Кинезин - Kinesin

А кинезин белок, принадлежащий к классу моторные белки нашел в эукариотический клетки.
Кинезины движутся микротрубочка (MT) филаменты, и питаются за счет гидролиза аденозинтрифосфат (АТФ) (таким образом, кинезины АТФазы ), тип фермента. Активное движение кинезинов поддерживает несколько клеточных функций, включая митоз, мейоз и транспортировка сотовых грузов, например, в аксональный транспорт. Большинство кинезинов движется к положительному концу микротрубочек, что в большинстве клеток влечет за собой транспортировку грузов, таких как белковые и мембранные компоненты, от центра клетки к периферии.[1] Этот вид транспорта известен как антероградный транспорт. В отличие, динеины моторные белки, которые движутся к минус-концу микротрубочки в ретроградный транспорт.
Открытие
Кинезины были обнаружены в 1985 году на основании их подвижности в цитоплазме, выделенной из гигантского аксона кальмара.[2]
Оказалось, что они представляют собой двигатели антероградного внутриклеточного транспорта на основе МТ.[3] Член-основатель этого суперсемейства, кинезин-1, был выделен в виде гетеротетрамерный быстрый мотор транспорта аксональных органелл, состоящий из 2 идентичных моторных субъединиц (KHC) и 2 «легких цепей» (KLC), посредством аффинной очистки микротрубочек из экстрактов нервных клеток.[4] Впоследствии из экстрактов яиц / эмбрионов иглокожих был очищен другой гетеротримерный мотор на основе МТ, направленный на плюс-конец, названный кинезин-2, состоящий из 2 различных KHC-связанных моторных субъединиц и дополнительной субъединицы «KAP».[5] и наиболее известен своей ролью в транспортировке белковых комплексов (IFT-частиц) по аксонемам во время ресничка биогенез.[6] Молекулярно-генетический и геномный подходы привели к признанию того, что кинезины образуют разнообразное суперсемейство моторов, которые ответственны за множественные события внутриклеточной подвижности в эукариотических клетках.[7][8][9][10] Например, геномы млекопитающих кодируют более 40 белков кинезина,[11] организованы по крайней мере в 14 семейств, названных от кинезин-1 до кинезин-14.[12]
Структура
Общая структура
Члены суперсемейства кинезинов различаются по форме, но прототипный мотор кинезина-1 состоит из двух молекул тяжелой цепи кинезина (KHC), которые образуют димер белка (пара молекул), связывающая две легкие цепи (KLC), уникальные для разных грузов.
Тяжелая цепь кинезина-1 включает глобулярную голова (моторный домен) на аминоконцевом конце, соединенный коротким гибким шейным линкером с стебель - длинная центральная альфа-спираль спиральная катушка домен - который заканчивается карбокси-концом хвост домен, который связан с легкими цепями. Стебли двух KHC переплетаются, образуя спиральная катушка который направляет димеризацию двух KHC. В большинстве случаев транспортируемый груз связывается с легкими цепями кинезина в Мотив TPR последовательность KLC, но в некоторых случаях груз связывается с С-концевыми доменами тяжелых цепей.[13]
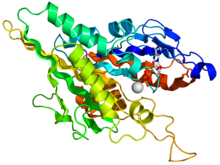
Кинезин моторный домен
| Кинезин моторный домен | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Идентификаторы | |||||||||
| Символ | Кинезин моторный домен | ||||||||
| Pfam | PF00225 | ||||||||
| ИнтерПро | IPR001752 | ||||||||
| УМНАЯ | SM00129 | ||||||||
| PROSITE | PS50067 | ||||||||
| SCOP2 | 1bg2 / Объем / СУПФАМ | ||||||||
| CDD | cd00106 | ||||||||
| |||||||||
Голова - это сигнатура кинезина, и ее аминокислотная последовательность хорошо сохраняется среди различных кинезинов. Каждая голова имеет два отдельных участок связывания: один для микротрубочек, а другой для АТФ. Связывание и гидролиз АТФ, а также ADP высвобождение изменяет конформацию связывающих микротрубочки доменов и ориентацию шейного линкера по отношению к голове; это приводит к движению кинезина. Несколько структурных элементов в голове, в том числе центральный бета-лист домен и домены Switch I и II, как предполагается, опосредуют взаимодействия между двумя сайтами связывания и шейным доменом. Кинезины структурно связаны с G белки, которые гидролизуют GTP вместо АТФ. Несколько структурных элементов являются общими для двух семейств, в частности, домена Switch I и Switch II.

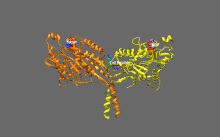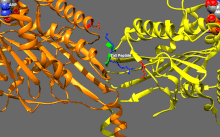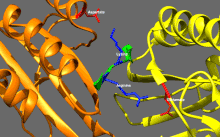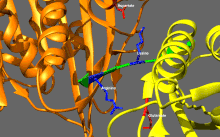
Базовая регуляция кинезина
Кинезины, как правило, обладают низкой базальной ферментативной активностью, которая становится значительной при активации микротрубочек.[16] Кроме того, многие члены суперсемейства кинезинов могут самоингибироваться за счет связывания хвостового домена с моторным доменом.[17] Такое самоторможение затем можно устранить с помощью дополнительных правил, таких как привязка к грузу или грузовым адаптерам.[18][19]
Грузовой транспорт
В клетке небольшие молекулы, такие как газы и глюкоза, размытый туда, где они нужны. В теле клетки синтезируются крупные молекулы, внутриклеточные компоненты, такие как пузырьки и органеллы, такие как митохондрии слишком большие (и цитозоль слишком многолюдно), чтобы добраться до места назначения. Моторные белки выполняют роль транспортировки больших грузов по клетке к нужному месту назначения. Кинезины - это моторные белки, которые транспортируют такой груз, двигаясь в одном направлении. микротрубочка отслеживает гидролиз одной молекулы аденозинтрифосфат (АТФ) на каждом этапе.[20] Считалось, что АТФ гидролиз приводил в действие каждый шаг, высвобождаемая энергия толкала голову вперед к следующему месту связывания.[21] Однако было высказано предположение, что головка диффундирует вперед, и сила связывания с микротрубочкой - это то, что тянет за собой груз.[22] Вдобавок вирусы, например ВИЧ, используют кинезины, чтобы позволить вирусным частицам перемещаться после сборки.[23]
Есть убедительные доказательства того, что грузы in vivo транспортируются с помощью нескольких двигателей.[24][25][26][27]
Направление движения
Моторные белки перемещаются в определенном направлении по микротрубочке. Микротрубочки полярны; Это означает, что головки связываются с микротрубочкой только в одной ориентации, в то время как связывание АТФ дает каждому шагу свое направление посредством процесса, известного как застегивание шейного линкера.[28]
Ранее было известно, что кинезин перемещает груз к плюсовому (+) концу микротрубочки, также известный как антероградный транспорт / ортоградный транспорт.[29] Однако недавно было обнаружено, что в почкующихся дрожжевых клетках кинезин Cin8 (член семейства Kinesin-5) также может двигаться в направлении минус-конца или ретроградного транспорта. Это означает, что эти уникальные дрожжевые кинезиновые гомотетрамеры обладают новой способностью двигаться в двух направлениях.[30][31][32] До сих пор было показано, что кинезин движется к минус-концу только в группе, при этом двигатели скользят в антипараллельном направлении в попытке разделить микротрубочки.[33] Эта двойная направленность наблюдалась в идентичных условиях, когда свободные молекулы Cin8 движутся к минус-концу, а поперечно-сшивающий Cin8 перемещается к плюс-концам каждой поперечно сшитой микротрубочки. Одно конкретное исследование проверило скорость, с которой двигались двигатели Cin8, их результаты показали диапазон примерно 25-55 нм / с в направлении полюсов шпинделя.[34] На индивидуальной основе было обнаружено, что при изменении ионных условий двигатели Cin8 могут развивать скорость до 380 нм / с.[34] Предполагается, что двунаправленность дрожжевых моторов кинезина-5, таких как Cin8 и Cut7, является результатом связывания с другими моторами Cin8 и помогает выполнять роль динеина в почкующихся дрожжах, в отличие от человеческого гомолога этих моторов, плюс направил Eg5.[35] Это открытие в белках семейства кинезин-14 (таких какDrosophila melanogaster NCD, почкующиеся дрожжи KAR3 иArabidopsis thaliana ATK5) позволяет кинезину двигаться в противоположном направлении, к минус-концу микротрубочки.[36] Это не типично для кинезина, скорее, это исключение из нормального направления движения.

Другой тип моторного белка, известный какдинеины, двигайтесь к минус-концу микротрубочки. Таким образом, они транспортируют груз от периферии ячейки к центру. Примером этого может быть транспорт, происходящий от концевых бутонов нейронального аксона к телу клетки (соме). Это известно какретроградный транспорт.
Предлагаемые механизмы движения
Кинезин осуществляет транспорт, «идя» по микротрубочке. Было предложено два механизма для объяснения этого движения.
- В механизме «ручное управление» головки кинезина проходят одна мимо другой, чередуя ведущее положение.
- В механизме «дюймового червя» всегда ведет одна кинезиновая головка, продвигаясь вперед на шаг до того, как ведомая голова догоняет.
Несмотря на некоторые сохраняющиеся разногласия, растущее количество экспериментальных данных указывает на то, что передаточный механизм более вероятен.[37][38]
Связывание и гидролиз АТФ заставляют кинезин перемещаться по «качели» вокруг точки поворота.[39][40] Этот механизм качелей объясняет наблюдения, что связывание АТФ с отсутствием нуклеотидов, связанным с микротрубочками, приводит к наклону моторного домена кинезина относительно микротрубочки. Критически важно, что до этого наклона шейный линкер не может принять свою состыкованную с моторной головкой, обращенную вперед конформацию. АТФ-индуцированный наклон дает возможность шейному линкеру состыковаться в этой обращенной вперед конформации. Эта модель основана на моделях CRYO-EM структуры кинезина, связанной с микротрубочками, которые представляют начальное и конечное состояния процесса, но не могут разрешить точные детали перехода между структурами.
Теоретическое моделирование
Предложен ряд теоретических моделей кинезина молекулярного моторного белка.[41][42][43] Многие проблемы встречаются в теоретических исследованиях, учитывая остающуюся неопределенность в отношении роли белковых структур, точного способа преобразования энергии АТФ в механическую работу и роли тепловых флуктуаций. Это довольно активное направление исследований. Особенно необходимы подходы, которые лучше увязывают с молекулярной архитектурой белка и данными, полученными в результате экспериментальных исследований.
Динамика одиночных молекул уже хорошо описана.[44] но кажется, что эти наномасштабные машины обычно работают в больших командах. Недавние экспериментальные исследования показали, что кинезины, перемещаясь по микротрубочкам, взаимодействуют друг с другом,[45][46] короткодействующие и слабые притягивающие (1,6 ± 0,5 KBТ). Одна из разработанных моделей учитывает эти взаимодействия частиц,[44] где динамические скорости изменяются соответственно энергии взаимодействия. Если энергия положительна, скорость создания связей (q) будет выше, а скорость разрыва связей (r) будет ниже. Можно понять, что скорость входа и выхода в микротрубочке также будет изменяться под действием энергии (см. Рисунок 1 в ссылке 30). Если второй сайт занят, скорость входа будет α * q, а если предпоследний сайт занят, скорость выхода будет β * r. Этот теоретический подход согласуется с результатами моделирования Монте-Карло для этой модели, особенно для предельного случая очень большой отрицательной энергии. Обычный полностью асимметричный простой процесс исключения для результатов (или TASEP) может быть восстановлен из этой модели, сделав энергию равной нулю.

Митоз
В последние годы было обнаружено, что молекулярные моторы на основе микротрубочек (включая ряд кинезинов) играют роль в митоз (деление клеток). Кинезины важны для правильной длины веретена и участвуют в раздвижении микротрубочек внутри веретена во время прометафазы и метафазы, а также в деполимеризации минус-концов микротрубочек на центросомах во время анафазы.[47] В частности, белки семейства кинезин-5 действуют внутри веретена, раздвигая микротрубочки, в то время как Кинезин 13 семья действует, чтобы деполимеризовать микротрубочки.
Члены суперсемейства кинезинов
Члены суперсемейства кинезинов человека включают следующие белки, которые в стандартизированной номенклатуре, разработанной сообществом исследователей кинезина, разделены на 14 семейств, от кинезин-1 до кинезин-14:[12]
- 1А - KIF1A, 1Б - KIF1B, 1С - KIF1C = кинезин-3
- 2А - KIF2A, 2С - KIF2C = кинезин-13
- 3B - KIF3B или 3С - KIF3C , 3А - KIF3A = кинезин-2
- 4А - KIF4A, 4Б - KIF4B = кинезин-4
- 5А - KIF5A, 5Б - KIF5B, 5С - KIF5C = кинезин-1
- 6 – KIF6 = кинезин-9
- 7 – KIF7 = кинезин-4
- 9 – KIF9 = кинезин-9
- 11 – KIF11 = кинезин-5
- 12 – KIF12 = кинезин-12
- 13А - KIF13A, 13Б - KIF13B = кинезин-3
- 14 – KIF14 = кинезин-3
- 15 – KIF15 = кинезин-12
- 16B - KIF16B = кинезин-3
- 17 – KIF17 = кинезин-2
- 18А - KIF18A, 18Б - KIF18B = кинезин-8
- 19 – KIF19 = кинезин-8
- 20А - KIF20A, 20Б - KIF20B = кинезин-6
- 21А - KIF21A, 21Б - KIF21B = кинезин-4
- 22 – KIF22 = кинезин-10
- 23 – KIF23 = кинезин-6
- 24 – KIF24 = кинезин-13
- 25 – KIF25 = кинезин-14
- 26А - KIF26A, 26Б - KIF26B = кинезин-11
- 27 – KIF27 = кинезин-4
- C1 - KIFC1, C2 - KIFC2, C3 - KIFC3 = кинезин-14
легкие цепи кинезина-1:
белок, связанный с кинезином-2:
- KIFAP3 (также известный как КАП-1, КАП3)
Смотрите также
- Аксональный транспорт
- Дайнейн
- Внутрилагеллярный транспорт вдоль реснички
- Кинезин 8
- Кинезин 13
- КРП
- Молекулярный мотор
- Транспорт многомоторными белками
Рекомендации
- ^ Берг Дж., Тимочко Дж. Л., Страйер Л. (2002). «Кинезин и динеин движутся по микротрубочкам». Биохимия. 5-е издание.
- ^ Endow SA, Kull FJ, Лю Х. (октябрь 2010 г.). «Кинезины с первого взгляда». Журнал клеточной науки. 123 (Pt 20): 3420–4. Дои:10.1242 / jcs.064113. PMID 20930137.
- ^ Vale RD (февраль 2003 г.). «Молекулярный моторный набор инструментов для внутриклеточного транспорта». Клетка. 112 (4): 467–80. Дои:10.1016 / S0092-8674 (03) 00111-9. PMID 12600311. S2CID 15100327.
- ^ Вэйл Р.Д., Риз Т.С., Шитц МП (август 1985 г.). «Идентификация нового белка, генерирующего силу, кинезина, участвующего в подвижности микротрубочек». Клетка. 42 (1): 39–50. Дои:10.1016 / S0092-8674 (85) 80099-4. ЧВК 2851632. PMID 3926325.
- ^ Cole DG, Chinn SW, Wedaman KP, Hall K, Vuong T., Scholey JM (ноябрь 1993 г.). «Новый гетеротримерный белок, родственный кинезину, очищенный из яиц морского ежа». Природа. 366 (6452): 268–70. Bibcode:1993Натура.366..268C. Дои:10.1038 / 366268a0. PMID 8232586. S2CID 4367715.
- ^ Розенбаум Дж. Л., Уитман Г. Б. (ноябрь 2002 г.). «Внутрилагеллярный транспорт». Обзоры природы. Молекулярная клеточная биология. 3 (11): 813–25. Дои:10.1038 / nrm952. PMID 12415299. S2CID 12130216.
- ^ Ян Дж. Т., Лаймон Р. А., Гольдштейн Л. С. (март 1989 г.). «Трехдоменная структура тяжелой цепи кинезина, выявленная с помощью анализа последовательности ДНК и связывания микротрубочек». Клетка. 56 (5): 879–89. Дои:10.1016/0092-8674(89)90692-2. PMID 2522352. S2CID 44318695.
- ^ Айзава Х., Секин Ю., Такемура Р., Чжан З., Нангаку М., Хирокава Н. (декабрь 1992 г.). «Семья кинезина в центральной нервной системе мышей». Журнал клеточной биологии. 119 (5): 1287–96. Дои:10.1083 / jcb.119.5.1287. ЧВК 2289715. PMID 1447303.
- ^ Энос А.П., Моррис Н.Р. (март 1990 г.). «Мутация гена, кодирующего кинезиноподобный белок, блокирует ядерное деление у A. nidulans». Клетка. 60 (6): 1019–27. Дои:10.1016 / 0092-8674 (90) 90350-Н. PMID 2138511. S2CID 27420513.
- ^ Мелух ПБ, Роуз Мэриленд (март 1990 г.). «KAR3, ген, связанный с кинезином, необходимый для слияния ядер дрожжей». Клетка. 60 (6): 1029–41. Дои:10.1016 / 0092-8674 (90) 90351-E. PMID 2138512. S2CID 19660190.
- ^ Хирокава Н., Нода Ю., Танака Ю., Нива С. (октябрь 2009 г.). «Моторные белки суперсемейства кинезинов и внутриклеточный транспорт». Обзоры природы. Молекулярная клеточная биология. 10 (10): 682–96. Дои:10.1038 / nrm2774. PMID 19773780. S2CID 18129292.
- ^ а б Лоуренс CJ, Доу Р.К., Кристи К.Р., Кливленд Д.В., Доусон С.К., Эндов С.А., Голдштейн Л.С., Гудсон Х.В., Хирокава Н., Ховард Дж., Малмберг Р.Л., Макинтош-младший, Мики Х, Митчисон Т. , Schliwa M, Scholey JM, Vale RD, Walczak CE, Wordeman L (октябрь 2004 г.). «Стандартизированная номенклатура кинезинов». Журнал клеточной биологии. 167 (1): 19–22. Дои:10.1083 / jcb.200408113. ЧВК 2041940. PMID 15479732.
- ^ Хирокава Н., Пфистер К.К., Йорифудзи Х., Вагнер М.С., Брэди С.Т., Блум Г.С. (март 1989 г.). «Субмолекулярные домены кинезина бычьего мозга, идентифицированные с помощью электронной микроскопии и декорирования моноклональных антител». Клетка. 56 (5): 867–78. Дои:10.1016/0092-8674(89)90691-0. PMID 2522351. S2CID 731898.
- ^ PDB: 1BG2; Кулл Ф.Дж., Саблин Е.П., Лау Р., Флеттерик Р.Дж., Вейл Р.Д. (апрель 1996 г.). «Кристаллическая структура моторного домена кинезина обнаруживает структурное сходство с миозином». Природа. 380 (6574): 550–5. Bibcode:1996 Натур. 380..550J. Дои:10.1038 / 380550a0. ЧВК 2851642. PMID 8606779.
- ^ а б Каан Х.Й., Хакни Д.Д., Козельски Ф. (август 2011 г.). «Структура моторно-хвостового комплекса кинезин-1 раскрывает механизм аутоингибирования». Наука. 333 (6044): 883–5. Bibcode:2011Наука ... 333..883K. Дои:10.1126 / science.1204824. ЧВК 3339660. PMID 21836017.
- ^ Стюарт Р. Дж., Талер Дж. П., Гольдштейн Л. С. (июнь 1993 г.). «Направление движения микротрубочек является внутренним свойством моторных доменов тяжелой цепи кинезина и белка ncd дрозофилы». Труды Национальной академии наук Соединенных Штатов Америки. 90 (11): 5209–13. Bibcode:1993ПНАС ... 90.5209С. Дои:10.1073 / пнас.90.11.5209. ЧВК 46685. PMID 8506368.
- ^ Верхей KJ, Хаммонд JW (ноябрь 2009 г.). «Управление движением: регулирование кинезиновых моторов». Обзоры природы. Молекулярная клеточная биология. 10 (11): 765–77. Дои:10.1038 / nrm2782. PMID 19851335. S2CID 10713993.
- ^ Сиддики Н., Светслот А.Дж., Бахманн А., Рот Д., Хуссейн Н., Брандт Дж. И др. (Июнь 2019). «PTPN21 и Hook3 снимают аутоингибирование KIF1C и активируют внутриклеточный транспорт». Nature Communications. 10 (1): 2693. Bibcode:2019НатКо..10.2693S. Дои:10.1038 / s41467-019-10644-9. ЧВК 6584639. PMID 31217419.
- ^ Blasius TL, Cai D, Jih GT, Toret CP, Verhey KJ (январь 2007 г.). «Два связывающих партнера взаимодействуют, чтобы активировать молекулярный мотор кинезин-1». Журнал клеточной биологии. 176 (1): 11–7. Дои:10.1083 / jcb.200605099. ЧВК 2063617. PMID 17200414.
- ^ Шнитцер М.Дж., Блок С.М. (июль 1997 г.). «Кинезин гидролизует один АТФ за шаг 8 нм». Природа. 388 (6640): 386–90. Bibcode:1997Натура.388..386S. Дои:10.1038/41111. PMID 9237757. S2CID 4363000.
- ^ Вэйл Р. Д., Миллиган Р. А. (апрель 2000 г.). «Как все движется: заглянуть под капот молекулярных моторных белков». Наука. 288 (5463): 88–95. Bibcode:2000Sci ... 288 ... 88V. Дои:10.1126 / science.288.5463.88. PMID 10753125.
- ^ Mather WH, Fox RF (октябрь 2006 г.). «Смещенный шаговый механизм кинезина: усиление молнии шейного линкера». Биофизический журнал. 91 (7): 2416–26. Bibcode:2006BpJ .... 91.2416M. Дои:10.1529 / biophysj.106.087049. ЧВК 1562392. PMID 16844749.
- ^ Gaudin R, de Alencar BC, Jouve M, Bèrre S, Le Bouder E, Schindler M, Varthaman A, Gobert FX, Benaroch P (октябрь 2012 г.). «Критическая роль кинезина KIF3A в жизненном цикле ВИЧ в первичных макрофагах человека». Журнал клеточной биологии. 199 (3): 467–79. Дои:10.1083 / jcb.201201144. ЧВК 3483138. PMID 23091068.
- ^ Гросс СП, Вершинин М, Шубейта Г.Т. (июнь 2007 г.). «Грузовой транспорт: два мотора иногда лучше, чем один». Текущая биология. 17 (12): R478–86. Дои:10.1016 / j.cub.2007.04.025. PMID 17580082. S2CID 8791125.
- ^ Хэнкок WO (август 2008 г.). «Внутриклеточный транспорт: кинезины работают вместе». Текущая биология. 18 (16): R715–7. Дои:10.1016 / j.cub.2008.07.068. PMID 18727910. S2CID 7540556.
- ^ Кунвар А., Вершинин М., Сюй Дж., Гросс С.П. (август 2008 г.). «Шаги, стробирование деформации и неожиданная кривая силы и скорости для транспорта с несколькими двигателями». Текущая биология. 18 (16): 1173–83. Дои:10.1016 / j.cub.2008.07.027. ЧВК 3385514. PMID 18701289.
- ^ Клумпп С., Липовски Р. (ноябрь 2005 г.). «Совместная транспортировка грузов несколькими молекулярными моторами». Труды Национальной академии наук Соединенных Штатов Америки. 102 (48): 17284–9. arXiv:q-bio / 0512011. Bibcode:2005PNAS..10217284K. Дои:10.1073 / pnas.0507363102. ЧВК 1283533. PMID 16287974.
- ^ Rice S, Lin AW, Safer D, Hart CL, Naber N, Carragher BO, Cain SM, Pechatnikova E, Wilson-Kubalek EM, Whittaker M, Pate E, Cooke R, Taylor EW, Milligan RA, Vale RD (декабрь 1999 г.) . «Структурное изменение моторного белка кинезина, которое управляет моторикой». Природа. 402 (6763): 778–84. Bibcode:1999Натура.402..778R. Дои:10.1038/45483. PMID 10617199. S2CID 573909.
- ^ Лодиш Х, Берк А, Зипурски С.Л., Мацудаира П., Балтимор Д., Дарнелл Дж. (2000). «Кинезин, динеин и внутриклеточный транспорт». Цитировать журнал требует
| журнал =(помощь) - ^ Роосталу Дж., Хентрих С., Билинг П., Телли И. А., Шибель Е., Суррей Т. (апрель 2011 г.). «Направленное переключение кинезина Cin8 через моторную муфту». Наука. 332 (6025): 94–9. Bibcode:2011Научный ... 332 ... 94R. Дои:10.1126 / science.1199945. PMID 21350123. S2CID 90739364.
- ^ Фаллесен Т., Роосталу Дж., Дуэльберг С., Прюсснер Дж., Суррей Т. (ноябрь 2017 г.). «Ансамбли двунаправленного кинезина Cin8 создают аддитивные силы в обоих направлениях движения». Биофизический журнал. 113 (9): 2055–2067. Bibcode:2017BpJ ... 113.2055F. Дои:10.1016 / j.bpj.2017.09.006. ЧВК 5685778. PMID 29117528.
- ^ Эдамацу М (март 2014 г.). «Двунаправленная подвижность кинезина-5 делящихся дрожжей, Cut7». Сообщения о биохимических и биофизических исследованиях. 446 (1): 231–4. Дои:10.1016 / j.bbrc.2014.02.106. PMID 24589736.
- ^ Роосталу Дж., Хентрих С., Билинг П., Телли И. А., Шибель Е., Суррей Т. (апрель 2011 г.). «Направленное переключение кинезина Cin8 через моторную муфту». Наука. 332 (6025): 94–9. Bibcode:2011Научный ... 332 ... 94R. Дои:10.1126 / science.1199945. PMID 21350123. S2CID 90739364.
- ^ а б Герсон-Гурвиц А., Тиде С., Мовшович Н., Фридман В., Подольская М., Даниэли Т. и др. (Ноябрь 2011 г.). «Направленность отдельных моторов кинезина-5 Cin8 модулируется петлей 8, ионной силой и геометрией микротрубочек». Журнал EMBO. 30 (24): 4942–54. Дои:10.1038 / emboj.2011.403. ЧВК 3243633. PMID 22101328.
- ^ Валентин М.Т., Фордайс П.М., Блок С.М. (декабрь 2006 г.). "Eg5 усиливается!". Отделение клеток. 1 (1): 31. Дои:10.1186/1747-1028-1-31. ЧВК 1716758. PMID 17173688.
- ^ Амброуз Дж. К., Ли В., Маркус А., Ма Х., Сир Р. (апрель 2005 г.). «Направленный на минус-конец кинезин с активностью трекингового белка на плюс-конце участвует в морфогенезе веретена». Молекулярная биология клетки. 16 (4): 1584–92. Дои:10.1091 / mbc.e04-10-0935. ЧВК 1073643. PMID 15659646.
- ^ Йылдыз А., Томишиге М., Вале Р.Д., Сельвин П.Р. (январь 2004 г.). «Кинезин ходит из рук в руки». Наука. 303 (5658): 676–8. Bibcode:2004Наука ... 303..676л. Дои:10.1126 / science.1093753. PMID 14684828. S2CID 30529199.
- ^ Asbury CL (февраль 2005 г.). «Кинезин: самый крошечный двуногий в мире». Текущее мнение в области клеточной биологии. 17 (1): 89–97. Дои:10.1016 / j.ceb.2004.12.002. PMID 15661524.
- ^ Резюме Синделара, Даунинг К. Х. (март 2010 г.). «Механизм на атомном уровне для активации молекулярных моторов кинезина». Труды Национальной академии наук Соединенных Штатов Америки. 107 (9): 4111–6. Bibcode:2010ПНАС..107.4111С. Дои:10.1073 / pnas.0911208107. ЧВК 2840164. PMID 20160108.
- ^ Lay Summary (18 февраля 2010 г.). «Самый маленький мотор в жизни, грузовой перевозчик ячеек, движется как качели». PhysOrg.com. Получено 31 мая 2013.
- ^ Ацбергер П.Дж., Пескин С.С. (январь 2006 г.). «Модель броуновской динамики кинезина в трех измерениях, включающая профиль силы растяжения грузового троса наматываемой спирали». Вестник математической биологии. 68 (1): 131–60. arXiv:0910.5753. Дои:10.1007 / s11538-005-9003-6. PMID 16794924. S2CID 13534734.
- ^ Пескин С.С., Остер Г. (апрель 1995 г.). «Скоординированный гидролиз объясняет механическое поведение кинезина». Биофизический журнал. 68 (4 Прил.): 202S – 210S, обсуждение 210S – 211S. ЧВК 1281917. PMID 7787069.
- ^ Могильнер А., Фишер А.Дж., Баскин Р.Дж. (июль 2001 г.). «Структурные изменения в шейном линкере кинезина объясняют зависимость механического цикла двигателя от нагрузки». Журнал теоретической биологии. 211 (2): 143–57. Дои:10.1006 / jtbi.2001.2336. PMID 11419956.
- ^ а б Селис-Гарза Д., Теймури Х., Коломейский А.Б. (2015). «Корреляции и симметрия взаимодействий влияют на коллективную динамику молекулярных моторов». Журнал статистической механики: теория и эксперимент. 2015 (4): P04013. arXiv:1503.00633. Bibcode:2015JSMTE..04..013C. Дои:10.1088 / 1742-5468 / 2015/04 / p04013. S2CID 14002728.
- ^ Зейтц А., Суррей Т. (январь 2006 г.). «Процессивное движение одиночных кинезинов по переполненным микротрубочкам, визуализированное с помощью квантовых точек». Журнал EMBO. 25 (2): 267–77. Дои:10.1038 / sj.emboj.7600937. ЧВК 1383520. PMID 16407972.
- ^ Вилфан А., Фрей Э., Швабль Ф., Тормэлен М., Сонг Ю. Х., Мандельков Э. (октябрь 2001 г.). «Динамика и кооперативность украшения микротрубочек моторным белком кинезином». Журнал молекулярной биологии. 312 (5): 1011–26. Дои:10.1006 / jmbi.2001.5020. PMID 11580246.
- ^ Гошима Дж., Вале РД (август 2005 г.). «Зависимая от клеточного цикла динамика и регуляция митотических кинезинов в клетках S2 Drosophila». Молекулярная биология клетки. 16 (8): 3896–907. Дои:10.1091 / mbc.E05-02-0118. ЧВК 1182325. PMID 15958489.
дальнейшее чтение
- Лоуренс С.Дж., Доу Р.К., Кристи К.Р., Кливленд Д.В., Доусон С.К., Эндов С.А., Голдштейн Л.С., Гудсон Х.В., Хирокава Н., Ховард Дж., Малмберг Р.Л., Макинтош-младший, Мики Х., Митчисон Т.Дж., Окада Й., Редди А.С., Сакстон В.М. , Schliwa M, Scholey JM, Vale RD, Walczak CE, Wordeman L (октябрь 2004 г.). «Стандартизированная номенклатура кинезинов». Журнал клеточной биологии. 167 (1): 19–22. Дои:10.1083 / jcb.200408113. ЧВК 2041940. PMID 15479732.
внешняя ссылка
- MBInfo - Кинезин транспортирует грузы по микротрубочкам
- Анимированная модель кинезиновой ходьбы
- Семинар Рона Вейла: «Молекулярные моторные белки»
- Анимация движения кинезина Библиотека изображений ASCB
- Мерфи, В.Ф. (2004-05-12). «Движение на основе микротрубочек». fabric.medicalengineer.co.uk. Архивировано из оригинал на 2007-07-22. Получено 2015-12-10.
- Внутренняя жизнь клетки, трехмерная анимация с кинезином, транспортирующим везикулу
- Домашняя страница Kinesin
- Кинезин в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- EC 3.6.4.4
- EC 3.6.4.5
- Структуры кинезина для трехмерной электронной микроскопии из EM Data Bank (EMDB)