Лили вирус X - Lily virus X
| Лили вирус X | |
|---|---|
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область: | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Kitrinoviricota |
| Учебный класс: | Alsuviricetes |
| Заказ: | Tymovirales |
| Семья: | Alphaflexiviridae |
| Род: | Потексвирус |
| Разновидность: | Лили вирус X |
Лили вирус X (LVX) является патогенным ssRNA (+) растение вирус семьи Alphaflexiviridae и порядок Tymovirales. Типовой вид рода Потексвирус.
Описание
LVX описан в 4-м отчете ICTV (1982). Он обнаружен в основном в лилиях, хотя к нему чувствительны и другие растения, и считается, что вирус передается только механическим путем. Нет известных векторов; однако применение инсектицидов эффективно уменьшило распространение LVX, из чего можно сделать вывод, что передача вируса опосредована насекомыми.[1] Симптомы этого вируса пока неизвестны, что затрудняет оценку степени естественного заражения и распространения.
Структура

LVX - вирус без оболочки со спиральной симметрией. Считается, что все потексвирусы, включая LVX, содержат чуть менее 9 белковых субъединиц на виток спирали.[3] Этот паттерн образования нуклеокапсида приводит к тому, что нуклеокапсид становится удлиненным, гибким, нитчатым вирусом, как и большинство вирусов растений. В отличие от других потексвирусов (средняя длина 550 нм) LVX имеет длину 470 нм и ширину 13 нм.[4] LVX отличается от лилии бессимптомно карлавирус (LSV) серологическими тестами, такими как тесты с маркировкой иммунного золота.[5]
Геном
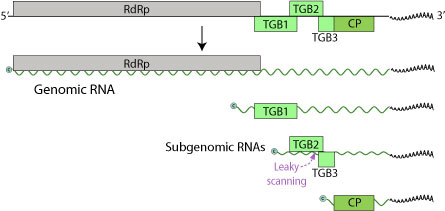
Геном LVX состоит из одной однонитевой (+) последовательности РНК размером 5,9-7 тыс. Пар оснований. В геноме всего три Открытые рамки для чтения (ORF), кодирующие пять белков (RdRp, TGBp1, TGBp2, TGBp3, CP).[6] 5 ’конец закрыт, а 3’ конец полиаденилирован.
Этих белков 24 килодальтон (кДа), 12 кДа и 22 кДа, третий из которых представляет собой белок оболочки, а также кодирует РНК-зависимая РНК-полимераза (RdRp).[7] Стоп-кодон ORF 24 кДа и стартовый кодон ORF 12 кДа перекрывают друг друга. Хотя многие геномы потексвирусов содержат небольшую ORF, которая находится непосредственно в 5 ’от ORF этого белка оболочки, она не обнаруживается в геноме LVX. Это различие удивительно, поскольку данная ORF была идентифицирована по крайней мере в шести других геномах потексвирусов. Однако при рассмотрении белков, кодируемых LVX и другими потексвирусами, было обнаружено значительное сходство между аминокислотными последовательностями. На 5 ’конце LVX находится усеченная ORF, которая кодирует полипептид, содержащий мотив GDD. Этот мотив также обнаруживается на С-конце белков, кодируемых другими потексвирусами. Исследование белка LVX 24 кДа показывает, что он содержит мотив связывания нуклеозидтрифосфата (GXXGXGKS / T). Этот мотив также обнаружен в белке 25 кДа бессимптомного вируса лилии, а также в белках 25-26 кДа других потексвирусов, таких как вирус X картофеля и вирус S.
LVX также содержит предполагаемую субгеномную промоторную последовательность потексвируса (GGTTAAGTT --- GAA) вверх по течению (5 ’) белка 24 кДа. Эта последовательность также обнаруживается выше кодонов инициации белков оболочки у подобных вирусов, в первую очередь бессимптомного вируса лилии. Исследователи изучили листья лилии, инфицированные LVX, и обнаружили наличие субгеномных РНК. Эти РНК имели длину около 2000 и 850 нуклеотидов, что соответствовало предполагаемым субгеномным промоторам. Исходя из этого, было определено, что они, вероятно, действуют как мессенджеры для белков 24 кДа и 22 кДа (белок оболочки). В последовательности LVX есть область между ORF 12 кДа и ORF 22 кДа (белка оболочки), которая не транслируется. В других последовательностях потексвируса эта область содержит соответствующие ОРС 7 кДа и 11 кДа; однако в этой области последовательности LVX имеется непрерывная рамка считывания. Эта рамка считывания начинается в положении нуклеотида 1236 и заканчивается в положении 1646. Если эта область должна транслироваться, кодируемый белок будет аналогичен упомянутым выше белкам 7 кДа и 11 кДа, оканчиваясь 18 нуклеотидами внутри ORF белка оболочки LVX. Трансляция этой ORF LVX не переводится, потому что в геноме отсутствует кодон инициации.
Тройной генный блок 1 (TGBp1) - многофункциональный белок. Он способствует трансляции вирусных РНК, действуя как РНК. геликаза, разделяющая двухцепочечную РНК для функций RdRp. Более того, он может действовать как подавитель РНК-интерференция, который представляет собой иммунную защиту от накопления вирусных РНК.[8] TGBp1 переносит вирусный геном в соседние растительные клетки напрямую через плазмодесматы, что обеспечивает эффективное размножение за счет обхода барьера клеточной стенки хозяина.[9] TGBp1 также увеличивает пределы исключения плазмодесмального размера, позволяя вирусному геному более легко переходить от одной клетки к другой. Наконец, он подавляет молчание РНК, не позволяя хозяину отключить геном РНК.
TGBp2 и TGBp3 представляют собой мембранно-связывающие белки, участвующие в прикреплении и проникновении. TGBp3 экспрессируется посредством сканирования с утечкой субгеномной мРНК TGBp2. ORF TGBp2 имеет слабый кодон инициации, который иногда пропускается рибосомой при инициации трансляции. В этом случае 40S рибосомальная субъединица продолжает сканирование, пока не встретит инициирующий кодон TGBp3 и не начнет трансляцию.
CP - это белок оболочки LVX, который вместе с TGBp1 и вирусной мРНК образует комплексы рибонуклеопротеидов.
«Нуклеотидная последовательность LVX, по-видимому, является уникальной среди потексвирусов, поскольку в ней явно отсутствует небольшая открытая рамка считывания, 5’ к цистрону белка оболочки, общая для всех других потексвирусов ».[10] Кроме того, в области TGBp3 генома отсутствовал нормальный стартовый кодон.[11] 5'-некодирующая область начинается с GGAAAA, тогда как у других секвенированных потексвирусов, вероятно, все начинаются с GAAAA.[12] Филогенетический анализ кодирующей последовательности LVX показал, что LVX наиболее тесно связан с Умеренный вирус желтого края клубники.[13]

Репликация
LVX не имеет известного вектора, но, скорее всего, распространяется и проникает в клетку посредством механической инокуляции насекомыми. Репликация LVX, как и других ssRNA (+) вирусов, происходит в цитоплазме клеток. Как только вирус попадает в клетку-хозяина, вирус не покрывается оболочкой и высвобождает РНК вирусного генома в цитоплазму. Затем вирусная моноцистронная РНК транслируется в РНК-зависимую РНК-полимеразу, кодируемую 5’-проксимальной ORF. Репликация LVX происходит на вирусных фабриках, которые организованы белком TGBp1.[15] TGBp1 работает над перестройкой актина и эндомембран хозяина и создает сборку спиральных механизмов. Эти спиральные структуры окружены эндомембранами хозяина, которые создают область, в которой вирус может реплицироваться более эффективно.[16]
Репликация ssRNA (+) вируса генерирует вирус dsRNA (+), который затем транскрибируется и реплицируется для создания большего количества геномов LVX mRNA и ssRNA (+) соответственно. Субгеномные промоторы вируса приводят к трансляции sgRNA, что приводит к образованию капсида и белков движения. Они используются для организации структуры вириона и вирусных геномов. Затем можно производить и собирать новые вирусные частицы. С помощью белков тройного генного блока LVX эти завершенные частицы могут преодолевать большие расстояния между плазмодесмами и более короткими межклеточными пространствами, чтобы инфицировать другие клетки-хозяева.[17]
Сопутствующее заболевание
LVX не имеет ассоциированных заболеваний.
Симптомы
Нет никаких известных выдающихся симптомов, которые возникают в естественном диапазоне хозяев LVX, Lilium formosanum; однако Ян (1997) обнаружил, что лилии, инфицированные LVX, растут медленнее и, как правило, меньше, чем неинфицированные экземпляры. Механическая инокуляция других чувствительных видов растений, таких как Тетрагония тетрагониоидес и Chenopodium murale, приводят к местным хлоротическим поражениям.[18]
Тропизм
Лабораторные тесты показали, что LVX способен инфицировать несколько других растений, не считая естественных, хотя и не встречается в природе Lilium formosanum.[19] К другим видам, которые оказались восприимчивыми, относятся:[20]
- Chenopodium capitatum
- Chenopodium murale
- Chenopodium quinoa
- Gomphrena globosa
- Lilium formosanum
- Nicotiana benthamiana
- Никотиана Кливлендии
- Тетрагония тетрагониоидес
LVX способен поражать все ткани лилии. Вирионы были цитопатологически обнаружены во всех частях растений-хозяев и содержат приблизительно 5% нуклеиновой кислоты и 95% белка без содержания липидов.[21] Ожидается, что путь передачи будет механическим заражением насекомыми-переносчиками, поскольку распространение вируса сдерживалось инсектицидами, а не спреями минерального масла.[22]
Рекомендации
- ^ Asjes, C.J. (1991). Борьба с воздушным полевым распространением вируса распада тюльпанов, бессимптомного вируса лилии и вируса лилии X в лилиях с помощью минеральных масел, синтетических пиретроидов и нематицида в Нидерландах Нидерландский журнал пути растений, 97 (3), 129-138.
- ^ Кендалл, А., Макдональд, М., Биан, В., Боулз, Т., Баумгартен, С.С., Ши, Дж.,… Стаббс, Г. (2008). Структура вирусов гибких нитчатых растений. Журнал вирусологии, 82 (19), 9546–9554.
- ^ Кендалл, А., Макдональд, М., Биан, В., Боулз, Т., Баумгартен, С.С., Ши, Дж.,… Стаббс, Г. (2008). Структура вирусов гибких нитчатых растений. Журнал вирусологии, 82 (19), 9546–9554.
- ^ Стоун, О. (1980). Два новых потексвируса из однодольных растений. Acta Hort. 110, 59-63.
- ^ Ян Т. (1997). Цитологические характеристики и обнаружение вирусов Lilium Spp. (Докторская диссертация). Университет Флориды, Гейнсвилл, Флорида.
- ^ Мемелинк, Дж., Ван дер Влугт, C.I.M., Линторст, Х.Дж.М., Деркс, A.F.L.M., Асьес, К.Дж., Бол, Дж.Ф. (1990). Гомологии между геномами карлавируса (бессимптомный вирус лилии) и потексвируса (вирус лилии X) из растений лилии. Журнал общей вирусологии, 71, 917-924.
- ^ Мемелинк, Дж., Ван дер Влугт, C.I.M., Линторст, H.J.M., Деркс, A.F.L.M., Asjes, C.J., Bol, J.F. (1990). Гомологии между геномами карлавируса (бессимптомный вирус лилии) и потексвируса (вирус лилии X) из растений лилии. Журнал общей вирусологии, 71, 917-924.
- ^ Любич-Верчот, Дж. (2005). Новая модель межклеточного транспорта для потексвирусов. Молекулярные взаимодействия растений и микробов, 18 (4), 283-290.
- ^ Любич-Верчот, Дж. (2005). Новая модель межклеточного транспорта для потексвирусов. Молекулярные взаимодействия растений и микробов, 18 (4), 283-290.
- ^ Ян Т. (1997). Цитологические характеристики и обнаружение вирусов Lilium Spp. (Докторская диссертация). Университет Флориды, Гейнсвилл, Флорида.
- ^ Chen, J., Shi, Y.-H., Adams, M.J., Chen, J.-P. (2005). Полная последовательность геномной РНК изолята вируса лилии X (род Potexvirus). Архив вирусологии, 150 (4), 825-832.
- ^ Chen, J., Shi, Y.-H., Adams, M.J., Chen, J.-P. (2005). Полная последовательность геномной РНК изолята вируса лилии X (род Potexvirus). Архив вирусологии, 150 (4), 825-832.
- ^ Chen, J., Shi, Y.-H., Adams, M.J., Chen, J.-P. (2005). Полная последовательность геномной РНК изолята вируса лилии X (род Potexvirus). Архив вирусологии, 150 (4), 825-832.
- ^ Chen, J., Shi, Y.-H., Adams, M.J., Chen, J.-P. (2005). Полная последовательность геномной РНК изолята вируса лилии X (род Potexvirus). Архив вирусологии, 150 (4), 825-832.
- ^ Ян Т. (1997). Цитологические характеристики и обнаружение вирусов Lilium Spp. (Докторская диссертация). Университет Флориды, Гейнсвилл, Флорида.
- ^ Любич-Верчот, Дж. (2005). Новая модель межклеточного транспорта для потексвирусов. Молекулярные взаимодействия растений и микробов, 18 (4), 283-290.
- ^ Ян Т. (1997). Цитологические характеристики и обнаружение вирусов Lilium Spp. (Докторская диссертация). Университет Флориды, Гейнсвилл, Флорида.
- ^ Стоун, О. (1980). Два новых потексвируса из однодольных растений. Acta Hort. 110, 59-63.
- ^ Стоун, О. (1980). Два новых потексвируса из однодольных растений. Acta Hort. 110, 59-63.
- ^ Филлипс, S (1986). "Потексвирус лилии X". Вирусы растений онлайн. Архивировано из оригинал 8 декабря 2015 г.. Получено 7 декабря, 2015.
- ^ Стоун, О. (1980). Два новых потексвируса из однодольных растений. Acta Hort. 110, 59-63.
- ^ Asjes, C.J. (1991). Борьба с воздушным полевым распространением вируса распада тюльпанов, бессимптомного вируса лилии и вируса лилии X в лилиях с помощью минеральных масел, синтетических пиретроидов и нематицидов в Нидерландах. Нидерландский журнал пути растений, 97 (3), 129-138.