPost Индия. 2015-04-23. Получено 2019-05-06.
- ^ Мартин, Р. Эйдан. "Кожа зубов". Получено 2007-08-28.
- ^ Фюрстнер, Райнер; Бартлотт, Вильгельм; Нейнхейс, Кристоф; Вальцель, Питер (2005-02-01). «Смачивающие и самоочищающиеся свойства искусственных супергидрофобных поверхностей». Langmuir. 21 (3): 956–961. Дои:10.1021 / la0401011. ISSN 0743-7463. PMID 15667174.
- ^ Лаудер, Джордж V .; Уэйнрайт, Дилан К .; Domel, August G .; Уивер, Джеймс С.; Вэнь, Ли; Бертольди, Катя (2016). «Структура, биомиметика и гидродинамика поверхности кожи рыб». Physical Review Fluids. 1 (6): 060502. Bibcode:2016ПхРвФ ... 1ф0502Л. Дои:10.1103 / PhysRevFluids.1.060502. S2CID 18118663.
- ^ а б Фельд, Катрин; Колборг, Энн Ноер; Нюборг, Камилла Мари; Салевский, Мирко; Стеффенсен, Джон Фленг; Берг-Соренсен, Кирстина (24 мая 2019 г.). «Зубцы кожи трех медленно плавающих видов акул: микроскопия и визуализация потока». Биомиметики. 4 (2): 38. Дои:10.3390 / biomimetics4020038. ISSN 2313-7673. ЧВК 6631580. PMID 31137624.
- ^ Флетчер, Томас; Альтрингем, Джон; Пиколл, Джеффри; Wignall, Поль; Доррелл, Роберт (7 августа 2014 г.). «Гидродинамика ископаемых рыб». Труды Королевского общества B: биологические науки. 281 (1788): 20140703. Дои:10.1098 / rspb.2014.0703. ISSN 0962-8452. ЧВК 4083790. PMID 24943377.
- ^ Мартин, Р. Эйдан. «Важность хрящевой ткани». Центр исследования акул ReefQuest. Получено 2009-08-29.
- ^ а б Hage, W .; Bruse, M .; Бехерт, Д. У. (1 мая 2000 г.). «Эксперименты с трехмерными риблами как идеализированная модель акульей кожи». Эксперименты с жидкостями. 28 (5): 403–412. Bibcode:2000ExFl ... 28..403B. Дои:10.1007 / s003480050400. ISSN 1432-1114. S2CID 122574419.
- ^ а б Мотта, Филипп; Хабеггер, Мария Лаура; Лэнг, Эми; Хютер, Роберт; Дэвис, Джессика (01.10.2012). «Морфология чешуи и гибкость короткоперого мако Isurus oxyrinchus и черноперой акулы Carcharhinus limbatus». Журнал морфологии. 273 (10): 1096–1110. Дои:10.1002 / jmor.20047. ISSN 1097-4687. PMID 22730019. S2CID 23881820.
- ^ а б Доу, Чжаолян; Ван, Цзядао; Чен, Даронг (1 декабря 2012 г.). «Бионические исследования рыбьей чешуи для уменьшения сопротивления». Журнал бионической инженерии. 9 (4): 457–464. Дои:10.1016 / S1672-6529 (11) 60140-6. ISSN 1672-6529. S2CID 137143652.
- ^ «Экспериментальные исследования характеристик снижения сопротивления бионической поверхности с захватывающими воду микроструктурами рыбьей чешуи» (PDF).
- ^ Палмер, Колин; Янг, Марк Т. (14 января 2015 г.). «Уменьшение сопротивления поверхности и контроль разделения потока у пелагических позвоночных, что имеет значение для интерпретации морфологии чешуек в таксонах ископаемых». Королевское общество открытой науки. 2 (1): 140163. Bibcode:2015RSOS .... 240163P. Дои:10.1098 / rsos.140163. ISSN 2054-5703. ЧВК 4448786. PMID 26064576.
- ^ а б Лаудер, Джордж V .; Уэйнрайт, Дилан К .; Domel, August G .; Уивер, Джеймс С.; Вэнь, Ли; Бертольди, Катя (18 октября 2016 г.). «Структура, биомиметика и гидродинамика поверхности кожи рыб». Physical Review Fluids. 1 (6): 060502. Bibcode:2016ПхРвФ ... 1ф0502Л. Дои:10.1103 / PhysRevFluids.1.060502.
- ^ Мутурамалингам, Мутукумар; Villemin, Leo S .; Брюкер, Кристоф (29 апреля 2019 г.). «Образование полос в потоке над биомиметическими массивами рыбьей чешуи». Журнал экспериментальной биологии. 222 (Пт 16): jeb205963. arXiv:1904.12752. Bibcode:2019arXiv190412752M. Дои:10.1242 / jeb.205963. PMID 31375542. S2CID 139103148.
- ^ Bandyopadhyay, Promode R .; Хеллум, Арен М. (23 октября 2014 г.). «Моделирование того, как образцы кожи акул и дельфинов управляют образцами завихренности переходной стенки и турбулентности с помощью пространственно-временных механизмов сброса фазы». Научные отчеты. 4: 6650. Bibcode:2014НатСР ... 4Э6650Б. Дои:10.1038 / srep06650. ISSN 2045-2322. ЧВК 4206846. PMID 25338940.
- ^ Magin, Chelsea M .; Купер, Скотт П .; Бреннан, Энтони Б. (1 апреля 2010 г.). «Нетоксичные противообрастающие стратегии». Материалы сегодня. 13 (4): 36–44. Дои:10.1016 / S1369-7021 (10) 70058-4. ISSN 1369-7021.
- ^ а б Лю, Юньхун; Ли, Гуанджи (2012-12-15). «Новый метод создания« эффекта лотоса »на биомиметической коже акулы». Журнал коллоидной и интерфейсной науки. 388 (1): 235–242. Bibcode:2012JCIS..388..235L. Дои:10.1016 / j.jcis.2012.08.033. ISSN 0021-9797. PMID 22995249.
- ^ "Sharklet Discovery | Sharklet Technologies, Inc". www.sharklet.com. Получено 2018-09-26.
- ^ Лаудер, Джордж V .; Оффнер, Йоханнес (01.03.2012). «Гидродинамическая функция кожи акулы и два биомиметических применения». Журнал экспериментальной биологии. 215 (5): 785–795. Дои:10.1242 / jeb.063040. ISSN 1477-9145. PMID 22323201.
- ^ а б Domel, August G .; Саадат, Мехди; Уивер, Джеймс С.; Хадж-Харири, Хоссейн; Бертольди, Катя; Лаудер, Джордж В. (28 февраля 2018 г.). «Дизайн в стиле кожи акулы, улучшающий аэродинамические характеристики». Журнал интерфейса Королевского общества. 15 (139): 20170828. Дои:10.1098 / rsif.2017.0828. ЧВК 5832729. PMID 29436512.
- ^ а б c d е Сир, J.Y .; Huysseune, A.N.N. (2003). «Формирование кожных скелетных и зубных тканей у рыб: сравнительный и эволюционный подход». Биологические обзоры. 78 (2): 219–249. Дои:10.1017 / S1464793102006073. PMID 12803422. S2CID 19556201.
- ^ а б c d е Le Guellec, D .; Morvan-Dubois, G .; Отец, J.Y. (2004). "Развитие кожи костистых рыб с особым упором на отложение коллагена в дерме рыбок данио (Данио Рерио)". Международный журнал биологии развития. 48 (2–3): 217–231. Дои:10.1387 / ijdb.15272388. PMID 15272388.
- ^ а б c d Отец, J.Y. (2001). «Зубы вне рта у костистых рыб: как извлечь выгоду из нарушения развития». Эволюция и развитие. 3 (2): 104–108. Дои:10.1046 / j.1525-142x.2001.003002104.x. PMID 11341672. S2CID 13353402.
- ^ а б Сир, J.Y .; Акименко, М.А. (2003). «Развитие чешуи у рыб: обзор с описанием экспрессии звукового ежа (шш) у рыбок данио (Данио Рерио)". Международный журнал биологии развития. 48 (2–3): 233–247. Дои:10.1387 / ijdb.15272389. PMID 15272389.
- ^ а б Monnot, M.J .; Бабин, П.Дж .; Poleo, G .; Андре, М .; Laforest, L .; Ballagny, C .; Акименко, М.А. (1999). «Эпидермальная экспрессия гена аполипопротеина E во время развития плавников и чешуек, а также регенерации плавников у рыбок данио». Динамика развития. 214 (3): 207–215. Дои:10.1002 / (SICI) 1097-0177 (199903) 214: 3 <207 :: AID-AJA4> 3.0.CO; 2-5. PMID 10090147.
- ^ Соренсон, Л .; Сантини, Ф .; Carnevale, G .; Альфаро, M.E. (2013). «Временное дерево с несколькими локусами рыб-хирургов (Acanthuridae, Percomorpha) с пересмотренной семейной таксономией». Молекулярная филогенетика и эволюция. 68 (1): 150–160. Дои:10.1016 / j.ympev.2013.03.014. PMID 23542000.
- ^ Как у иглобрюха появились странные колючки Phys.org, 25 июля 2019.
- ^ Shono, T .; Thiery, A.P .; Cooper, R.L .; Курокава, Д .; Britz, R .; Окабе, М .; Фрейзер, Г.Дж. (2019). «Эволюция и разнообразие шипов на коже иглобрюхих рыб». iScience. 19: 1248–1259. Дои:10.1016 / j.isci.2019.06.003. ЧВК 6831732. PMID 31353167.
- ^ Кулидж Э., Хедрик М.С. и Милсом В.К. (2011) «Вентиляционные системы». В: McKenzie DJ, Farrell AP и Brauner CJ (ред.) Физиология рыб: примитивные рыбы, Elsevier, стр. 182–213. ISBN 9780080549521
- ^ Ротшильд, Анна (01.04.2013). "Слизь Hagfish: одежда будущего?". Новости BBC. Получено 2013-04-02.
- ^ Йонг, Эд (2019-01-23). «Никто не подготовлен к слизи миксины». Атлантический океан. Получено 2019-01-26.
- ^ Фрил, Дж. П.; Лундберг, Дж. Г. (1996). "Микромизон акамай, ген. et sp. nov., небольшой безглазый сом банджо (Siluriformes: Aspredinidae) из речных русел нижнего бассейна Амазонки ". Копея. 1996 (3): 641–648. Дои:10.2307/1447528. JSTOR 1447528.
- ^ Садовый, Ю .; Randall, J.E .; Расотто, Мария Б. (май 2005 г.). «Структура кожи у шести видов драконов (Gobiesociformes; Callionymidae): межвидовые различия в типах железистых клеток и секреции слизи». Журнал биологии рыб. 66 (5): 1411–1418. Дои:10.1111 / j.0022-1112.2005.00692.x.
- ^ Есть ли у тунцов чешуя? Северо-восточный научный центр рыболовства, NOAA Fisheries. Доступ 4 августа 2019 г.
- ^ Левит 11: 9–10
- ^ а б c Арье Цитрон, "Все о кошерной рыбе"
- ^ Проверка кошерной рыбы Сертификация кошерности OU. Дата обращения 9 августа 2019.
- ^ а б Lee, H.J .; Kusche, H .; Мейер, А. (2012). «Ручное кормодобывание у цихлид, питающихся чешуей: его потенциальная роль в формировании морфологической асимметрии». PLOS ONE. 7 (9): e44670. Bibcode:2012PLoSO ... 744670L. Дои:10.1371 / journal.pone.0044670. ЧВК 3435272. PMID 22970282.
- ^ Froese, R. и D. Pauly. Редакторы. «Глоссарий: лепидофагия». FishBase. Получено 2007-04-12.
- ^ а б c Яновец, Джефф (2005). «Функциональная морфология кормления у чешуеда» Катоприон менто" (PDF). Журнал экспериментальной биологии. 208 (Pt 24): 4757–4768. Дои:10.1242 / jeb.01938. PMID 16326957. S2CID 15566769.
- ^ Martin, C .; ПК. Уэйнрайт (2011). «Трофическая новизна связана с исключительными темпами морфологического разнообразия двух видов адаптивной радиации куколки Cyprinodon». Эволюция. 65 (8): 2197–2212. Дои:10.1111 / j.1558-5646.2011.01294.x. PMID 21790569. S2CID 23695342.
- ^ Хори, М. (1993). «Частотно-зависимый естественный отбор в маневренности щитовок цихлид». Наука. 260 (5105): 216–219. Bibcode:1993Наука ... 260..216H. Дои:10.1126 / science.260.5105.216. PMID 17807183. S2CID 33113282.
- ^ Стюарт, Т. А .; Альбертсон, Р. К. (2010). «Эволюция уникального аппарата для кормления хищников: функциональная анатомия, развитие и генетический локус латеральности челюстей у цихлид, питающихся чешуей озера Танганьика». BMC Биология. 8 (1): 8. Дои:10.1186/1741-7007-8-8. ЧВК 2828976. PMID 20102595.
дальнейшее чтение
- Хельфман, Г.С., Б.Б. Коллетт, Д.Е. Facey (1997). Разнообразие рыб. Blackwell Science. С. 33–36. ISBN 978-0-86542-256-8.CS1 maint: несколько имен: список авторов (связь)
- Шульце, Х. (2016). «Чешуя, эмаль, космин, ганоин и ранние остеихтианы». Comptes Rendus Palevol. 15 (1–2): 83–102. Дои:10.1016 / j.crpv.2015.04.001.
| О рыбы |  | ||||
|---|---|---|---|---|---|
| Анатомия и физиология | |||||
| Сенсорный системы | |||||
| Размножение | |||||
| Передвижение | |||||
| Другой поведение | |||||
| К среда обитания | |||||
| Другие типы | |||||
| Коммерческий |
| ||||
| Основные группы | |||||
| Списки | |||||
| |||||