Цитоскелет - Cytoskeleton
| Клеточная биология | |
|---|---|
| В животная клетка | |
 Компоненты типичной животной клетки:
|

В цитоскелет сложная, динамичная сеть взаимосвязанных белковые нити присутствует в цитоплазма из всех клетки, в том числе бактерии и археи.[1] Он простирается от ядро клетки к клеточная мембрана и состоит из аналогичных белков в различных организмах. В эукариоты, он состоит из трех основных компонентов: микрофиламенты, промежуточные нити и микротрубочки, и все они способны к быстрому росту или разборке в зависимости от требований клетки.[2]
Цитоскелет может выполнять множество функций. Его основная функция состоит в том, чтобы придать клетке ее форму и механическое сопротивление деформации, а также за счет связи с внеклеточными соединительная ткань и другие клетки стабилизируют целые ткани.[3][4] Цитоскелет также может сокращаться, тем самым деформируя клетку и среду клетки и позволяя клетки мигрировать.[5] Более того, он задействован во многих клеточная сигнализация путей и в поглощении внеклеточного материала (эндоцитоз ),[6] сегрегация хромосомы в течение клеточное деление,[3] то цитокинез стадия деления клеток,[7] как строительные леса для организации содержимого ячейки в пространстве[5] И в внутриклеточный транспорт (например, движение пузырьки и органеллы внутри клетки)[3] и может быть шаблоном для построения клеточная стенка.[3] Кроме того, он может образовывать специализированные структуры, такие как жгутики, реснички, ламеллиподии и подосомы. Структура, функции и динамическое поведение цитоскелета могут сильно отличаться в зависимости от организма и типа клеток.[3][7] Даже внутри одной клетки цитоскелет может изменяться за счет ассоциации с другими белками и предыдущей историей сети.[5]
Крупномасштабный пример действия цитоскелета: сокращение мышц. Это осуществляется группами высокоспециализированных ячеек, работающих вместе. Основным компонентом цитоскелета, который помогает показать истинную функцию сокращения мышц, является микрофиламент. Микрофиламенты состоят из наиболее распространенного клеточного белка, известного как актин.[8] Во время сокращения мышца внутри каждой мышечной клетки, миозин молекулярные моторы коллективно действуют на параллельные актин нити. Сокращение мышц начинается от нервных импульсов, которые затем вызывают высвобождение повышенного количества кальция из саркоплазматический ретикулум. Повышение содержания кальция в цитозоле позволяет мышцам начать сокращение с помощью двух белков: тропомиозин и тропонин.[8] Тропомиозин подавляет взаимодействие между актином и миозином, в то время как тропонин ощущает увеличение кальция и снимает подавление.[9] Это действие сокращает мышечную клетку и, благодаря синхронному процессу во многих мышечных клетках, всю мышцу.
История
В 1903 г. Кольцов Николай Константинович предположил, что форма клеток определяется сетью канальцев, которую он назвал цитоскелетом. Концепция белковой мозаики, которая динамически координирует биохимию цитоплазмы, была предложена Рудольфом Петерсом в 1929 году.[10] а срок (цитоскелет, на французском языке) был впервые представлен французским эмбриологом Пол Винтреберт в 1931 г.[11]
Когда цитоскелет был впервые представлен, считалось, что это неинтересное гелеобразное вещество, которое помогает органеллам оставаться на месте.[12] Было проведено много исследований, чтобы попытаться понять назначение цитоскелета и его компонентов. С помощью Стюарта Хамероффа и Роджера Пенроуза было обнаружено, что микротрубочки вибрируют внутри нейроны в головном мозге, предполагая, что мозговые волны возникают из-за более глубоких колебаний микротрубочек.[13] Это открытие продемонстрировало, что цитоскелет - это не просто гелеобразное вещество, и что у него действительно есть цель.[оспаривается ]
Первоначально считалось, что цитоскелет принадлежит исключительно эукариотам, но в 1992 году было обнаружено, что он присутствует и у прокариот. Это открытие произошло после осознания того, что бактерии обладают белками, гомологичными тубулину и актину; основные компоненты цитоскелета эукариот.[14]
Эукариотический цитоскелет
Эукариотический клетки содержат три основных типа филаментов цитоскелета: микрофиламенты, микротрубочки, и промежуточные нити. В нейроны промежуточные волокна известны как нейрофиламенты.[15] Каждый тип состоит из полимеризация особого типа субъединица белка и имеет свою характерную форму и внутриклеточный распространение. Микрофиламенты бывают полимеры белка актин и имеют диаметр 7 нм. Микротрубочки состоят из тубулин и имеют диаметр 25 нм. Промежуточные филаменты состоят из различных белков, в зависимости от типа клетки, в которой они находятся; они обычно имеют диаметр 8-12 нм.[1] Цитоскелет придает клетке структуру и форму, а исключая макромолекулы из некоторых цитозоль, это увеличивает уровень макромолекулярное скопление в этом отсеке.[16] Элементы цитоскелета широко и тесно взаимодействуют с клеточными мембранами.[17]
Исследования в нейродегенеративные расстройства такие как болезнь Паркинсона, Болезнь Альцгеймера, болезнь Хантингтона, и боковой амиотрофический склероз (БАС) указывают на поражение цитоскелета при этих заболеваниях.[18] Болезнь Паркинсона характеризуется деградацией нейронов, что приводит к тремору, ригидности и другим немоторным симптомам. Исследования показали, что сборка и стабильность микротрубочек в цитоскелете нарушены, что приводит к деградации нейронов со временем.[19] При болезни Альцгеймера тау-белки которые стабилизируют нарушение функции микротрубочек по мере прогрессирования заболевания, вызывающего патологию цитоскелета.[20] Избыток глутамина в белке Хантингтона, участвующий в связывании везикул с цитоскелетом, также считается фактором развития болезни Хантингтона.[21] Боковой амиотрофический склероз приводит к потере подвижности, вызванной деградацией мотонейронов, а также к дефектам цитоскелета.[22]
Дополнительные белки, включая моторные белки регулируют и связывают филаменты с другими клеточными соединениями и друг с другом и необходимы для контролируемой сборки филаментов цитоскелета в определенных местах.[23]
Ряд низкомолекулярных цитоскелетные препараты были обнаружены, которые взаимодействуют с актином и микротрубочками. Эти соединения оказались полезными при изучении цитоскелета, а некоторые из них имеют клиническое применение.
Микрофиламенты
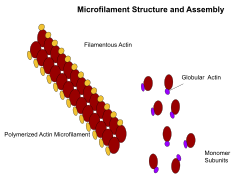
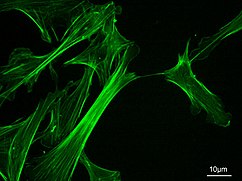
Микрофиламенты, также известные как актиновые филаменты, состоят из линейных полимеров G-актин белки, и генерируют силу, когда растущий (положительный) конец филамента толкает барьер, такой как клеточная мембрана. Они также служат дорожками для движения миозин молекулы, которые прикрепляются к микрофиламенту и «ходят» по ним. Как правило, основным компонентом микрофиламентов является актин. Мономер G-актина объединяется с образованием полимера, который продолжает формировать микрофиламент (актиновую нить). Затем эти субъединицы собираются в две цепи, которые переплетаются в так называемые F-актин цепи.[24] Миозин, движущийся по филаментам F-актина, генерирует сократительные силы в так называемых актомиозиновых волокнах, как в мышцах, так и в большинстве немышечных типов клеток.[25] Структуры актина контролируются Семья Ро малых GTP-связывающих белков, таких как сам Rho для сократительных актомиозиновых филаментов («стрессовых волокон»), Rac для ламеллиподий и Cdc42 для филоподий.
Функции включают:
- Сокращение мышц
- Движение клеток
- Внутриклеточный транспорт / торговля
- Обслуживание эукариотический форма ячейки
- Цитокинез
- Цитоплазматический поток[24]
Промежуточные нити
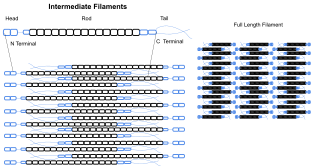
Промежуточные филаменты являются частью цитоскелета многих эукариотический клетки. Эти волокна диаметром в среднем 10 нанометров более стабильны (прочно связаны), чем микрофиламенты, и гетерогенные составляющие цитоскелета. подобно актин филаментов, они функционируют в поддержании формы ячеек за счет несущего напряжения (микротрубочки, напротив, сопротивляются сжатию, но также могут выдерживать растяжение во время митоз и во время позиционирования центросомы). Промежуточные волокна организуют внутреннюю трехмерную структуру ячейки, закрепляя органеллы и выступая в качестве структурных компонентов ядерная пластинка. Они также участвуют в некоторых соединениях клетка-клетка и клетка-матрица. Ядерная пластинка существуют во всех животных и во всех тканях. Некоторым животным нравится плодовая муха не имеют цитоплазматических промежуточных филаментов. У тех животных, которые экспрессируют цитоплазматические промежуточные филаменты, они тканеспецифичны.[4] Кератиновые промежуточные нити в эпителиальный клетки обеспечивают защиту от различных механических нагрузок, которые может выдержать кожа. Они также обеспечивают защиту органов от метаболических, окислительных и химических стрессов. Укрепление эпителиальных клеток этими промежуточными филаментами может предотвратить начало апоптоз или гибель клеток за счет снижения вероятности стресса.[26]
Промежуточные филаменты чаще всего известны как система поддержки или «каркас» для клетки и ядра, а также играют роль в некоторых функциях клетки. В сочетании с белками и десмосомы промежуточные филаменты образуют связи клетка-клетка и закрепляют соединения клетка-матрица, которые используются для обмена сообщениями между клетками, а также для жизненно важных функций клетки. Эти соединения позволяют клетке общаться через десмосомы нескольких клеток, чтобы регулировать структуры ткани на основе сигналов из клеточной среды. Было показано, что мутации в белках IF вызывают серьезные медицинские проблемы, такие как преждевременное старение, мутации десмина, поражающие органы, Болезнь Александра, и мышечная дистрофия.[27]
Различные промежуточные филаменты:
- сделано из виментины. Промежуточные филаменты виментина обычно присутствуют в мезенхимальных клетках.
- сделано из кератин. Кератин в основном присутствует в эпителиальных клетках.
- нейрофиламенты нервных клеток.
- сделано из ламинат, обеспечивая структурную поддержку ядерной оболочки.
- сделано из десмин, играют важную роль в структурной и механической поддержке мышечных клеток.[28]
Микротрубочки
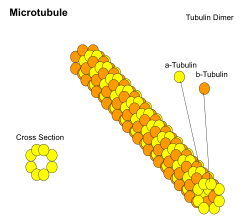
Микротрубочки представляют собой полые цилиндры диаметром около 23 нм (диаметр просвета около 15 нм), чаще всего состоящие из 13 протофиламенты которые, в свою очередь, являются полимерами альфа и бета тубулин. У них очень динамичное поведение, обязательное GTP для полимеризации. Обычно они организуются центросома.
В девяти тройках (звездчатых) они образуют центриоли, а в девяти дублетах, ориентированных около двух дополнительных микротрубочек (колесообразных), они образуют реснички и жгутики. Последнее образование обычно называют расположением «9 + 2», в котором каждый дублет соединен с другим белком. динеин. Поскольку и жгутики, и реснички являются структурными компонентами клетки и поддерживаются микротрубочками, их можно рассматривать как часть цитоскелета. Реснички бывают двух типов: подвижные и неподвижные. Реснички короткие, их больше, чем жгутиков. Подвижные реснички имеют ритмичное колебательное или биение движения по сравнению с неподвижными ресничками, которые получают сенсорную информацию для клетки; обработка сигналов от других ячеек или окружающих их жидкостей. Кроме того, микротрубочки контролируют биение (движение) ресничек и жгутиков.[29] Кроме того, динеиновые плечи, прикрепленные к микротрубочкам, действуют как молекулярные двигатели. Движение ресничек и жгутиков создается за счет скольжения микротрубочек друг за другом, для чего требуется АТФ.[29]Они играют ключевые роли в:
- внутриклеточный транспорт (связанный с динеинами и кинезины они транспортируют органеллы любить митохондрии или пузырьки ).
- то аксонема из реснички и жгутики.
 Схема поперечного сечения реснички, показывающая расположение микротрубочек «9 + 2»
Схема поперечного сечения реснички, показывающая расположение микротрубочек «9 + 2» - то митотическое веретено.
- синтез клеточной стенки у растений.

В дополнение к ролям, описанным выше, Стюарт Хамерофф и Роджер Пенроуз предположили, что микротрубочки функционируют в сознании.[30]
Сравнение
| Цитоскелет тип[31] | Диаметр (нм )[32] | Структура | Примеры субъединиц[31] |
|---|---|---|---|
| Микрофиламенты | 6 | Двойная спираль | Актин |
| Средний нити | 10 | Два антипараллельных спирали / димеры, образующие тетрамеры |
|
| Микротрубочки | 23 | Протофиламенты, в свою очередь состоящий из субъединиц тубулина в комплексе с Stathmin[33] | α- и β-тубулин |
Септины
Септины - это группа высококонсервативных GTP связывающие белки, обнаруженные в эукариоты. Различные формы септинов белковые комплексы друг с другом. Они могут собираться в нити и кольца. Следовательно, септины можно рассматривать как часть цитоскелета.[34] Функция септинов в клетках заключается в том, что они служат локализованным местом прикрепления для других белки, и предотвращение распространение определенных молекул из одного отсека клетки в другой.[34] В дрожжевых клетках они создают каркас, чтобы обеспечить структурную поддержку во время деления клеток и разделить части клетки. Недавние исследования человеческих клеток показывают, что септины создают клетки вокруг бактериальных патогенов, обездвиживая вредные микробы и предотвращая их вторжение в другие клетки.[35]
Спектрин
Спектрин - это цитоскелет белок которая выстилает внутриклеточную сторону плазматическая мембрана в эукариотических клетках. Спектрин образует пятиугольные или шестиугольные формы, образуя строительные леса и играет важную роль в поддержании плазматическая мембрана целостность и структура цитоскелета.[36]
Цитоскелет дрожжей
В зародыше дрожжи (важно модельный организм ), актин образует корковые пятна, актиновые кабели, цитокинетическое кольцо и колпачок. Корковые пятна представляют собой дискретные актиновые тела на мембране, жизненно важные для эндоцитоз, особенно переработка глюкансинтазы, которая важна для клеточная стенка синтез. Кабели Actin представляют собой пучки актиновые нити и занимаются транспортировкой пузырьки к крышке (которая содержит ряд различных белков для поляризации роста клеток) и в расположении митохондрий. В цитокинетический кольцо образует и сужается вокруг участка деление клеток.[37]
Прокариотический цитоскелет
До работы Jones et al., 2001 считалось, что клеточная стенка является решающим фактором для многих форм бактериальных клеток, включая палочки и спирали. При изучении было обнаружено, что многие деформированные бактерии имеют мутации, связанные с развитием клеточная оболочка.[38] Когда-то считалось, что цитоскелет является особенностью только эукариотический клетки, но гомологи ко всем основным белкам цитоскелета эукариот, которые были обнаружены в прокариоты.[39] Гарольд Эриксон отмечает, что до 1992 года считалось, что только эукариоты имеют компоненты цитоскелета. Однако исследования в начале 90-х годов показали, что у бактерий и архей есть гомологи актина и тубулина, и что они лежат в основе микротрубочек и микрофиламентов эукариот.[40] Хотя эволюционные отношения настолько далеки, что не очевидны только из сравнения последовательностей белков, сходство их трехмерных структуры и сходные функции по поддержанию формы и полярности клеток обеспечивают убедительное доказательство того, что эукариотические и прокариотические цитоскелеты действительно гомологичны.[41] Три лаборатории независимо друг от друга обнаружили, что белок FtsZ, уже известный как ключевой игрок в бактериальном цитокинезе, имеет «сигнатурную последовательность тубулина», присутствующую во всех α-, β- и γ-тубулинах.[40] Однако некоторые структуры бактериального цитоскелета, возможно, еще не идентифицированы.[25][42]
FtsZ
FtsZ был первым идентифицированным белком прокариотического цитоскелета. Как и тубулин, FtsZ образует филаменты в присутствии гуанозинтрифосфат (GTP), но эти нити не группируются в канальцы. В течение деление клеток, FtsZ - это первый белок, который переместится в сайт деления, и он важен для набора других белков, которые синтезируют новые клеточная стенка между делящимися клетками.
MreB и ParM
Прокариотические актин-подобные белки, такие как MreB, участвуют в поддержании формы клеток. Все несферические бактерии имеют гены кодирующие актин-подобные белки, и эти белки образуют спиральную сеть под клеточной мембраной, которая направляет белки, участвующие в клеточной стенке биосинтез.[43]
Немного плазмиды кодируют отдельную систему, которая включает актин-подобный белок ParM. Нити ParM экспонируются динамическая нестабильность, и может разделять плазмидную ДНК на делящиеся дочерние клетки по механизму аналогичный к тому, что используется микротрубочками во время эукариотического митоз.[25][44]
Crescentin
Бактерия Caulobacter crescentus содержит третий белок, полумесяц, относящийся к промежуточным филаментам эукариотических клеток. Кресцентин также участвует в поддержании формы клеток, например спиральной и вибриоид формы бактерий, но механизм, с помощью которого это происходит, в настоящее время неясен.[45] Кроме того, кривизна может быть описана смещением серповидных нитей после нарушения синтеза пептидогликана.[46]
Общие черты и различия между прокариотами и эукариотами
По определению, цитоскелет состоит из белков, которые могут образовывать продольные массивы (волокна) у всех организмов. Эти белки, образующие волокна, подразделяются на 4 класса. Тубулин -любить, актин -любить, Цитоскелетные АТФазы Walker A (WACA-белки) и промежуточные нити.[7][25]
Тубулиноподобные белки тубулин у эукариот и FtsZ, TubZ, RepX у прокариот. Актин-подобные белки актин у эукариот и MreB, FtsA у прокариот. Примером WACA-белков, которые в основном обнаруживаются у прокариот, является Разум. Примерами промежуточных волокон, которые почти исключительно обнаруживаются у животных (например, у эукариот), являются ламины, кератины, виментин, нейрофиламенты, и десмин.[7]
Хотя тубулиноподобные белки разделяют некоторые аминокислотная последовательность подобие, их эквивалентность в белковая складка и сходство в GTP место связывания более поразительно. То же самое верно и для актин-подобных белков, их структуры и АТФ связывающий домен.[7][25]
Белки цитоскелета обычно коррелируют с формой клеток, сегрегацией ДНК и делением клеток у прокариот и эукариот. Какие белки выполняют какую задачу - разные вещи. Например, сегрегация ДНК у всех эукариот происходит за счет использования тубулина, но у прокариот могут использоваться белки WACA, актиноподобные или тубулиноподобные белки. У эукариот деление клеток опосредуется актином, но у прокариот обычно тубулиноподобными (часто FtsZ-кольцом) белками и иногда (Crenarchaeota ) ESCRT-III, который у эукариот все еще играет роль на последнем этапе деления.[7]
Цитоплазматический поток
Цитоплазматический поток, также известный как циклоз, представляет собой активное движение содержимого клетки по компонентам цитоскелета. Хотя в основном это наблюдается у растений, все типы клеток используют этот процесс для транспортировки отходов, питательных веществ и органелл в другие части клетки.[47] Клетки растений и водорослей обычно больше, чем многие другие клетки; поэтому цитоплазматический поток важен в этих типах клеток. Это связано с тем, что дополнительный объем клетки требует потока цитоплазмы, чтобы перемещать органеллы по всей клетке.[48] Органеллы движутся микрофиламенты в цитоскелете, управляемом миозин двигатели заедают и толкают актин пучки нитей.[47]
Смотрите также
использованная литература
- ^ а б Хардин Дж, Бертони Дж, Кляйнсмит Л. Дж. (2015). Мир клетки Беккера (8-е изд.). Нью-Йорк: Пирсон. С. 422–446. ISBN 978013399939-6.
- ^ МакКинли, Майкл; Дин О'Лафлин, Валери; Пеннефатер-О'Брайен, Элизабет; Харрис, Рональд (2015). Человеческая анатомия (4-е изд.). Нью-Йорк: Образование Макгроу Хилл. п. 29. ISBN 978-0-07-352573-0.
- ^ а б c d е Альбертс Б. и др. (2008). Молекулярная биология клетки (5-е изд.). Нью-Йорк: Наука Гарланд. ISBN 978-0-8153-4105-5.
- ^ а б Herrmann H, Bär H, Kreplak L, Стрелков С.В., Aebi U (июль 2007 г.). «Промежуточные филаменты: от клеточной архитектуры до наномеханики». Обзоры природы. Молекулярная клеточная биология. 8 (7): 562–73. Дои:10.1038 / nrm2197. PMID 17551517. S2CID 27115011.
- ^ а б c Флетчер Д.А., Маллинз Р.Д. (январь 2010 г.). «Клеточная механика и цитоскелет». Природа. 463 (7280): 485–92. Bibcode:2010Натура.463..485F. Дои:10.1038 / природа08908. ЧВК 2851742. PMID 20110992.
- ^ Гели М.И., Рицман Х. (апрель 1998 г.). «Эндоцитарная интернализация в клетках дрожжей и животных: похожие и разные». Журнал клеточной науки. 111 (Pt 8) (8): 1031–7. PMID 9512499.
- ^ а б c d е ж Уикстед Б., Чайка К. (август 2011 г.). «Эволюция цитоскелета». Журнал клеточной биологии. 194 (4): 513–25. Дои:10.1083 / jcb.201102065. ЧВК 3160578. PMID 21859859.
- ^ а б Купер, Джеффри М. (2000). «Актин, миозин и движение клеток». Клетка: молекулярный подход. 2-е издание. В архиве из оригинала от 28.04.2018.
- ^ Берг Дж. М., Тимочко Дж. Л., Страйер Л. (2002). «Миозины движутся вдоль актиновых волокон». Биохимия. 5-е издание. В архиве из оригинала от 2018-05-02.
- ^ Петерс Р.А. "The Harben Lectures, 1929. Перепечатано в: Peters, R.A. (1963) Биохимические поражения и летальный синтез, стр. 216. Pergamon Press, Oxford". Цитировать журнал требует
| журнал =(Помогите) - ^ Frixione E (июнь 2000 г.). «Повторяющиеся взгляды на структуру и функцию цитоскелета: 300-летний эпос». Подвижность клеток и цитоскелет. 46 (2): 73–94. Дои:10.1002 / 1097-0169 (200006) 46: 2 <73 :: AID-CM1> 3.0.CO; 2-0. PMID 10891854. S2CID 16728876.
- ^ Хардин Дж (03.12.2015). Мир клетки Беккера (9-е изд.). Пирсон. п. 351. ISBN 978-0-321-93492-5.
- ^ Эльзевир. «Открытие квантовых колебаний в« микротрубочках »внутри нейронов мозга подтверждает противоречивую теорию сознания 20-летней давности». www.elsevier.com. В архиве из оригинала от 07.11.2016. Получено 2017-11-20.
- ^ Уикстед Б., Чайка К. (август 2011 г.). «Эволюция цитоскелета». Журнал клеточной биологии. 194 (4): 513–25. Дои:10.1083 / jcb.201102065. ЧВК 3160578. PMID 21859859.
- ^ Таран, АС; Шувалова, ЛД; Лагаркова, М.А. Алиева И.Б. (22 июня 2020 г.). "Болезнь Хантингтона - взгляд на взаимодействие белка HTT, микротрубочек и актиновых компонентов цитоскелета". Клетки. 9 (6). Дои:10.3390 / ячейки9061514. PMID 32580314.
- ^ Минтон А.П. (октябрь 1992 г.). «Конфайнмент как детерминант макромолекулярной структуры и реакционной способности». Биофизический журнал. 63 (4): 1090–100. Bibcode:1992BpJ .... 63.1090M. Дои:10.1016 / S0006-3495 (92) 81663-6. ЧВК 1262248. PMID 1420928. В архиве из оригинала от 15.05.2013.
- ^ Доэрти Г.Дж., МакМахон ХТ (2008). «Посредничество, модуляция и последствия взаимодействий мембрана-цитоскелет». Ежегодный обзор биофизики. 37: 65–95. Дои:10.1146 / annurev.biophys.37.032807.125912. PMID 18573073. S2CID 17352662.
- ^ Пелуччи, Сильвия; Стрингхи, Рамона; Марчелло, Елена (2020). «Дендритные шипы при болезни Альцгеймера: как актиновый цитоскелет способствует синаптической недостаточности». Международный журнал молекулярных наук. 21 (3): 908. Дои:10.3390 / ijms21030908. ISSN 1422-0067. ЧВК 7036943. PMID 32019166.
- ^ Пеллегрини Л., Ветцель А., Гранно С., Хитон Г., Харви К. (февраль 2017 г.). «Обратно к канальцу: динамика микротрубочек при болезни Паркинсона». Клеточные и молекулярные науки о жизни. 74 (3): 409–434. Дои:10.1007 / s00018-016-2351-6. ЧВК 5241350. PMID 27600680.
- ^ Бамбург-младший, Блум Г.С. (август 2009 г.). «Патологии цитоскелета при болезни Альцгеймера». Подвижность клеток и цитоскелет. 66 (8): 635–49. Дои:10.1002 / см. 20388. ЧВК 2754410. PMID 19479823.
- ^ Кавистон Дж. П., Хольцбаур Е. Л. (апрель 2009 г.). «Белок Хантингтин является важным интегратором внутриклеточного везикулярного обмена». Тенденции в клеточной биологии. 19 (4): 147–55. Дои:10.1016 / j.tcb.2009.01.005. ЧВК 2930405. PMID 19269181.
- ^ Жюльен Дж. П., Миллекэмпс С., Криз Дж. (2005). «Дефекты цитоскелета при боковом амиотрофическом склерозе (заболевание двигательных нейронов)». Симпозиум Фонда Новартис. 264: 183–92, обсуждение 192–6, 227–30. PMID 15773754.
- ^ Альбертс, Брюс (2015). Молекулярная биология клетки. Наука о гирляндах. п. 889. ISBN 978-0-8153-4464-3.
- ^ а б Купер, Джеффри М. (2000). «Структура и организация актиновых волокон». Клетка: молекулярный подход. 2-е издание. В архиве из оригинала от 2018-05-02.
- ^ а б c d е Gunning PW, Ghoshdastider U, Whitaker S, Popp D, Robinson RC (июнь 2015 г.). «Эволюция композиционно и функционально различных актиновых филаментов». Журнал клеточной науки. 128 (11): 2009–19. Дои:10.1242 / jcs.165563. PMID 25788699.
- ^ Пан X, Хоббс Р.П., Куломб, Пенсильвания (февраль 2013 г.). «Растущее значение кератиновых промежуточных филаментов в нормальном и больном эпителии». Текущее мнение в области клеточной биологии. 25 (1): 47–56. Дои:10.1016 / j.ceb.2012.10.018. ЧВК 3578078. PMID 23270662.
- ^ Herrmann H, Bär H, Kreplak L, Стрелков С.В., Aebi U (июль 2007 г.). «Промежуточные филаменты: от клеточной архитектуры до наномеханики». Обзоры природы. Молекулярная клеточная биология. 8 (7): 562–73. Дои:10.1038 / nrm2197. PMID 17551517. S2CID 27115011.
- ^ Паулин Д., Ли Зи (ноябрь 2004 г.). «Десмин: основной белок промежуточных филаментов, необходимый для структурной целостности и функции мышц». Экспериментальные исследования клеток. 301 (1): 1–7. Дои:10.1016 / j.yexcr.2004.08.004. PMID 15501438.
- ^ а б Лодиш, Харви; Берк, Арнольд; Зипурский, С. Лоуренс; Мацудаира, Пол; Балтимор, Дэвид; Дарнелл, Джеймс (2 мая 2018 г.). «Реснички и жгутики: структура и движение». В архиве из оригинала 2 мая 2018 г.. Получено 2 мая 2018 - через www.ncbi.nlm.nih.gov. Цитировать журнал требует
| журнал =(Помогите) - ^ Хамерофф, С. и Пенроуз, Р. Обзоры по физике жизни, 2014, 11, 39-78
- ^ а б Если иное не указано в полях, ref:Бор WF (2003). Медицинская физиология: клеточный и молекулярный подход. Elsevier / Saunders. п. 1300. ISBN 978-1-4160-2328-9. Стр.25
- ^ Fuchs E, Cleveland DW (январь 1998 г.). «Структурный каркас из промежуточных волокон в здоровье и болезни». Наука. 279 (5350): 514–9. Bibcode:1998Sci ... 279..514F. Дои:10.1126 / science.279.5350.514. PMID 9438837.
- ^ Steinmetz MO (май 2007 г.).«Структура и термодинамика взаимодействия тубулина и статмина». Журнал структурной биологии. 158 (2): 137–47. Дои:10.1016 / j.jsb.2006.07.018. PMID 17029844.
- ^ а б Мостовой С., Коссарт П. (февраль 2012 г.). «Септины: четвертый компонент цитоскелета». Обзоры природы. Молекулярная клеточная биология. 13 (3): 183–94. Дои:10.1038 / nrm3284. PMID 22314400. S2CID 2418522.
- ^ Mascarelli A (декабрь 2011 г.). «Белки септина берут в плен бактерии: клеточная защита от микробных патогенов имеет терапевтический потенциал». Природа. Дои:10.1038 / природа.2011.9540. S2CID 85080734.
- ^ Ха Г.Ю., Гланц С.Б., Дже С., Морроу Дж. С., Ким Дж. Х. (декабрь 2001 г.). «Кальпаиновый протеолиз альфа II-спектрина в нормальном мозге взрослого человека». Письма о неврологии. 316 (1): 41–4. Дои:10.1016 / S0304-3940 (01) 02371-0. PMID 11720774. S2CID 53270680.
- ^ Прюин Д., Бретчер А. (февраль 2000 г.). «Поляризация роста клеток дрожжей». Журнал клеточной науки. 113 (Pt 4) (4): 571–85. PMID 10652251.
- ^ Jones, Laura J. F .; Карбаллидо-Лопес, Рут; Эррингтон, Джеффри (23 марта 2001 г.). «Контроль формы клеток в бактериях: спиральные актин-подобные нити в Bacillus subtilis». Ячейка. 104 (6): 913–922. Дои:10.1016 / S0092-8674 (01) 00287-2. PMID 11290328. S2CID 14207533.
- ^ Ши Ю.Л., Ротфилд Л. (сентябрь 2006 г.). «Бактериальный цитоскелет». Обзоры микробиологии и молекулярной биологии. 70 (3): 729–54. Дои:10.1128 / MMBR.00017-06. ЧВК 1594594. PMID 16959967.
- ^ а б Эриксон HP (февраль 2017 г.). «Открытие прокариотического цитоскелета: 25 лет». Молекулярная биология клетки. 28 (3): 357–358. Дои:10.1091 / mbc.E16-03-0183. ЧВК 5341718. PMID 28137947.
- ^ Мичи КА, Лёве Дж (2006). «Динамические нити цитоскелета бактерий» (PDF). Ежегодный обзор биохимии. 75: 467–92. Дои:10.1146 / annurev.biochem.75.103004.142452. PMID 16756499.
- ^ Бригель А., Диас Д.П., Ли З., Дженсен Р.Б., Франгакис А.С., Дженсен Г.Дж. (октябрь 2006 г.). «Множественные большие пучки филаментов наблюдаются у Caulobacter crescentus с помощью электронной криотомографии». Молекулярная микробиология. 62 (1): 5–14. Дои:10.1111 / j.1365-2958.2006.05355.x. PMID 16987173.
- ^ Попп Д., Нарита А., Маэда К., Фудзисава Т., Гошдастидер Ю., Иваса М., Маеда И., Робинсон Р.С. (май 2010 г.). «Структура, организация и динамика волокон в листах MreB». Журнал биологической химии. 285 (21): 15858–65. Дои:10.1074 / jbc.M109.095901. ЧВК 2871453. PMID 20223832.
- ^ Попп Д., Нарита А., Ли Л.Дж., Гошдастидер Ю., Сюэ Б., Сринивасан Р., Баласубраманиан М.К., Танака Т., Робинсон Р.К. (июнь 2012 г.). «Новая актиноподобная структура филаментов из Clostridium tetani». Журнал биологической химии. 287 (25): 21121–9. Дои:10.1074 / jbc.M112.341016. ЧВК 3375535. PMID 22514279.
- ^ Осмис Н., Кун Дж. Р., Якобс-Вагнер С. (декабрь 2003 г.). «Бактериальный цитоскелет: промежуточная филаментоподобная функция в форме клетки». Ячейка. 115 (6): 705–13. Дои:10.1016 / S0092-8674 (03) 00935-8. PMID 14675535. S2CID 14459851.
- ^ Эсу, Осигве (январь 2010 г.). «Динамика бактериального промежуточного филамента кресцентина in vitro и in vivo». PLOS ONE. 5 (1): e8855. Bibcode:2010PLoSO ... 5.8855E. Дои:10.1371 / journal.pone.0008855. ЧВК 2816638. PMID 20140233. Получено 12 сентября 2017.
- ^ а б Вудхаус Ф.Г., Гольдштейн Р. Э. (август 2013 г.). «Цитоплазматический поток в растительных клетках возникает естественным образом за счет самоорганизации микрофиламентов». Труды Национальной академии наук Соединенных Штатов Америки. 110 (35): 14132–7. arXiv:1308.6422. Bibcode:2013PNAS..11014132W. Дои:10.1073 / pnas.1302736110. ЧВК 3761564. PMID 23940314.
- ^ Гольдштейн Р. Е., ван де Меент Дж. В. (август 2015 г.). «Физическая перспектива цитоплазматического потока». Фокус интерфейса. 5 (4): 20150030. Дои:10.1098 / rsfs.2015.0030. ЧВК 4590424. PMID 26464789.
внешние ссылки
- Ежемесячные новости и блог Cytoskeleton
- MBInfo - Динамика цитоскелета
- Цитоскелет, подвижность клеток и двигатели - Виртуальная библиотека биохимии, молекулярной биологии и клеточной биологии
- База данных цитоскелета, клинические испытания, свежая литература, лабораторный регистр ...
- Анимация адгезии лейкоцитов (Анимация с некоторыми изображениями сборки и динамики актина и микротрубочек.)
- http://cellix.imba.oeaw.ac.at/ Цитоскелет и подвижность клеток, включая видео
- Обзорная статья в открытом доступе на возникающей сложности цитоскелета (появилась в Успехи в физике, 2013)