Lystrosaurus - Lystrosaurus
| Lystrosaurus | |
|---|---|
 | |
| Lystrosaurus hedini скелет в Музее палеонтологии, Тюбинген | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Терапсида |
| Clade: | †Дицинодонтия |
| Семья: | †Lystrosauridae |
| Род: | †Lystrosaurus Справиться, 1870 |
| Разновидность | |
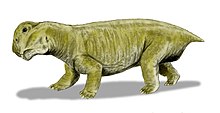
Lystrosaurus (/ˌлɪsтрoʊˈsɔːrəs/; 'совковая ящерица'; правильный греческий - λίστρον Lístron «Инструмент для выравнивания или разглаживания, лопата, лопата, мотыга») был травоядный род из дицинодонт терапсиды от поздняя пермь и Ранний триас эпохи (около 250 миллионов лет назад). Он жил в том, что сейчас Антарктида, Индия, Китай, Монголия, Европейская Россия и Южная Африка. В настоящее время известно от четырех до шести видов, хотя с 1930-х по 1970-е годы считалось, что количество видов намного выше. Они были размером от маленькой собаки до 2,5 метров в длину.[1]
Быть дицинодонт, Lystrosaurus было всего два зуба (пара бивень -подобно клыки ), и считается, что у него был рогатый клюв, который использовался для откусывания кусочков растительности. Lystrosaurus был крепким, травоядным животным, размером примерно с свинья. Строение его плеч и тазобедренных суставов говорит о том, что Lystrosaurus переехал с полуразложенная походка. Передние конечности были даже более крепкими, чем задние, и считается, что животное было мощным землекопом, который гнездился в норах.
Lystrosaurus пережил Пермско-триасовое вымирание, 252 миллиона лет назад. в Ранний триас, они были наиболее распространенными наземными позвоночными, составляя до 95% от общего числа особей в некоторых ископаемых пластах.[2] Исследователи предложили различные гипотезы, почему Lystrosaurus пережил событие вымирания и процветал в раннем триасе.
Описание

Lystrosaurus был дицинодонт терапсид от 0,6 до 2,5 м (от 2 до 8 футов) в длину, в среднем около 0,9 м (3 фута) в зависимости от вида.[3]
В отличие от других терапсидов, дицинодонты имели очень короткие морды и не имели зубов, за исключением бивень -подобный верх клыки. Считается, что дицинодонты имели роговой клювы как черепахи, для срезания кусочков растительности, которые затем растирались на роговой вторичное небо когда рот был закрыт. Сустав челюсти был слабым и двигался вперед и назад с резким движением вместо более обычных движений в стороны или вверх и вниз. Считается, что мышцы челюсти прикреплялись к черепу необычно далеко вперед и занимали много места на верхней и задней части черепа. В результате глаза были посажены высоко и далеко вперед на черепе, а лицо было коротким.[4]
Особенности скелета указывают на то, что Lystrosaurus переехал с полуразложенная походка. Нижний задний угол лопатка (лопатка) была сильно окостенела (построена из прочной кости), что предполагает, что движение лопатки способствовало увеличению длины шага передних конечностей и уменьшало сгибание тела в стороны.[5] Пятерка крестцовые позвонки были массивными, но не слились друг с другом и с таз, делая спину более жесткой и уменьшая изгибы в стороны при ходьбе. Терапсиды с менее чем пятью крестцовыми позвонками, как полагают, имели раскидистые конечности, как у современных ящериц.[5] В динозавры и млекопитающие, имеющих прямые конечности, крестцовые позвонки срослись друг с другом и с таз.[6] Считается, что опора над каждой вертлужной впадиной (тазобедренная впадина) предотвращала вывих из бедренная кость (бедренная кость) пока Lystrosaurus шел полу-раскидистой походкой.[5] Передние конечности Lystrosaurus были массивными,[5] и Lystrosaurus считается, что он был мощным роетелем.
Распространение и виды
Lystrosaurus окаменелости были найдены во многих Поздняя пермь и Ранний триас земной костные ложа, наиболее обильно в Африке и в меньшей степени в некоторых частях нынешней Индии, Китая, Монголии, европейской части России и Антарктиды (которая в то время не находилась над Южным полюсом).[5]
Виды, найденные в Африке

Наиболее Lystrosaurus окаменелости были найдены в Бальфур и Катбергские образования из Кару бассейн в Южная Африка; эти образцы предлагают наилучшие перспективы для идентификации видов, поскольку они наиболее многочисленны и изучаются в течение длительного времени. Как это часто бывает с окаменелостями, в палеонтологическое сообщество относительно того, сколько именно видов было найдено в бассейне Кару.[7] Исследования 1930-1970-х годов показали большое количество (23 в одном случае).[7] Однако к 1980-м и 1990-м годам в Кару было признано только шесть видов: L. curvatus, L. platyceps, L. oviceps, Л. маккайги, L. murrayi, и L. decivis. Исследование 2011 года сократило это число до четырех, рассматривая окаменелости, ранее помеченные как L. platyceps и L. oviceps как члены L. curvatus.[8]
Л. маккайги является самым крупным и, по-видимому, наиболее специализированным видом, в то время как L. curvatus был наименее специализированным. А Lystrosaurus-подобное ископаемое, Kwazulusaurus shakai, также был обнаружен в Южной Африке. Хотя не относится к тому же род, К. шакай очень похож на L. curvatus. Поэтому некоторые палеонтологи предположили, что К. шакай возможно был предком или был тесно связан с предками L. curvatus, пока Л. маккайги возникла из другой линии.[7]Л. маккайги встречается только в отложениях из Пермский период период и, по-видимому, не пережил Пермско-триасовое вымирание. Его особые черты и внезапное появление в Окаменелости без явного предка может указывать на то, что он иммигрировал в Кару из области, в которой не были обнаружены позднепермские отложения.[7]
L. curvatus находится в относительно узкой полосе отложений незадолго до и после вымирания и может использоваться в качестве приблизительного маркера границы между Пермский период и Триасовый периоды. Череп, идентифицированный как L. curvatus найден в позднепермских отложениях из Замбия. В течение многих лет считалось, что не существует пермских образцов L. curvatus в Кару, что привело к предположениям, что L. curvatus иммигрировал из Замбии в Кару. Однако повторное исследование пермских образцов в Кару выявило некоторые из них как L. curvatus, и нет необходимости предполагать иммиграцию.[7]
L. murrayi и L. decivis встречаются только в отложениях триаса.[7]

Другие виды
Lystrosaurus georgi окаменелости были найдены в самых ранних триасовых отложениях Москва Бассейн в России. Вероятно, он был тесно связан с африканским Lystrosaurus curvatus,[5] который считается одним из наименее специализированных видов и был обнаружен в отложениях очень поздней перми и очень раннего триаса.[7]
История окаменелостей

Доктор Элиас Рут Бидл, миссионер из Филадельфии и страстный коллекционер окаменелостей, обнаружил первый Lystrosaurus череп. Бидл написал выдающемуся палеонтологу Отниэль Чарльз Марш, но не получил ответа. Соперник Марша, Эдвард Дринкер Коуп, очень заинтересовался находкой, описал и назвал Lystrosaurus в Труды Американского философского общества в 1870 г.[9] Его название происходит от Древнегреческий слова листрон "лопата" и sauros "ящерица".[10] Марш запоздало приобрел череп в мае 1871 года, хотя его интерес к уже описанному образцу оставался неясным; возможно, он хотел внимательно изучить описание и иллюстрацию Коупа.[9]
Тектоника плит
Открытие Lystrosaurus окаменелости в Угольном утесе в Трансантарктические горы Эдвином Х. Колбертом и его командой в 1969–70 помогли поддержать гипотезу тектоника плит и укрепить теорию, поскольку Lystrosaurus уже были обнаружены в нижнем триасе на юге Африки, а также в Индии и Китае.[11][12]
Палеоэкология
Доминирование раннего триаса
Lystrosaurus отличается преобладанием южных Пангея в течение миллионов лет в течение Ранний триас. По крайней мере один неопознанный вид этого рода пережил массовое вымирание в конце перми и, в отсутствие хищников и травоядных конкурентов, продолжал процветать и повторно излучать на ряд видов в пределах рода,[13] став самой распространенной группой наземных позвоночных в раннем триасе; некоторое время 95% наземных позвоночных были Lystrosaurus.[14][13] Это единственный раз, когда какой-либо один вид или род наземных животных доминировал на Земле в такой степени.[15] Несколько других пермских родов терапсид также пережили массовое вымирание и появились в породах триаса - тероцефалы Тетрацинодон, Moschorhinus и Ictidosuchoides - но, похоже, не были обильными в триасе;[7] полное экологическое восстановление заняло 30 миллионов лет, Рано и Средний триас.[16]
Было сделано несколько попыток объяснить, почему Lystrosaurus пережил Пермско-триасовое вымирание, "мать всех массовых вымираний",[17] и почему он доминировал в фауне раннего триаса в такой беспрецедентной степени:
- Следы роста на окаменелых клыках предполагают, что Lystrosaurus живущий в Антарктиде ~ 250 млн лет назад может войти в длительное состояние оцепенение аналогично спячка. Это могло быть самым старым свидетельством состояния, подобного спячке, у позвоночных животных и указывает на то, что оцепенение возникло у позвоночных до того, как эволюционировали млекопитающие и динозавры.[18][19][20]
- Одна из недавних теорий состоит в том, что в результате вымирания снизилось содержание кислорода в атмосфере и увеличилось ее содержание. углекислый газ содержание, так что многие наземные виды вымерли, потому что им стало слишком трудно дышать.[14] Поэтому было высказано предположение, что Lystrosaurus выжил и стал доминирующим, потому что его роющий образ жизни позволил ему справиться с атмосферой «затхлого воздуха», и что специфические особенности его анатомии были частью этой адаптации: бочкообразная грудь, вмещавшая большие легкие, короткая внутренняя часть ноздри что способствовало учащенному дыханию и высокому нервные отростки (выступы на дорсальной стороне позвонки ), что давало больший толчок мышцам, которые расширялись и сокращались его грудной клеткой. Однако во всех этих пунктах есть слабые места: грудь Lystrosaurus не был значительно больше по сравнению с другими дицинодонты которые вымерли; хотя триасовые дицинодонты, по-видимому, имели более длинные нервные отростки, чем их пермские коллеги, эта особенность может быть связана с позой, движением или даже размером тела, а не с эффективностью дыхания; L. murrayi и L. decivis гораздо более многочисленны, чем другие норные норы раннего триаса, такие как Проколофон или же Тринаксодон.[7]

- Предположение, что Lystrosaurus помог выжить и доминировать, будучи полуводным, имеет аналогичную слабость: хотя темноспондилы становились более многочисленными в триасовых отложениях Кару, их было гораздо меньше, чем L. murrayi и L. decivis.[7]
- Наиболее специализированные и крупные животные подвергаются более высокому риску массового исчезновения; это может объяснить, почему неспециализированные L. curvatus выжили, в то время как более крупные и специализированные Л. маккайги погибли вместе со всеми другими крупными пермскими травоядными и хищниками.[7] Несмотря на то что Lystrosaurus в целом выглядит приспособленным для питания растений, похожих на Дикроидиум, который доминировал в раннем триасе, более крупные размеры Л. маккайги возможно, вынудили его полагаться на более крупных членов Глоссоптерис флора, которая не пережила конец пермского вымирания.[7]
- Только тероцефал длиной 1,5 м (5 футов) Moschorhinus и большой архозаврический Протерозух кажутся достаточно большими, чтобы охотиться на триасовый период Lystrosaurus видов, и эта нехватка хищников могла быть причиной Lystrosaurus демографический бум в раннем триасе.[7]
- По словам Бентона, «возможно, выживание Lystrosaurus просто удача ".[13]
Смотрите также
- Эволюция млекопитающих - Получение млекопитающих от предшественника синапсидов и адаптивное излучение видов млекопитающих
- Списки синапсидов - Статья со списком Википедии
Рекомендации
- ^ Клувер, Майкл Альберт (1978). Ископаемые рептилии южноафриканского кару. Южноафриканский музей. ISBN 9780908407583.
- ^ Damiani, R.J .; Neveling, J .; Модесто С.П. и Йейтс А.М. (2004). "Барендскраал, разнообразный район амниот из Lystrosaurus зона ассоциаций раннего триаса Южной Африки ». Palaeontologia Africana. 39: 53–62.
- ^ "Листрозавр | Ископаемые и факты". Энциклопедия Британника. Получено 2019-03-18.
- ^ Коуэн, Р. (2000). История жизни (3-е изд.). Blackwell Scientific. С. 167–68. ISBN 978-0-632-04444-3.
- ^ а б c d е ж Сурков, М.В .; Каландадзе, Н. Оф & Бентон, М.Дж. (июнь 2005 г.). "Lystrosaurus georgi, дицинодонт из нижнего триаса России » (PDF). Журнал палеонтологии позвоночных. 25 (2): 402–413. Дои:10.1671 / 0272-4634 (2005) 025 [0402: LGADFT] 2.0.CO; 2. ISSN 0272-4634. Архивировано из оригинал (PDF) на 2008-12-19. Получено 2008-07-07.
- ^ Бентон, Майкл Дж. (2004). «Происхождение и родство динозавров». В Weishampel, David B .; Додсон, Питер и Осмольска Хальска (ред.). Динозаврия (2-е изд.). Беркли: Калифорнийский университет Press. стр.7 –19. ISBN 978-0-520-24209-8.
- ^ а б c d е ж грамм час я j k л м Бота, Дж. И Смит, Р.М.Х. (2005). "Lystrosaurus видовой состав через пермо-триасовую границу в бассейне Кару в Южной Африке ». Lethaia. 40 (2): 125–137. Дои:10.1111 / j.1502-3931.2007.00011.x. Полная версия онлайн на "Lystrosaurus видовой состав через пермо-триасовую границу в бассейне Кару в Южной Африке » (PDF). Архивировано из оригинал (PDF) на 2008-09-10. Получено 2008-07-02.
- ^ Grine, F.E .; Forster, C.A .; Клувер, М.А., Георги, Дж. А. (2006). Палеобиология амниот. Перспективы эволюции млекопитающих, птиц и рептилий. Издательство Чикагского университета. С. 432–503.
- ^ а б Уоллес, Дэвид Рейнс (2000). Месть костохотчиков: динозавры, жадность и величайшая научная вражда позолоченного века. Houghton Mifflin Harcourt. С. 44–45. ISBN 978-0-618-08240-7.
- ^ Лидделл, Генри Джордж & Роберт Скотт (1980). Греко-английский лексикон (Сокращенное изд.). Объединенное Королевство: Oxford University Press. ISBN 978-0-19-910207-5.
- ^ Наоми Любик, Изучение Антарктики, Geotimes, 2005.
- ^ Трюик, Стив (2016). «Тектоника плит в биогеографии». Международная энциклопедия географии: люди, земля, окружающая среда и технологии. John Wiley & Sons, Ltd., стр. 1–9. Дои:10.1002 / 9781118786352.wbieg0638. ISBN 9781118786352.
- ^ а б c Майкл Дж. Бентон (2006). Когда жизнь почти умерла. Величайшее массовое вымирание в истории. Лондон: Темза и Гудзон. ISBN 978-0-500-28573-2.
- ^ а б [1] В архиве 12 октября 2007 г. Wayback Machine
- ^ BBC: Жизнь до динозавров
- ^ Сахни, С. и Бентон, М.Дж. (2008). «Восстановление после самого глубокого массового вымирания всех времен» (PDF). Труды Королевского общества B: биологические науки. 275 (1636): 759–65. Дои:10.1098 / rspb.2007.1370. ЧВК 2596898. PMID 18198148.
- ^ Эрвин Д.Х. (1993). Великий палеозойский кризис; Жизнь и смерть в перми. Издательство Колумбийского университета. ISBN 978-0-231-07467-4.
- ^ «Ископаемые свидетельства состояния« спячки »у антарктических животных возрастом 250 миллионов лет». Phys.org. Получено 7 сентября 2020.
- ^ «Ископаемые остатки предполагают, что животные впали в спячку уже 250 миллионов лет». UPI. Получено 7 сентября 2020.
- ^ Уитни, Меган Р .; Сидор, Кристиан А. (27 августа 2020 г.). «Свидетельства оцепенения бивней Lystrosaurus из раннего триаса Антарктиды». Биология коммуникации. 3 (1): 471. Дои:10.1038 / с42003-020-01207-6. ISSN 2399-3642. ЧВК 7453012. PMID 32855434.
 Текст и изображения доступны под Международная лицензия Creative Commons Attribution 4.0.
Текст и изображения доступны под Международная лицензия Creative Commons Attribution 4.0.
внешняя ссылка
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||





