Аксональный транспорт - Axonal transport
Аксональный транспорт, также называемый аксоплазматический транспорт или же аксоплазматический поток, это клеточный процесс отвечает за перемещение митохондрии, липиды, синаптические везикулы, белки, и другие органеллы в и из нейрон с Тело клетки, сквозь цитоплазма своего аксон называется аксоплазма.[1] Поскольку некоторые аксоны имеют длину порядка метра, нейроны не могут полагаться на диффузию, чтобы нести продукты ядра и органелл к концам своих аксонов. Аксональный транспорт также отвечает за перемещение молекул, предназначенных для деградации, от аксона обратно в тело клетки, где они расщепляются лизосомы.[2]
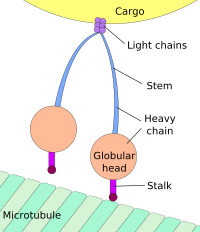
Движение к телу клетки называется ретроградным транспортом, а движение к синапс называется антероградным транспортом.[3][4]
Механизм

Подавляющее большинство аксональных белков синтезируется в теле нейрональных клеток и транспортируется по аксонам. Немного трансляция мРНК был продемонстрирован в аксонах.[5][6] Транспорт аксонов происходит на протяжении всей жизни нейрона и необходим для его роста и выживания. Микротрубочки (сделано из тубулин ) проходят по длине аксона и служат основными «дорожками» цитоскелета для транспортировки. Кинезин и динеин находятся моторные белки которые перемещают грузы в антерограде (вперед от сома к кончику аксона) и ретроградному (назад к соме (телу клетки)) направлениям соответственно. Моторные белки связывают и транспортируют несколько различных грузов, включая митохондрии, цитоскелет полимеры, аутофагосомы и синаптические везикулы содержащий нейротрансмиттеры.
Аксональный транспорт может быть быстрым или медленным, а также антероградным (от тела клетки) или ретроградным (перенос материалов от аксона к телу клетки).
Быстрый и медленный транспорт
Везикулярные грузы перемещаются относительно быстро (50–400 мм / день), тогда как транспортировка растворимых (цитозольных) и цитоскелетных белков занимает намного больше времени (перемещение менее 8 мм / день).[7] Основной механизм быстрого аксонального транспорта был понят в течение десятилетий, но механизм медленного аксонального транспорта стал ясен только недавно в результате продвинутых методы визуализации.[8] Методы флуоресцентной маркировки (например, флуоресцентная микроскопия ) сделали возможным прямую визуализацию транспорта в живых нейронах. (Смотрите также: Антероградное отслеживание.)
Недавние исследования показали, что движение цитоскелетных «медленных» грузов на самом деле происходит быстро, но в отличие от быстрых грузов, они часто останавливаются, что значительно снижает общую скорость транзита. Этот механизм известен как модель медленного аксонального транспорта «Stop and Go» и был широко подтвержден для транспорта нейрофиламентов цитоскелетного белка.[9] Движение растворимых (цитозольных) грузов является более сложным, но, по-видимому, имеет аналогичную основу, когда растворимые белки организуются в мультибелковые комплексы, которые затем передаются посредством временных взаимодействий с более быстро движущимися грузами, перемещающимися в быстром аксональном транспорте.[10][11][12] Аналогия - разница в тарифах на проезд между местными и скоростными поездами метро. Хотя оба типа поездов движутся между станциями с одинаковой скоростью, местному поезду требуется гораздо больше времени, чтобы добраться до конца линии, потому что он останавливается на каждой станции, тогда как экспресс делает всего несколько остановок по пути.
Антероградный транспорт
Антероградный (также называемый «ортоградным») перенос - это движение молекул / органелл наружу, от Тело клетки (также называемый сома ) к синапс или же клеточная мембрана.
Антероградное движение отдельных грузов (в транспортные везикулы ) как быстрых, так и медленных компонентов вдоль микротрубочка[4] опосредовано кинезины.[2] Некоторые кинезины участвуют в медленном транспорте,[8] хотя механизм создания «пауз» в пути медленных составных грузов пока неизвестен.
Существует два класса медленного антероградного транспорта: медленный компонент a (SCa), который несет в основном микротрубочки, и нейрофиламенты 0,1-1 миллиметра в день, и медленный компонент b (SCb), который переносит более 200 различных белков и актин со скоростью до 6 миллиметров в день.[8] Медленный компонент b, который также несет актин, транспортируется в аксонах клеток сетчатки со скоростью 2-3 миллиметра в день.
Во время реактивации из-за задержки вирус простого герпеса (HSV) входит в свой литический цикл, и использует механизмы антероградного транспорта для миграции из ганглии задних корешков нейроны кожи или слизистой оболочки, на которые он впоследствии влияет.[13]
Рецептор карго для моторов антероградного транспорта, кинезины, был идентифицирован как белок-предшественник амилоида (АРР), родительский белок, который производит сенильные бляшки, обнаруживаемые при болезни Альцгеймера.[14] Пептид из 15 аминокислот на карбоксильном конце цитоплазмы АРР связывается с высоким сродством с обычным кинезином-1 и опосредует транспорт экзогенного груза в гигантском аксоне кальмара.[15]
Марганец, контрастное вещество для T1-взвешенная МРТ, перемещается антероградным транспортом после стереотаксической инъекции в мозг экспериментальных животных и, таким образом, выявляет схему с помощью МРТ всего мозга у живых животных, как впервые было сделано Робией Паутлер, Элейн Бирер и Расс Джейкобс. Исследования на мышах с нокаутом кинезин-легкая цепь-1 показали, что Mn2+ перемещается посредством транспорта на основе кинезина в зрительном нерве и в головном мозге. Транспорт как в проекциях гиппокампа, так и в зрительном нерве также зависит от APP.[16] Транспорт из гиппокампа в передний мозг снижается с возрастом, а место назначения изменяется из-за наличия бляшек болезни Альцгеймера.[17]
Ретроградный транспорт
Ретроградный транспорт перемещает молекулы / органеллы от концов аксона к Тело клетки. Ретроградный аксональный транспорт опосредуется цитоплазматическим динеин, и используется, например, для отправки химических сообщений и эндоцитоз продукты направляются в эндолизосомы от аксона обратно в клетку.[2] Работает в среднем in vivo скорость около 2 мкм / сек,[18][19] быстрый ретроградный транспорт может преодолевать 10-20 сантиметров в день.[2]
Быстрый ретроградный транспорт возвращает использованные синаптические пузырьки и другие материалы в сому и сообщает соме о состояниях на терминалах аксонов. Ретроградный транспорт переносит сигналы выживания из синапса обратно в тело клетки, такие как TRK, рецептор фактора роста нервов.[20] Некоторые патогены используют этот процесс для вторжения в нервную систему. Они входят в дистальные кончики аксона и перемещаются к соме ретроградным транспортом. Примеры включают столбнячный токсин и вирусы простого герпеса, бешенства и полиомиелита. При таких инфекциях задержка между инфицированием и появлением симптомов соответствует времени, необходимому для того, чтобы патогены достигли соматических клеток.[21] Вирус простого герпеса перемещается по аксонам в обоих направлениях в зависимости от своего жизненного цикла, при этом полярность ретроградного транспорта доминирует для поступающих капсидов.[22]
Последствия прерывания
Всякий раз, когда транспорт аксонов подавляется или прерывается, нормальная физиология становится патофизиологией, и накопление аксоплазмы, называемое аксональный сфероид, может привести. Поскольку транспорт аксонов может быть нарушен множеством способов, аксональные сфероиды можно увидеть при многих различных классах заболеваний, включая генетические, травматические, ишемические, инфекционные, токсические, дегенеративные и специфические заболевания белого вещества называется лейкоэнцефалопатии. Несколько редких нейродегенеративные заболевания связаны с генетические мутации в моторных белках, кинезин и динеин, и в этих случаях вероятно, что транспорт аксонов является ключевым игроком в опосредовании патологии.[23][24] Дисфункциональный аксональный транспорт также связан со спорадическими (распространенными) формами нейродегенеративных заболеваний, таких как Болезнь Альцгеймера и Болезнь Паркинсона.[8] Это в основном связано с многочисленными наблюдениями, что большие скопления аксонов неизменно наблюдаются в пораженных нейронах, и что гены, о которых известно, что они играют роль в семейных формах этих заболеваний, также предположительно играют роль в нормальном аксональном транспорте. Однако прямых доказательств участия аксонального транспорта в последних заболеваниях мало, и другие механизмы (такие как прямая синаптотоксичность) могут быть более актуальными.
Остановка аксоплазматического кровотока на краю ишемической области при сосудистых ретинопатиях приводит к набуханию нервных волокон, которое приводит к мягкому экссудату или ватным пятнам.
Поскольку аксон зависит от аксоплазматического транспорта жизненно важных белков и материалов, повреждения, такие как диффузное повреждение аксонов который прерывает транспорт, вызовет дистальный аксон дегенерирует в процессе, называемом Валлеровское вырождение. Противораковые препараты которые мешают раку, изменяя микротрубочки (которые необходимы для деление клеток ) повреждают нервы, потому что микротрубочки необходимы для транспорта аксонов.
Инфекционное заболевание
В вирус бешенства достигает центральной нервной системы ретроградным потоком аксоплазмы.[25] Столбнячный нейротоксин усваивается в нервномышечное соединение путем связывания нидоген белков и ретроградно транспортируется к соме в сигнальных эндосомах.[26] Нейротропные вирусы, такие как герпесвирусы, перемещаются внутри аксонов с помощью клеточного транспортного механизма, как было показано в работе группы Элейн Беарэр.[27][28] Другие инфекционные агенты также подозреваются в использовании аксонального транспорта.[29] В настоящее время считается, что такие инфекции способствуют развитию болезни Альцгеймера и других нейродегенеративных неврологических расстройств.[30][31]
Смотрите также
Рекомендации
- ^ Сабри Дж., О'Коннор Т.П., Киршнер М.В. (июнь 1995 г.). «Аксональный транспорт тубулина в пионерных нейронах Ti1 in situ». Нейрон. 14 (6): 1247–56. Дои:10.1016/0896-6273(95)90271-6. PMID 7541635.
- ^ а б c d Озтас Э (2003). «Нейрональное отслеживание» (PDF). Нейроанатомия. 2: 2–5. В архиве (PDF) из оригинала от 25 октября 2005 г.
- ^ Карп Г, ван дер Гир П. Клеточная и молекулярная биология: концепции и эксперименты (4-е изд.). Джон Вили. п.344. ISBN 978-0-471-46580-5.
- ^ а б Медведь М.Ф., Коннорс Б.В., Парадсо Массачусетс (2007). Неврология: исследование мозга (3-е изд.). Липпинкотт Уильямс и Уилкинс. п.41. ISBN 978-0-7817-6003-4.
- ^ Джустетто М., Хегде А.Н., Си К., Касадио А., Инокучи К., Пей В., Кандел Э. Р., Шварц Дж. Х. (ноябрь 2003 г.). «Аксональный транспорт мРНК фактора элонгации трансляции 1альфа эукариот связывает транскрипцию в ядре с долгосрочным облегчением в синапсе». Труды Национальной академии наук Соединенных Штатов Америки. 100 (23): 13680–5. Bibcode:2003ПНАС..10013680Г. Дои:10.1073 / pnas.1835674100. ЧВК 263873. PMID 14578450.
- ^ Си К. Джустетто Си К., Джустетто М., Эткин А., Хсу Р., Янисевич А. М., Миньячи М. С., Ким Дж. Х., Чжу Х., Кандел Е. Р. (декабрь 2003 г.). «Нейрональная изоформа CPEB регулирует локальный синтез белка и стабилизирует синапс-специфическое долгосрочное облегчение при аплизии». Клетка. 115 (7): 893–904. Дои:10.1016 / s0092-8674 (03) 01021-3. PMID 14697206. S2CID 15552012.
- ^ Мадай, Сандра; Twelvetrees, Элисон Э .; Мугамян, Армен Дж .; Хольцбаур, Эрика Л.Ф. (октябрь 2014 г.). «Аксональный транспорт: специфические для груза механизмы движения и регуляции». Нейрон. 84 (2): 292–309. Дои:10.1016 / j.neuron.2014.10.019. ЧВК 4269290. PMID 25374356.
- ^ а б c d Рой С., Чжан Б., Ли В.М., Трояновский JQ (январь 2005 г.). «Дефекты аксонального транспорта: общая тема нейродегенеративных заболеваний». Acta Neuropathologica. 109 (1): 5–13. Дои:10.1007 / s00401-004-0952-x. PMID 15645263. S2CID 11635065.
- ^ Браун А (март 2003 г.). «Аксональный транспорт мембранных и немембранозных грузов: единая перспектива». Журнал клеточной биологии. 160 (6): 817–21. Дои:10.1083 / jcb.200212017. ЧВК 2173776. PMID 12642609.
- ^ Скотт Д.А., Дас У, Тан И, Рой С. (май 2011 г.). «Механистическая логика, лежащая в основе аксонального транспорта цитозольных белков». Нейрон. 70 (3): 441–54. Дои:10.1016 / j.neuron.2011.03.022. ЧВК 3096075. PMID 21555071.
- ^ Рой С., Винтон М.Дж., Блэк М.М., Трояновский Д.К., Ли В.М. (март 2007 г.). «Быстрый и прерывистый котранспорт медленных белков компонента-b». Журнал неврологии. 27 (12): 3131–8. Дои:10.1523 / JNEUROSCI.4999-06.2007. ЧВК 6672457. PMID 17376974.
- ^ Кузнецов А.В. (2011). «Аналитическое решение уравнений, описывающих медленный аксональный транспорт на основе гипотезы остановки и движения». Центральноевропейский журнал физики. 9 (3): 662–673. Bibcode:2011CEJPh ... 9..662K. Дои:10.2478 / s11534-010-0066-0.
- ^ Голландский ди-джей, Миранда-Саксена М., Боадл Р.А., Армати П., Каннингем А.Л. (октябрь 1999 г.). «Антероградный транспорт белков вируса простого герпеса в аксонах периферических нейронов плода человека: исследование с помощью иммуноэлектронной микроскопии». Журнал вирусологии. 73 (10): 8503–11. Дои:10.1128 / JVI.73.10.8503-8511.1999. ЧВК 112870. PMID 10482603.
- ^ Сатпуте-Кришнан П., ДеГиоргис Дж. А., Конли М. П., Джанг М., Носитель Е. Л. (октябрь 2006 г.). «Зип-код пептида, достаточный для антероградного транспорта в белке-предшественнике амилоида». Труды Национальной академии наук Соединенных Штатов Америки. 103 (44): 16532–7. Bibcode:2006ПНАС..10316532С. Дои:10.1073 / pnas.0607527103. ЧВК 1621108. PMID 17062754.
- ^ Seamster PE, Loewenberg M, Pascal J, Chauviere A, Gonzales A, Cristini V, Bearer EL (октябрь 2012 г.). «Количественные измерения и моделирование грузомоторных взаимодействий при быстром переносе в живом аксоне». Физическая биология. 9 (5): 055005. Bibcode:2012ФБио ... 9e5005S. Дои:10.1088/1478-3975/9/5/055005. ЧВК 3625656. PMID 23011729.
- ^ Галлахер Дж. Дж., Чжан Икс, Зиомек Дж. Дж., Джейкобс Р. Э., Несущий Е. Л. (апрель 2012 г.). «Дефицит аксонального транспорта в цепи на основе гиппокампа и зрительный путь у животных с нокаутом APP, подтвержденный МРТ с усилением марганца». NeuroImage. 60 (3): 1856–66. Дои:10.1016 / j.neuroimage.2012.01.132. ЧВК 3328142. PMID 22500926.
- ^ Бирер Э.Л., Манифолд-Уиллер BC, Медина CS, Гонсалес АГ, Чавес Флорида, Якобс RE (октябрь 2018 г.) «Изменения функциональной схемы в стареющем мозге и влияние мутировавшего выражения АРР». Нейробиология старения. 70: 276–290. Дои:10.1016 / j.neurobiolaging.2018.06.018. ЧВК 6159914. PMID 30055413.
- ^ Гиббс К.Л., Кальмар Б., Сани Дж. Н., Гринсмит Л., Скьяво Г. (январь 2016 г.). «In vivo визуализация аксонального транспорта в моторных и сенсорных нейронах мышей». Журнал методов неврологии. 257: 26–33. Дои:10.1016 / j.jneumeth.2015.09.018. ЧВК 4666412. PMID 26424507.
- ^ Сани J, Скьяво G (2016). «Старше, но не медленнее: старение не влияет на динамику аксонального транспорта сигнальных эндосом in vivo». Имеет значение. 2 (6). Дои:10.19185 / questions.201605000018.
- ^ Цуй Б., Ву С., Чен Л., Рамирес А., Биэрр Э. Л., Ли В. П., Мобли В. К., Чу С. (август 2007 г.). «Одно за другим, отслеживание транспорта аксонов NGF с использованием квантовых точек». Труды Национальной академии наук Соединенных Штатов Америки. 104 (34): 13666–71. Bibcode:2007PNAS..10413666C. Дои:10.1073 / pnas.0706192104. ЧВК 1959439. PMID 17698956.
- ^ Саладин, Кеннет. Анатомия и физиология: единство формы и функции. Шестое. Нью-Йорк: Макгроу-Хилл, 2010. 445. Печать.
- ^ Биэрр Е.Л., Брейкфилд XO, Шубак Д., Риз Т.С., ЛаВейл Дж. Х. (июль 2000 г.). «Ретроградный аксональный транспорт вируса простого герпеса: доказательства единственного механизма и роли тегумента». Труды Национальной академии наук Соединенных Штатов Америки. 97 (14): 8146–50. Bibcode:2000PNAS ... 97.8146B. Дои:10.1073 / пнас.97.14.8146. ЧВК 16684. PMID 10884436.
- ^ Maday S, Twelvetrees AE, Мугамян AJ, Holzbaur EL (октябрь 2014 г.). «Аксональный транспорт: грузо-специфические механизмы движения и регуляции». Нейрон. 84 (2): 292–309. Дои:10.1016 / j.neuron.2014.10.019. ЧВК 4269290. PMID 25374356.
- ^ Сани Дж. Н., Россор А. М., Товарищи А. Д., Тосолини А. П., Скьяво Г. (декабрь 2019 г.). «Аксональный транспорт и неврологические заболевания». Нат Рев Нейрол. 15 (12): 691–703. Дои:10.1038 / s41582-019-0257-2. PMID 31558780. S2CID 203437348.
- ^ Митрабхакди Э., Шуангшоти С., Ваннакрайрот П., Льюис Р.А., Сусуки К., Лаотаматас Дж., Хемачуда Т. (ноябрь 2005 г.). «Различие в нейропатогенетических механизмах при бешеном и паралитическом бешенстве человека». Журнал неврологических наук. 238 (1–2): 3–10. Дои:10.1016 / j.jns.2005.05.004. PMID 16226769. S2CID 25509462.
- ^ Bercsenyi K, Schmieg N, Bryson JB, Wallace M, Caccin P, Golding M, Zanotti G, Greensmith L, Nischt R, Schiavo G (ноябрь 2014 г.). «Поступление токсина столбняка. Нидогены являются терапевтическими мишенями для профилактики столбняка» (PDF). Наука. 346 (6213): 1118–23. Дои:10.1126 / science.1258138. PMID 25430769. S2CID 206560426.
- ^ Сатпуте-Кришнан П., ДеГиоргис Дж. А., предъявитель Е. Л. (декабрь 2003 г.). «Быстрый антероградный транспорт вируса простого герпеса: роль белка-предшественника амилоида болезни Альцгеймера». Ячейка старения. 2 (6): 305–18. Дои:10.1046 / j.1474-9728.2003.00069.x. ЧВК 3622731. PMID 14677633.
- ^ Ченг С.Б., Ферланд П., Вебстер П., Бирер Е.Л. (март 2011 г.). «Вирус простого герпеса танцует с белком-предшественником амилоида при выходе из клетки». PLOS ONE. 6 (3): e17966. Bibcode:2011PLoSO ... 617966C. Дои:10.1371 / journal.pone.0017966. ЧВК 3069030. PMID 21483850.
- ^ Носитель Э.Л., Сатпуте-Кришнан П. (сентябрь 2002 г.). «Роль цитоскелета в жизненном цикле вирусов и внутриклеточных бактерий: треки, двигатели и машины для полимеризации». Текущие целевые показатели по лекарствам. Инфекционные заболевания. 2 (3): 247–64. Дои:10.2174/1568005023342407. ЧВК 3616324. PMID 12462128.
- ^ Ицхаки Р.Ф., Токарный Р., Балин Б.Дж., Болл М.Дж., Несущий Е.Л., Браак Х. и др. (2016). «Микробы и болезнь Альцгеймера». Журнал болезни Альцгеймера. 51 (4): 979–84. Дои:10.3233 / JAD-160152. ЧВК 5457904. PMID 26967229.
- ^ «Нет места лучше асфальта для этих выносливых микробов». Новый ученый. 206 (2757): 15. 2010. Дои:10.1016 / s0262-4079 (10) 60991-8.