Ксилема - Xylem

Ксилема один из двух видов транспорта ткань в сосудистые растения, флоэма быть другим. Основная функция ксилемы - транспортировать воды от корней к стеблям и листьям, но также переносит питательные вещества.[1][2] Слово «ксилема» происходит от Греческий слово ξύλον (ксилон), что означает «дерево»; самая известная ткань ксилемы - это дерево, хотя он встречается повсюду в растении.[3] Термин был введен Карл Нэгели в 1858 г.[4][5]
Структура

Самая отличительная ксилема клетки представляют собой длинные трахеальные элементы, переносящие воду. Трахеиды и элементы судна отличаются своей формой; элементы сосуда короче и соединены в длинные трубки, которые называются сосуды.[6]
Ксилема также содержит два других типа клеток: паренхима и волокна.[7]
Ксилему можно найти:
- в сосудистые пучки, присутствует в недревесных растениях и недревесных частях древесных растений
- во вторичной ксилеме, заложенной меристема называется сосудистый камбий в древесных растениях
- как часть стеллярная композиция не разделены на связки, как во многих папоротники.
На переходных стадиях растений с вторичный рост, первые две категории не исключают друг друга, хотя обычно сосудистый пучок содержит первичная ксилема только.
Паттерн ветвления, демонстрируемый ксилемой, следует Закон Мюррея.[8]
Первичная и вторичная ксилема
Первичная ксилема образуется при первичном росте из прокамбий. Он включает протоксилему и метаксилему. Метаксилема развивается после протоксилемы, но до вторичной ксилемы. Метаксилема имеет более широкие сосуды и трахеиды, чем протоксилема.
Вторичная ксилема образуется при вторичном росте из сосудистый камбий. Хотя вторичная ксилема также обнаруживается у представителей голосеменное растение группы Гнетофита и Гинкгофита и в меньшей степени у членов Cycadophyta, две основные группы, в которых может быть обнаружена вторичная ксилема:
- хвойные породы (Хвойные): известно около 600 видов хвойных пород.[9] Все виды имеют вторичную ксилему, которая относительно однородна по структуре для всей этой группы. Многие хвойные деревья становятся высокими деревьями: вторичная ксилема таких деревьев используется и продается как мягкая древесина.
- покрытосеменные (Покрытосеменные): их примерно 250 000[9] известные виды покрытосеменных. Внутри этой группы вторичная ксилема встречается редко. однодольные.[10] Многие немонодольные покрытосеменные становятся деревьями, и их вторичная ксилема используется и продается как твердая древесина.
Основная функция - водный транспорт вверх
Ксилема, сосуды и трахеиды корней, стеблей и листьев соединены между собой, образуя непрерывную систему водопроводящих каналов, достигающих всех частей растений. Система транспортирует воду и растворимые минеральные питательные вещества от корней по всему растению. Он также используется для восполнения потери воды во время испарение и фотосинтез. Ксилема сок состоит в основном из воды и неорганических ионов, хотя может также содержать ряд органических химикатов. Транспорт пассивный, не питается от энергии, затрачиваемой трахея сами элементы, которые умерли по зрелости и больше не имеют живого содержания. Транспортировка сока вверх становится более трудной, поскольку высота растения увеличивается, и считается, что транспортировка воды вверх по ксилеме ограничивает максимальную высоту деревьев.[11] Три явления вызывают выделение ксилемного сока:
- Гипотеза потока давления: Сахар, производимый листьями и другими зелеными тканями, сохраняется во флоэме, создавая давление растворенного вещества дифференциал по сравнению с ксилемной системой, несущей гораздо меньшую нагрузку растворенных веществ - воды и минералов. Давление флоэмы может достигать нескольких МПа,[12] намного выше атмосферного давления. Селективная взаимосвязь между этими системами позволяет этой высокой концентрации растворенного вещества во флоэме вытягивать ксилемную жидкость вверх за счет отрицательного давления.
- Транспирационное притяжение: Аналогично испарение из воды с поверхностей мезофилл ячеек в атмосферу также создает отрицательное давление в верхней части растения. Это вызывает миллионы минут мениски образовываться в клеточной стенке мезофилла. Результирующий поверхностное натяжение вызывает отрицательное давление или напряжение в ксилеме, которая вытягивает воду из корней и почвы.
- Корневое давление: Если водный потенциал клеток корня более отрицательный, чем почва, обычно из-за высоких концентраций растворенное вещество, вода может двигаться осмос в корень из почвы. Это вызывает положительное давление, которое заставляет ксилему двигаться по направлению к листьям. В некоторых случаях сок будет вытеснен из листа через гидатод в явлении, известном как потоотделение. Самое высокое давление на корни приходится на утро, когда устьица открываются и позволяют начать транспирацию. У разных видов растений давление на корни может быть разным даже в одинаковых условиях; примеры включают до 145 кПа в Vitis riparia но около нуля в Celastrus orbiculatus.[13]
Основная сила, которая создает капиллярное действие движение воды вверх у растений - это адгезия между водой и поверхностью ксилемных каналов.[14][15] Капиллярное действие обеспечивает силу, которая устанавливает равновесную конфигурацию, уравновешивая силу тяжести. Когда транспирация удаляет воду из верхней части, поток необходим, чтобы вернуться к равновесию.
Переносная тяга возникает в результате испарения воды с поверхностей клетки в уходит. Это испарение заставляет поверхность воды погружаться в поры из клеточная стенка. От капиллярное действие вода образует вогнутую мениски внутри пор. Высокое поверхностное натяжение воды тянет вогнутость наружу, производя достаточно сила поднимать воду на высоту до ста метров от уровня земли до дерево высшие ветви.
Транспирационная тяга требует, чтобы сосуды, транспортирующие воду, были очень маленькими в диаметре; в противном случае, кавитация сломает толщу воды. И как вода испаряется из листьев через растение вытягивается больше, чтобы заменить его. Когда давление воды внутри ксилемы достигает экстремального уровня из-за низкого поступления воды из корней (например, если почва сухая), газы выходят из раствора и образуют пузырь - эмболия формы, которые быстро распространятся на другие соседние ячейки, если окаймленные ямы присутствуют (они имеют структуру, похожую на заглушку, называемую тором, которая закрывает отверстие между соседними клетками и останавливает распространение эмболии). Даже после того, как произошла эмболия, растения могут восполнить ксилему и восстановить ее функциональность.[16]
Теория когезии-напряжения
В теория сцепления-напряжения это теория из межмолекулярное притяжение это объясняет процесс воды течь вверх (против силы сила тяжести ) через ксилему растения. Он был предложен в 1894 г. Джон Джоли и Генри Горацио Диксон.[17][18] Несмотря на многочисленные возражения,[19][20] это наиболее широко распространенная теория переноса воды через сосудистую систему растений, основанная на классических исследованиях Диксона-Джоли (1894 г.), Евгения Аскенасси (1845–1903 гг.) (1895 г.),[21][22] и Диксон (1914, 1924).[23][24]
Вода - это полярная молекула. Когда две молекулы воды сближаются, слегка отрицательно заряженные кислород атом одного образует водородная связь со слегка положительно заряженным водород атом в другом. Эта сила притяжения, наряду с другими межмолекулярные силы, является одним из основных факторов, ответственных за возникновение поверхностное натяжение в жидкой воде. Это также позволяет растениям вытягивать воду от корня через ксилему к листу.
Вода постоянно теряется из-за испарения листа. Когда одна молекула воды теряется, другую затягивают процессы сцепления и напряжения. Тяга транспирации, используя капиллярное действие и внутреннее поверхностное натяжение воды, является основным механизмом движения воды в растениях. Однако это не единственный задействованный механизм. Любое использование воды в листьях заставляет воду перемещаться в них.
Транспирация в листьях создает напряжение (перепад давления) в стенках ячеек мезофилл клетки. Из-за этого напряжения вода поднимается от корней к листьям, чему способствует сплоченность (притяжение между отдельными молекулами воды из-за водородные связи ) и адгезия (липкость между молекулами воды и гидрофильный клеточные стенки растений). Этот механизм потока воды работает из-за водный потенциал (вода течет от высокого к низкому потенциалу), и правила простого распространение.[25]
За последнее столетие было проведено множество исследований механизма транспорта ксилемного сока; сегодня большинство ученых-растениеводов продолжают соглашаться с тем, что теория когезии-напряжения лучше всего объясняет этот процесс, но были предложены теории множественных сил, которые предполагают несколько альтернативных механизмов, включая продольные клеточные и ксилемные осмотическое давление градиенты, осевые градиенты потенциала в сосудах и межфазные градиенты, поддерживаемые гелем и газовыми пузырьками.[26][27]
Измерение давления

До недавнего времени перепад давления (всасывания) транспирационной тяги можно было измерить только косвенно, путем приложения внешнего давления с бомба высокого давления чтобы противодействовать этому.[28] Когда была разработана технология выполнения прямых измерений с помощью датчика давления, первоначально возникли некоторые сомнения в правильности классической теории, поскольку некоторые исследователи не смогли продемонстрировать отрицательное давление. Более поздние измерения, по большей части, подтверждают классическую теорию. Транспортировка ксилемы осуществляется за счет комбинации[29] транспирационного притяжения сверху и корневое давление снизу, что усложняет интерпретацию измерений.
Эволюция
Ксилема появилась в начале истории наземных растений. Ископаемые растения с анатомически сохранившейся ксилемой известны с Силурийский (более 400 миллионов лет назад), а ископаемые остатки, напоминающие отдельные клетки ксилемы, могут быть найдены в более ранних Ордовик горные породы.[нужна цитата ] Самая ранняя настоящая и узнаваемая ксилема состоит из трахеиды с добавлением спирально-кольцевого армирующего слоя клеточная стенка. Это единственный тип ксилемы, обнаруженный у самых ранних сосудистых растений, и этот тип клеток по-прежнему встречается в протоксилема (первообразованная ксилема) всех живых групп сосудистых растений. Позже развились несколько групп растений. без ямок клетки трахеиды независимо через конвергентная эволюция. У живых растений трахеиды без косточек не появляются в развитии до созревания метаксилема (после протоксилема).
У большинства растений без косточек трахеиды функционируют как первичные транспортные клетки. Другой тип сосудистого элемента, обнаруженный у покрытосеменных, - это элемент сосуда. Элементы сосуда соединяются встык, образуя сосуды, в которых вода течет беспрепятственно, как в трубе. Наличие ксилемных сосудов считается одним из ключевых нововведений, которые привели к успеху покрытосеменные.[30] Однако наличие сосудистых элементов не ограничивается покрытосеменными, и они отсутствуют в некоторых архаичных или «базальных» ветвях покрытосеменных: (например, Amborellaceae, Тетрацентровые, Trochodendraceae, и Winteraceae ), а их вторичная ксилема описывается Артур Кронквист как «примитивно бессосудистые». Кронквист считал сосуды Gnetum сходиться с таковыми у покрытосеменных.[31] Является ли отсутствие сосудов у базальных покрытосеменных примитивный Состояние оспаривается, альтернативная гипотеза утверждает, что элементы сосудов произошли от предшественников покрытосеменных и впоследствии были потеряны.

Для фотосинтеза растения должны поглощать CO
2 из атмосферы. Однако за это приходится платить: хотя устьицы открыты, чтобы позволить CO
2 при входе вода может испариться.[32] Вода теряется намного быстрее, чем CO
2 абсорбируется, поэтому растениям необходимо его заменить, и были разработаны системы для транспортировки воды из влажной почвы к месту фотосинтеза.[32] Ранние растения засасывали воду между стенками своих клеток, а затем развили способность контролировать потерю воды (и CO
2 приобретение) через устьица. Специализированные водные транспортные ткани вскоре превратились в гидроидов, трахеид, затем вторичную ксилему, затем энтодерму и, наконец, сосуды.[32]
Высота CO
2 Уровни силурийско-девонского периода, когда растения впервые заселяли землю, означали, что потребность в воде была относительно низкой. Так как CO
2 был выведен из атмосферы растениями, при его захвате было потеряно больше воды, и возникли более элегантные транспортные механизмы.[32] По мере развития механизмов переноса воды и водонепроницаемой кутикулы растения могли выживать, не будучи постоянно покрытыми водной пленкой. Этот переход от пойкилогидрия к гомойогидрия открыл новый потенциал для колонизации.[32] Затем растениям нужна была прочная внутренняя структура, которая содержала длинные узкие каналы для транспортировки воды из почвы во все различные части надземного растения, особенно в те части, где происходил фотосинтез.
В силурийское время CO
2 был легко доступен, поэтому для его приобретения требовалось мало воды. К концу карбона, когда CO
2 уровень снизился примерно до сегодняшнего, примерно в 17 раз больше воды терялось на единицу CO
2 поглощение.[32] Однако даже в эти «легкие» первые дни вода была в цене, и ее приходилось доставлять к частям растения из влажной почвы, чтобы избежать высыхание. Этот ранний водный транспорт использовал преимущества сцепление-напряжение механизм, присущий воде. Вода имеет тенденцию диффундировать в более сухие области, и этот процесс ускоряется, когда вода может быть злой по ткани с небольшими промежутками. В небольших проходах, например, между стенками растительных клеток (или в трахеидах), столб воды ведет себя как резина - когда молекулы испаряются с одного конца, они тянут молекулы за собой по каналам. Следовательно, только транспирация является движущей силой переноса воды у ранних растений.[32] Однако без специальных транспортных сосудов механизм сцепления-натяжения не может транспортировать воду более чем на 2 см, что сильно ограничивает размер самых ранних растений.[32] Этот процесс требует постоянной подачи воды с одного конца для поддержания цепей; чтобы не утомлять его, растения разработали водонепроницаемую кутикула. Ранняя кутикула могла не иметь пор, но не покрывала всю поверхность растения, так что газообмен продолжался.[32] Однако иногда обезвоживание было неизбежным; Ранние растения справляются с этим, храня много воды между клеточными стенками, и, когда дело доходит до этого, выдерживают трудные времена, задерживая жизнь, пока не поступит больше воды.[32]

Чтобы быть свободными от ограничений небольшого размера и постоянной влажности, которые создавала паренхиматическая транспортная система, растениям требовалась более эффективная водная транспортная система. В течение ранний силурийский, они разработали специализированные клетки, которые были одревесневший (или имели аналогичные химические соединения)[32] чтобы избежать взрыва; этот процесс совпал с гибелью клеток, позволяя опустошить их внутренности и пропустить через них воду.[32] Эти более широкие, мертвые, пустые клетки были в миллион раз более проводящими, чем межклеточный метод, что давало возможность переноса на большие расстояния и более высокие CO
2 скорости диффузии.
Самые ранние макрофоссилии, несущие водопроводные трубки, - это силурийские растения, относящиеся к роду Cooksonia.[33] Раннедевонские претрахеофиты Аглаофитон и Хорнеофитон имеют структуру, очень похожую на гидроиды современных мхов. Растения продолжали изобретать новые способы уменьшения сопротивления потоку внутри своих ячеек, тем самым повышая эффективность водного транспорта. Полосы на стенках трубок, заметные начиная с раннего силурия,[34] это ранняя импровизация, помогающая плавно течь воды.[35] Ленточные трубки, а также трубки с ямчатым орнаментом на стенках одревесневали.[36] и, когда они образуют одноклеточные каналы, считаются трахеиды. Эти конструкции транспортных ячеек «следующего поколения» имеют более жесткую конструкцию, чем гидроиды, что позволяет им справляться с более высокими уровнями давления воды.[32] Трахеиды могут иметь единственное эволюционное происхождение, возможно, в пределах роголистников,[37] объединение всех трахеофитов (но они могли эволюционировать более одного раза).[32]
Водный транспорт требует регулирования, а динамическое управление обеспечивается устьица.[38]Регулируя количество газообмена, они могут ограничить количество воды, теряемой при транспирации. Это важная роль там, где водоснабжение не является постоянным, и действительно, кажется, что устьица развились до трахеид и присутствуют у несосудистых роголистников.[32]
An энтодерма вероятно, образовались в течение Силу-девона, но первые ископаемые свидетельства такой структуры относятся к каменноугольному периоду.[32] Эта структура в корнях покрывает ткань, транспортирующую воду, и регулирует ионный обмен (и предотвращает попадание нежелательных патогенов и т. Д. В систему транспорта воды). Энтодерма также может оказывать давление вверх, вытесняя воду из корней, когда транспирации недостаточно.
Когда растения развили этот уровень контролируемого переноса воды, они стали действительно гомоатомными, способны извлекать воду из окружающей среды через корнеподобные органы, а не полагаться на пленку поверхностной влаги, что позволило им вырасти до гораздо большего размера.[32] В результате своей независимости от своего окружения они потеряли способность выживать при высыхании - дорогостоящая черта для сохранения.[32]
В течение девона максимальный диаметр ксилемы увеличивался со временем, а минимальный диаметр оставался довольно постоянным.[35] К среднему девону диаметр трахеид некоторых линий растений (Зостерофиллофиты ) вышла на плато.[35] Более широкие трахеиды позволяют воде транспортироваться быстрее, но общая скорость транспортировки зависит также от общей площади поперечного сечения самого пучка ксилемы.[35] Увеличение толщины сосудистых пучков, по-видимому, также коррелирует с шириной осей растений и высотой растений; это также тесно связано с появлением листьев[35] и повышенная плотность устьиц, что увеличивает потребность в воде.[32]
В то время как более широкие трахеиды с прочными стенками позволяют достичь более высокого давления переноса воды, это увеличивает проблему кавитации.[32] Кавитация возникает, когда внутри сосуда образуется пузырь воздуха, разрывая связи между цепочками молекул воды и не позволяя им поднимать больше воды за счет своего когезионного натяжения. После кавитации из трахеиды нельзя удалить эмболию и вернуть ее в рабочее состояние (за исключением нескольких развитых покрытосеменных растений).[39][40] которые разработали такой механизм). Поэтому растениям стоит избегать кавитации. Именно по этой причине, ямы в стенках трахеиды имеют очень маленький диаметр, чтобы предотвратить попадание воздуха и образование пузырьков. Циклы замораживания-оттаивания являются основной причиной кавитации. Повреждение стенки трахеиды почти неизбежно приводит к утечке воздуха и кавитации, отсюда важность параллельной работы многих трахеид.[32]
Кавитации трудно избежать, но, как только она возникла, у растений есть ряд механизмов для сдерживания повреждений.[32] Маленькие ямки соединяют соседние каналы, позволяя жидкости течь между ними, но не воздуху - хотя по иронии судьбы эти ямки, которые предотвращают распространение эмболий, также являются их основной причиной.[32] Эти покрытые ямками поверхности дополнительно уменьшают поток воды через ксилему на 30%.[32] Хвойные породы юрского периода разработали гениальное усовершенствование, используя клапанные структуры для изоляции кавитированных элементов. Эти структуры тор-марго имеют каплю, плавающую в середине пончика; когда одна сторона сбрасывает давление, капля засасывается в тор и блокирует дальнейший поток.[32] Другие растения просто принимают кавитацию; например, в начале каждой весны у дубов вырастает кольцо из широких сосудов, ни одна из которых не выдерживает зимних заморозков. Каждую весну клены используют давление корней, чтобы выдавливать сок вверх от корней, выдавливая любые пузырьки воздуха.
Рост до высоты также использовал другую черту трахеид - поддержку, обеспечиваемую их одревесневшими стенками. Отмершие трахеиды были сохранены, чтобы сформировать прочный древесный стебель, в большинстве случаев производимый вторичной ксилемой. Однако у ранних растений трахеиды были слишком механически уязвимы и сохраняли центральное положение со слоем жесткой склеренхима по внешнему ободку стеблей.[32] Даже когда трахеиды действительно играют структурную роль, они поддерживаются склеренхимной тканью.
Трахеиды заканчиваются стенками, которые оказывают большое сопротивление потоку;[35] Элементы емкости имеют перфорированные торцевые стенки и расположены последовательно для работы, как если бы они были одним непрерывным резервуаром.[35] Функция торцевых стен, которые были стандартным состоянием в Девоне, вероятно, заключалась в том, чтобы избежать эмболии. Эмболия - это образование воздушного пузыря в трахеиде. Это может произойти в результате замерзания или растворения газов из раствора. Как только образовалась эмболия, ее обычно нельзя удалить (но см. Ниже); пораженная клетка не может поднимать воду и становится бесполезной.
За исключением торцевых стенок, трахеиды преваскулярных растений могли работать при той же гидравлической проводимости, что и у первого сосудистого растения. Cooksonia.[35]
Размер трахеид ограничен, поскольку они состоят из одной клетки; это ограничивает их длину, что, в свою очередь, ограничивает их максимальный полезный диаметр до 80 мкм.[32] Электропроводность растет в четвертой степени диаметра, поэтому увеличение диаметра имеет огромные преимущества; элементы судна, состоящий из ряда ячеек, соединенных на концах, преодолел этот предел и позволил сформировать более крупные трубки, достигающие диаметра до 500 мкм и длины до 10 м.[32]
Сосуды впервые образовались во время сухого, низкого CO
2 периоды поздней перми, в хвощах, папоротниках и Selaginellales независимо, а позже появился в среднем меловом периоде у покрытосеменных и гнетофитов.[32]Сосуды позволяют древесине той же площади поперечного сечения транспортировать примерно в сто раз больше воды, чем трахеиды![32] Это позволило растениям заполнить больше стеблей структурными волокнами, а также открыло новую нишу для лозы, который мог переносить воду, не будучи таким толстым, как дерево, на котором они выросли.[32] Несмотря на эти преимущества, древесина на основе трахеид намного легче, а значит, дешевле в производстве, поскольку сосуды должны быть значительно усилены, чтобы избежать кавитации.[32]
Развитие
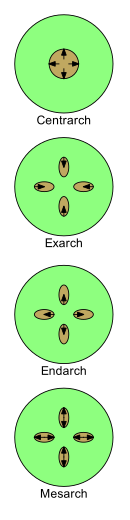
Развитие ксилемы можно описать четырьмя терминами: центрарх, экзарх, эндарх и мезарх. По мере развития у молодых растений характер его меняется от протоксилема к метаксилема (т.е. из первая ксилема к после ксилемы). Паттерны, в которых расположены протоксилема и метаксилема, важны при изучении морфологии растений.
Протоксилема и метаксилема
Как молодой сосудистое растение В его стеблях и корнях образуются одна или несколько нитей первичной ксилемы. Первая развивающаяся ксилема называется «протоксилема». По внешнему виду протоксилема обычно отличается более узкими сосудами, образованными более мелкими клетками. Некоторые из этих клеток имеют стенки, содержащие утолщения в виде колец или спиралей. Функционально протоксилема может расширяться: клетки могут увеличиваться в размерах и развиваться, в то время как стебель или корень удлиняются. Позже «метаксилема» развивается в тяжах ксилемы. Сосуды и клетки метаксилемы обычно больше; ячейки имеют утолщения, которые обычно имеют форму лестничных поперечных стержней (скалярная форма) или сплошных листов, за исключением отверстий или ямок (с ямками). Функционально метаксилема завершает свое развитие после прекращения удлинения, когда клетки больше не нуждаются в увеличении размера.[41][42]
Паттерны протоксилемы и метаксилемы
Существует четыре основных образца расположения протоксилемы и метаксилемы в стеблях и корнях.
- Centrarch относится к случаю, когда первичная ксилема образует единый цилиндр в центре стебля и развивается от центра кнаружи. Таким образом, протоксилема находится в центральном ядре, а метаксилема - в цилиндре вокруг него.[43] Этот образец был распространен на ранних наземных растениях, таких как "риниофиты ", но не присутствует ни в каких живых растениях.[нужна цитата ]
Остальные три термина используются там, где имеется более одной нити первичной ксилемы.
- Экзарх используется, когда в стебле или корне имеется более одной нити первичной ксилемы, и ксилема развивается снаружи внутрь к центру, то есть центростремительно. Таким образом, метаксилема находится ближе всего к центру стебля или корня, а протоксилема - к периферии. Корни сосудистые растения обычно считаются экзархами.[41]
- Endarch используется, когда в стебле или корне имеется более одной нити первичной ксилемы, и ксилема развивается изнутри наружу к периферии, то есть центробежно. Таким образом, протоксилема находится ближе всего к центру стебля или корня, а метаксилема - к периферии. Стебли семенные растения обычно имеют эндархическое развитие.[41]
- Мезарх используется, когда в стебле или корне имеется более одной нити первичной ксилемы, и ксилема развивается из середины нити в обоих направлениях. Таким образом, метаксилема находится как на периферической, так и на центральной сторонах цепи, а протоксилема находится между метаксилемой (возможно, окруженной ею). Листья и стебли многих папоротники есть разработка mesarch.[41]
История
В его книге Растение либри XVI (О растениях, в 16 книгах) (1583), итальянский врач и ботаник. Андреа Чезальпино предложил растения черпать воду из почвы не за счет магнетизма (ut magnes ferrum trahit, поскольку магнитное железо притягивает), ни всасыванием (вакуум), но путем абсорбции, как в случае с полотном, губками или порошками.[44] Итальянский биолог Марчелло Мальпиги был первым, кто описал и проиллюстрировал ксилемные сосуды, что он и сделал в своей книге Анатом подошвенный ... (1675).[45][примечание 1] Хотя Мальпиги считал, что ксилема содержит только воздух, британский врач и ботаник Неемия Рос, который был современником Мальпиги, считал, что сок поднимается как через кору, так и через ксилему.[46] Однако, по словам Грю, капиллярное действие в ксилеме сок поднимется всего на несколько дюймов; Чтобы поднять сок на верхушку дерева, Грю предположил, что паренхимные клетки становятся набухшими и, таким образом, не только сжимают сок в трахеидах, но и выталкивают некоторое количество сока из паренхимы в трахеиды.[47] В 1727 г. английский священник и ботаник. Стивен Хейлз показали, что транспирация листьями растения заставляет воду перемещаться через его ксилему.[48][заметка 2] К 1891 г. польско-немецкий ботаник Эдуард Страсбургер показали, что транспорт воды в растениях не требует, чтобы клетки ксилемы были живыми.[49]
Смотрите также
- Флоэма
- Континуум атмосферы почвенного растения
- Стела
- Всасывание
- Тилоз
- Сосудистый пучок
- Сосудистая ткань
- Ксилемный сок
Пояснительные примечания
- ^ Мальпиги впервые описал сосуды ксилемы и назвал трахеидные клетки. С п. 8 из (Мальпиги, 1675 г.): "… Haec tubulosa sunt & subrotunda, Identidem tamen angustantur, & perpetuo патент, nullumque, ut observare potui, effundunt humorem: Argentea lamina L, in spiram contorta, components, ut facile laceratione, (velut in bombycis, tracheis expert") продолговатый и континуатам фасции резолвантур. Lamina haec, si ulterius microscopio lustretur, specificulis squamatim Component; quod etiam in tracheis insctorum deprehenditur. robur, transversalium utriculorum ordines N superequitant, ita ut fiat veluti storea ". (… Эти [сосуды] трубчатые и несколько круглые, но часто становятся узкими, и они всегда открыты, и ни один, насколько [насколько] я мог понять, не выделяет жидкости: они состоят из серебристых пластин. L, скрученные в спираль, хотя их можно легко развязать, разорвав в эту несколько длинную и связанную полоску (как я сделал в случае с триахами тутового шелкопряда). Этот лист, если его дополнительно изучить под микроскопом, состоит из чешуевидных частиц; что также наблюдается в трахее насекомых. На этих винтовых сосудах, или, правильнее сказать, «трахее», стоят древесные нити. M, который удлиняется по длине поперечно - для большей прочности и твердости - линии поперечных ячеек N, так что он построен как циновка.)
- ^ Хейлз объяснил, что, хотя капиллярное действие может способствовать подъему воды в ксилеме, транспирация заставляет воду действительно перемещаться через ксилему. Из (Хейлз, 1727), стр. 100: «И по тому же [капиллярному] принципу, что мы видим в предыдущих экспериментах, растения так энергично впитывают влагу в свои тонкие капиллярные сосуды; эта влага, поскольку она уносится с потом [то есть транспирацией], (посредством действие тепла), тем самым дает возможность сосудам для сока почти постоянно притягивать свежие запасы, чего они не могли бы сделать, если бы были полностью насыщены влагой: ибо без потоотделения сок обязательно должен застаиваться, несмотря на то, что сосуды для сока так странно приспособленные своей необычайной крупностью, чтобы поднять сок на большую высоту, пропорциональную их очень маленькому диаметру ».
использованная литература
Цитаты
- ^ Перселл, Адам. «Ксилема и флоэма». Основы биологии. В архиве из оригинала от 04.05.2016.
- ^ Кейт Робертс, изд. (2007). Справочник по растениеводству. 1 (иллюстрированный ред.). Джон Вили и сыновья. п. 185. ISBN 9780470057230.
- ^ Ричард Б. Манке (1977). Обеспечение энергии: доклад Целевой группы Фонда двадцатого века по энергетической политике США (иллюстрированный ред.). Тата Макгроу-Хилл Образование. п.42. ISBN 9780070656178.
- ^ Нэгели, Карл (1858). "Das Wachstum des Stammes und der Wurzel bei den Gefäßpflanzen und die Anordnung der Gefäßstränge im Stengel" [Рост стебля и корня среди сосудистых растений и расположение сосудистых нитей в стебле]. Beiträge zur Wissenschaftlichen Botanik (Вклад в научную ботанику) (на немецком). 1: 1–156. С п. 9: «Ich die beiden Partien Dauergewebe, welche von dem Cambium nach aussen und nach innen gebildet werden, Phloëm und Xylem nennen». (Я назову две части постоянной ткани, которые образованы камбием снаружи и внутри, «флоэма» и «ксилема».)
- ^ Буват, Роджер (1989). «Флоэма». Онтогенез, дифференцировка клеток и структура сосудистых растений. С. 287–368. Дои:10.1007/978-3-642-73635-3_10. ISBN 978-3-642-73637-7.
- ^ Рэйвен, Питер А .; Эверт, Рэй Ф. и Эйххорн, Сьюзан Э. (1999). Биология растений. W.H. Фримен и компания. С. 576–577. ISBN 978-1-57259-611-5.
- ^ Ксилема В архиве 2011-09-16 на Wayback Machine. Британская энциклопедия
- ^ McCulloh, Katherine A .; Джон С. Сперри; Фредерик Р. Адлер (2003). «Водный транспорт на заводах подчиняется закону Мюррея». Природа. 421 (6926): 939–942. Bibcode:2003Натура 421..939М. Дои:10.1038 / природа01444. PMID 12607000. S2CID 4395034.
- ^ а б Уолтер С. Джадд (2002). Уолтер С. Джадд (ред.). Систематика растений: филогенетический подход (2-е изд.). ISBN 0-87893-403-0.
- ^ Дикисон, W.C. (2000). Интегративная анатомия растений (страница 196). Elsevier Science. ISBN 9780080508917. В архиве из оригинала от 06.11.2017.
- ^ Кох, Джордж У .; Силлетт, Стивен С.; Дженнингс, Грегори М .; Дэвис, Стивен Д. (2004). «Пределы высоты дерева». Природа. 428 (6985): 851–854. Bibcode:2004Натура.428..851K. Дои:10.1038 / природа02417. PMID 15103376. S2CID 11846291.
- ^ Knoblauch, Майкл; Кноблаух, Ян; Mullendore, Daniel L .; Сэвидж, Джессика А .; Бабст, Бенджамин А .; Beecher, Sierra D .; Dodgen, Adam C .; Jensen, Kaare H .; Холбрук, Н. Мишель (02.06.2016). «Проверка гипотезы Мюнха о переносе флоэмы на большие расстояния в растениях». eLife. 5: e15341. Дои:10.7554 / eLife.15341. ISSN 2050-084X. ЧВК 4946904. PMID 27253062.
- ^ Тим Дж. Тиббетс; Фрэнк У. Эверс (2000). «Давление корней и удельная проводимость лиан умеренного пояса: экзотика. Celastrus orbiculatus (Celastraceae) против коренных Vitis riparia (Vitaceae) ". Американский журнал ботаники. 87 (9): 1272–78. Дои:10.2307/2656720. JSTOR 2656720. PMID 10991898. В архиве из оригинала от 12.10.2007.
- ^ Крузиа, Пьер и Рихтер, Ханно. Физиология растений В архиве 2008-12-28 на Wayback Machine. Sinauer Associates.
- ^ Энтони Р. Йео; Тимоти Дж. Флауэрс, ред. (2007). Транспорт растворенных веществ растений. Оксфорд, Великобритания: Blackwell Publishing. п.221. ISBN 978-1-4051-3995-3.
- ^ Нардини, Андреа; Ло Гулло, Мария А .; Саллео, Себастьяно (2011). «Повторное заполнение эмболизированных каналов ксилемы: это вопрос разгрузки флоэмы?». Растениеводство. 180 (4): 604–611. Дои:10.1016 / j.plantsci.2010.12.011. ISSN 0168-9452. PMID 21421408.
- ^ Диксон, Генри Х .; Джоли, Дж. (1894). «На восхождении сока». Анналы ботаники. 8: 468–470.
- ^ Диксон, Генри Х .; Джоли, Дж. (1895). «На восхождении сока». Философские труды Лондонского королевского общества, серия B. 186: 563–576. Дои:10.1098 / рстб.1895.0012.
- ^ Тайри, М. (1997). «Теория сплоченности-натяжения сокодородного восхождения: текущие споры». Журнал экспериментальной ботаники. 48 (10): 1753–1765. Дои:10.1093 / jxb / 48.10.1753. В архиве из оригинала от 20.02.2015.
- ^ Wang, Z .; Chang, C.-C .; Hong, S.J .; Sheng, Y.-J .; Цао, Х.-К. (2012). «Капиллярный подъем в микроканале произвольной формы и смачиваемости: петля гистерезиса». Langmuir. 28 (49): 16917–16926. Дои:10.1021 / la3036242. PMID 23171321.
- ^ Аскенасы, Э. (1895). "Ueber das Saftsteigen" [О восхождении сока]. Botanisches Centralblatt (на немецком). 62: 237–238.
- ^ Аскенасы, Э. (1895). "Ueber das Saftsteigen" [О восхождении сока]. Verhandlungen des Naturhistorisch-medizinischen Vereins zu Heidelberg (Труды Общества естествознания и медицины в Гейдельберге). 2-я серия (на немецком языке). 5: 325–345.
- ^ Диксон, H (1914). Транспирация и подъем сока у растений. Лондон, Англия, Великобритания: Macmillan and Co.
- ^ Диксон, H (1924). Поток транспирации. Лондон: University of London Press, Ltd. стр. 80.
- ^ Кэмпбелл, Нил (2002). Биология. Сан-Франциско, Калифорния: Pearson Education, Inc., стр.759. ISBN 978-0-8053-6624-2.
- ^ Циммерман, Ульрих (2002). «Каковы движущие силы для подъема воды в канале ксилемы?». Physiologia Plantarum. 114 (3): 327–335. Дои:10.1034 / j.1399-3054.2002.1140301.x. PMID 12060254.
- ^ Тайри, Мелвин Т. (1997). «Теория сплоченности-напряжения при восхождении сока: текущие споры». Журнал экспериментальной ботаники. 48 (10): 1753–1765. Дои:10.1093 / jxb / 48.10.1753.
- ^ Давление водного потенциала ксилемы в стебле растения можно определить с помощью бомбы Шоландера. bio.usyd.edu.au
- ^ Эндрю Дж. МакЭлрон, Брендан Чоут, Грег А. Гамбетта, Крейг Р. Бродерсен (2013). «Поглощение и перенос воды в сосудистых растениях». Проект знаний в области экологического образования.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Карлквист, S .; E.L. Шнайдер (2002). «Переход трахеида – сосудистый элемент у покрытосеменных имеет несколько независимых признаков: кладистические последствия». Американский журнал ботаники. 89 (2): 185–195. Дои:10.3732 / ajb.89.2.185. PMID 21669726.
- ^ Кронквист, А. (август 1988 г.). Эволюция и классификация цветковых растений. Нью-Йорк, Нью-Йорк: Пресса ботанического сада Нью-Йорка. ISBN 978-0-89327-332-3.
- ^ а б c d е ж г час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф Сперри, Дж. С. (2003). «Эволюция водного транспорта и структуры ксилемы». Международный журнал наук о растениях. 164 (3): S115 – S127. Дои:10.1086/368398. JSTOR 3691719. S2CID 15314720.
- ^ Эдвардс, Д .; Дэвис, К.Л .; Акс, Л. (1992). "Сосудистая проводящая нить у раннего наземного растения Cooksonia". Природа. 357 (6380): 683–685. Bibcode:1992Натура.357..683E. Дои:10.1038 / 357683a0. S2CID 4264332.
- ^ Никлас, К. Дж .; Смоковитис В. (1983). "Доказательства проводящей нити в растениях раннего силурия (лландовериане): значение для эволюции наземных растений". Палеобиология. 9 (2): 126–137. Дои:10.1017 / S009483730000751X. JSTOR 2400461. S2CID 35550235.
- ^ а б c d е ж г час Никлас, К. Дж. (1985). «Эволюция диаметра трахеиды у ранних сосудистых растений и ее влияние на гидравлическую проводимость первичной ксилемы». Эволюция. 39 (5): 1110–1122. Дои:10.2307/2408738. JSTOR 2408738. PMID 28561493.
- ^ Никлас, К .; Пратт, Л. (1980). «Доказательства наличия лигнин-подобных компонентов в окаменелостях растений раннего силурия (лландовериан)». Наука. 209 (4454): 396–397. Bibcode:1980Sci ... 209..396N. Дои:10.1126 / science.209.4454.396. PMID 17747811. S2CID 46073056.
- ^ Qiu, Y.L .; Li, L .; Ван, Б .; Chen, Z .; Knoop, V .; Groth-malonek, M .; Домбровская, О .; Lee, J .; Kent, L .; Rest, J .; и другие. (2006). «Самые глубокие расхождения в наземных растениях, вытекающие из филогеномных данных». Труды Национальной академии наук. 103 (42): 15511–6. Bibcode:2006PNAS..10315511Q. Дои:10.1073 / pnas.0603335103. ЧВК 1622854. PMID 17030812.
- ^ Стюарт, W.N .; Ротвелл, Г. (1993). Палеобиология и эволюция растений. Издательство Кембриджского университета.
- ^ Кораткар, Санджай (24.02.2016). «Кавитация и эмболия сосудистых растений (со схемой)». Обсуждение биологии.
- ^ Дэниел М. Джонсон, Кэтрин А. Маккалох, Дэвид Р. Вудрафф, Фредерик К. Майнзерк (июнь 2012 г.). «Гидравлический запас прочности и устранение эмболии в стеблях и листьях: почему хвойные деревья и покрытосеменные так отличаются?» (PDF). Лесная служба США.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ а б c d Фостер, A.S .; Гиффорд, Э.М. (1974). Сравнительная морфология сосудистых растений (2-е изд.). Сан-Франциско: W.H. Фримен. стр.55–56. ISBN 978-0-7167-0712-7.
- ^ Taylor, T.N .; Taylor, E.L .; Крингс, М. (2009). Палеоботаника, биология и эволюция ископаемых растений (2-е изд.). Амстердам; Бостон: Academic Press. стр. 207ff., 212ff. ISBN 978-0-12-373972-8.
- ^ Уайт, А. Тоби; Казлев, М. Алан. «Глоссарий». palaeos.com. Архивировано из оригинал 20 декабря 2010 г.
- ^ Увидеть:
- Чезальпино, Андреа (1583). De Plantis libri XVI [О растениях в 16 книгах.] (на латыни). Флоренция, Италия: Джорджо Марескотти. п.4. С п. 4: "An quædam sicca secundum naturam humorem trahunt? Ut lintea, spongiæ, pulveres:…" (Или [как] сухие вещи притягивают [т. Е. Впитывают] в соответствии с природой жидкости? [Например, белье, губки, порошки:…)
- Беллорини, Кристина (2016). Мир растений в Тоскане эпохи Возрождения: медицина и ботаника. Абингдон-он-Темз, Англия: Рутледж. п. 72. ISBN 9781317011491.
- Крамер, Пол Дж .; Бойер, Джон С. (1995). Водные отношения растений и почв. Лондон, Англия: Elsevier Science. п. 2. ISBN 9780080924113.
- ^ Увидеть:
- Мальпиги, Марчелло (1675). Anatome Plantarum… (на латыни). Лондон, Англия, Великобритания: Лондонское королевское общество. п. 8.
- Янсен, Стивен; Шенк, Х. Йохен (2015). «О всплытии сока при наличии пузырей». Американский журнал ботаники. 102 (10): 1561–1563. Дои:10.3732 / ajb.1500305. PMID 26400778.
- Лэзенби, Элизабет Мэри (1995) "The Historia Plantarum Generalis Джона Рэя: Книга I - перевод и комментарии », докторская диссертация, Университет Ньюкасл-апон-Тайн, Англия, Великобритания, том 1, стр. 160. Доступно по адресу: Университет Ньюкасл-апон-Тайн, Великобритания.
- ^ Рос, Неемия (1682 г.). Анатомия растений…. Лондон, Англия: У. Роулинз. С. 124–125. Из стр. 124–125: «Большую часть года он [то есть сок] поднимается в Барк [т.е. кора], сбн. во внутреннем Прибыль рядом с Дерево, а в весна, в или через Дерево он сам, и только там ".
- ^ Увидеть:
- (Grew, 1682), стр. 126. Грю признал пределы капиллярного действия (со стр. 126): «… маленький Стекло-трубы [то есть капиллярные трубки], погруженные в воду, будут давать ей [т.е. воде] подъем на несколько дюймов; но есть определенный период, согласно сверлить из Труба, выше которого он не поднимется ». Грю предложил следующий механизм подъема сока в растениях (из стр. 126):« Но Мочевого пузыря [то есть паренхимные клетки] DP, которые его окружают [то есть столб трахеид], набухшие и набухшие с Сок, настоящим дави на него; и поэтому не только немного сократит отверстие, но и перелит или Часть от их Сок в нем: обоими способами Сок будет вынужден подняться выше ".
- Арбер, Агнес (1913). "Неемия вырос в 1641–1712 гг.". В Оливер, Фрэнсис Уолл (ред.). Создатели британской ботаники: сборник биографий живых ботаников. Кембридж, Англия: Издательство Кембриджского университета. п. 58.
- ^ Хейлз, Стивен (1727). Vegetable Staticks: Или отчет о некоторых статических экспериментах с соком в овощах:…. Лондон, Англия: W. & J. Innys и T. Woodward. п.100. ISBN 9780356030128.
- ^ Увидеть:
- Страсбургер, Эдуард (1891). Histologische Beiträge [Гистологический вклад] (на немецком). Vol. 3: Ueber den Bau und die Verrichtungen der Leitungsbahnen in den Pflanzen [О строении и функции сосудистых пучков растений]. Йена, Германия: Густав Фишер. стр. 607–625: Aufsteigen giftiger Flüssigkeiten bis zu bedeutender Höhe in der Pflanze [Подъем ядовитых жидкостей на значительные высоты у растений], стр. 645–671: Die Leitungsfähigkeit getödteter Pflanzentheile [Способность погибших частей растений проводить [воду]].
- (Jansen & Schenck, 2015), стр. 1561.
Общие ссылки
- К. Вэй; Э. Стейдл; М. Т. Тайри; П. М. Линтильхак (май 2001 г.). «Основы прямого измерения давления ксилемы». Растение, клетка и окружающая среда. 24 (5): 549–555. Дои:10.1046 / j.1365-3040.2001.00697.x. S2CID 5039439. является основным источником, использованным в параграфе о недавнем исследовании.
- Н. Мишель Холбрук; Майкл Дж. Бернс; Кристофер Б. Филд (ноябрь 1995 г.). «Отрицательное давление ксилемы в растениях: испытание техники уравновешивания давления». Наука. 270 (5239): 1193–4. Bibcode:1995Научный ... 270.1193H. Дои:10.1126 / science.270.5239.1193. S2CID 97217181. это первое опубликованное независимое испытание, показывающее, что бомба Шоландера действительно измеряет напряжение в ксилеме.
- Pockman, W.T .; J.S. Сперри; J.W. О'Лири (декабрь 1995 г.). «Устойчивое и значительное отрицательное давление воды в ксилеме». Природа. 378 (6558): 715–6. Bibcode:1995Натура 378..715П. Дои:10.1038 / 378715a0. S2CID 31357329. это второй опубликованный независимый тест, показывающий, что бомба Шоландера действительно измеряет напряжение в ксилеме.
- Кэмпбелл, Нил А .; Джейн Б. Рис (2002). Биология (6-е изд.). Бенджамин Каммингс. ISBN 978-0-8053-6624-2.
- Кенрик, Пол; Крейн, Питер Р. (1997). Происхождение и ранняя диверсификация наземных растений: кладистическое исследование. Вашингтон, округ Колумбия: Пресса Смитсоновского института. ISBN 978-1-56098-730-7.
- Мухаммад, А. Ф .; Р. Саттлер (1982). "Конструкция сосуда Gnetum и происхождение покрытосеменных растений ". Американский журнал ботаники. 69 (6): 1004–21. Дои:10.2307/2442898. JSTOR 2442898.
- Мелвин Т. Тайри; Мартин Х. Циммерманн (2003). Структура ксилемы и подъем сока (2-е изд.). Springer. ISBN 978-3-540-43354-5. недавнее обновление классической книги покойного Мартина Циммерманна о ксилемном транспорте
внешние ссылки
 СМИ, связанные с Ксилема в Wikimedia Commons
СМИ, связанные с Ксилема в Wikimedia Commons
| Животные | |
|---|---|
| Растения | |
| |