Слияние липидного бислоя - Lipid bilayer fusion
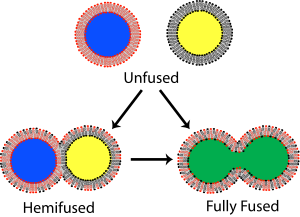
В мембранная биология, слияние это процесс, посредством которого два изначально различных липидные бислои объединить их гидрофобный сердечников, в результате чего получается одна взаимосвязанная конструкция. Если это слияние полностью проходит через обе створки обоих бислоев, водный образуется мостик, и внутреннее содержимое двух структур может смешиваться. Альтернативно, если только одна листочка из каждого бислоя участвует в процессе слияния, говорят, что бислои гемифузированы. При гемифузии липидные составляющие внешнего листка двух бислоев могут смешиваться, но внутренние листочки остаются отдельными. Водное содержимое, заключенное в каждом бислое, также остается разделенным.
Слияние участвует во многих клеточных процессах, особенно в эукариоты так как эукариотическая клетка широко разделена липидными двухслойными мембранами. Экзоцитоз, оплодотворение из яйцо к сперма и транспортировка отходов на лизосома - это лишь некоторые из многих эукариотических процессов, основанных на той или иной форме слияния. Слияние также является важным механизмом транспорта липидов от места их синтеза к мембране, где они необходимы. Даже проникновение болезнетворных микроорганизмов можно регулировать с помощью слияния, так как многие из них имеют двухслойное покрытие. вирусы имеют специальные слитые белки для проникновения в хозяйскую клетку.
Липидный механизм
В процессе слияния есть четыре основных шага, хотя каждый из этих шагов фактически представляет собой сложную последовательность событий.[1] Во-первых, задействованные мембраны должны агрегироваться, приближаясь друг к другу с точностью до нескольких нанометров. Во-вторых, два бислоя должны войти в очень тесный контакт (в пределах нескольких ангстрем). Чтобы достичь этого тесного контакта, две поверхности должны стать, по крайней мере, частично обезвоженными, так как обычно присутствующая связанная поверхностная вода заставляет бислои сильно отталкиваться на таком расстоянии. В-третьих, дестабилизация должна развиваться в одной точке между двумя бислоями, вызывая сильно локализованную перестройку двух бислоев. Наконец, по мере роста этого точечного дефекта компоненты двух бислоев смешиваются и диффундируют прочь от места контакта. В зависимости от того, происходит ли гемифузия или полное слияние, внутреннее содержимое мембран также может смешиваться в этот момент.
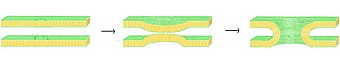
Точные механизмы, стоящие за этой сложной последовательностью событий, до сих пор остаются предметом споров. Чтобы упростить систему и позволить более точное исследование, было проведено множество экспериментов. in vitro с синтетическими липидными пузырьками. Эти исследования показали, что двухвалентные катионы играют решающую роль в процессе слияния, связываясь с отрицательно заряженными липидами, такими как фосфатидилсерин, фосфатидилглицерин и кардиолипин.[2] Одна из ролей этих ионов в процессе синтеза - экранировать отрицательный заряд на поверхности бислоя, уменьшая электростатическое отталкивание и позволяя мембранам приближаться друг к другу. Однако это явно не единственная роль, поскольку существует широко задокументированная разница в способности Mg2+ против Ca2+ вызвать слияние. Хотя Mg2+ вызывает обширную агрегацию, но не вызывает слияния, в то время как Ca2+ вызывает оба.[3] Было высказано предположение, что это несоответствие связано с разницей в степени обезвоживания. Согласно этой теории ионы кальция сильнее связываются с заряженными липидами, но менее прочно с водой. Возникающее в результате вытеснение кальция водой дестабилизирует поверхность раздела липид-вода и способствует тесному межслойному контакту.[4] Недавно предложенная альтернативная гипотеза состоит в том, что связывание кальция вызывает дестабилизирующее латеральное напряжение.[5] Каким бы ни был механизм синтеза, индуцированного кальцием, исходное взаимодействие явно электростатическое, поскольку цвиттерионный липиды не подвержены этому эффекту.[6][7]
В процессе слияния головная группа липидов не только участвует в плотности заряда, но и может влиять на дегидратацию и зарождение дефектов. Эти эффекты не зависят от воздействия ионов. Наличие незаряженного фосфатидилэтаноламина (ПЭ) головной группы увеличивает слияние при включении в бислой фосфатидилхолина. Некоторые объясняют это явление эффектом обезвоживания, аналогичным влиянию кальция.[8] Головная группа из полиэтилена связывает воду менее плотно, чем поликарбонат, и поэтому может облегчить закрытие соединения. Альтернативное объяснение состоит в том, что физическая, а не химическая природа PE может способствовать слиянию. Согласно гипотезе стебля слияния, сильно изогнутый мост должен образоваться между двумя бислоями, чтобы слияние произошло.[9] Поскольку PE имеет небольшую головную группу и легко образует перевернутые мицелла фаз он должен, согласно модели стебля, способствовать формированию этих стеблей.[10] Дополнительным свидетельством, приведенным в пользу этой теории, является тот факт, что было показано, что определенные липидные смеси поддерживают слияние только при повышении температуры выше температуры перехода этих инвертированных фаз.[11][12] Эта тема также остается спорной, и даже если существует криволинейная структура присутствует в процессе сварки, есть дебаты в литературе более, является ли она кубической, гексагональной или более экзотической расширенной фазой.[13]
Белки слияния
Ситуация еще более усложняется при рассмотрении слияния in vivo поскольку биологическое слияние почти всегда регулируется действием мембранные белки. Первыми из этих белков, которые были изучены, были вирусные слитые белки, которые позволяют вирус для вставки своего генетического материала в клетку-хозяина (вирусы с оболочкой - это вирусы, окруженные липидным бислоем; у некоторых других есть только белковая оболочка). В целом, существует два класса слитых белков вирусов: кислые и независимые от pH.[1] pH независимые слитые белки могут функционировать в нейтральных условиях и сливаться с плазматическая мембрана, позволяя вирусам проникать в клетку. Вирусы, использующие эту схему, включали ВИЧ, корь и герпес. Кислотные гибридные белки, такие как грипп активируются только при низком pH кислых эндосом и должны быть эндоцитозированный чтобы попасть в камеру.
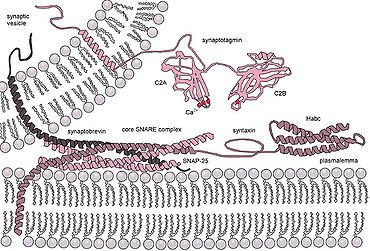
Эукариотический клетки используют совершенно разные классы гибридных белков, наиболее изученными из которых являются SNAREs. Белки SNARE используются для управления всеми везикулярный внутриклеточный трафик. Несмотря на годы исследований, многое еще неизвестно о функции этого класса белков. Фактически, до сих пор ведутся активные дебаты относительно того, связаны ли SNAREs с ранней стыковкой или участвуют позже в процессе слияния, облегчая гемифузию.[15] Даже после того, как будет освещена роль SNARE или других специфических белков, единое понимание слитых белков маловероятно, поскольку существует огромное разнообразие структур и функций внутри этих классов, и очень мало тем сохраняется.[16]
Fusion в лабораторной практике
При изучении молекулярной и клеточной биологии часто бывает желательно искусственно вызвать слияние. Хотя это может быть достигнуто с добавлением кальция, как обсуждалось ранее, эта процедура часто невозможна, потому что кальций регулирует многие другие биохимические процессы, и его добавление было бы серьезным затруднением. Кроме того, как уже упоминалось, кальций вызывает массивную агрегацию, а также слияние. Добавление полиэтиленгликоль (ПЭГ) вызывает слияние без значительной агрегации или биохимического нарушения. Эта процедура сейчас широко используется, например, путем сплавления В-клетки с миелома клетки.[17] Результирующий "гибридома »Из этой комбинации выражает желаемое антитело как определено вовлеченными В-клетками, но увековечивается из-за компонента миеломы. Механизм слияния PEG окончательно не определен, но некоторые исследователи считают, что PEG, связывая большое количество молекул воды, эффективно снижает химическая активность воды и, таким образом, обезвоживает головные группы липидов.[18] Слияние также может быть искусственно вызвано электропорация в процессе, известном как электросварка. Считается, что это явление связано с энергетически активные края формируется во время электропорации, которая может действовать как локальная точка дефекта для зарождения стебля между двумя бислоями.[19]
Анализы для измерения слияния мембран
Существует два уровня слияния: смешивание мембранных липидов и смешивание содержимого. Анализы слияния мембран сообщают либо о смешивании липидов мембран, либо о смешивании водного содержимого слитых объектов.
Анализы для измерения смешивания липидов
Анализы, оценивающие смешение липидов, используют зависящие от концентрации эффекты, такие как безызлучательный перенос энергии, тушение флуоресценции и образование эксимера пирена.
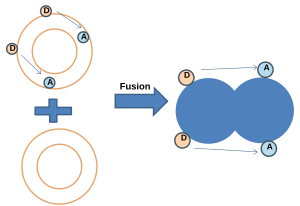

- Передача энергии NBD-родамином:[20] В этом методе мембрана, меченная как NBD (донор), так и родамином (акцептор), объединяется с немеченой мембраной. Когда NBD и родамин находятся на определенном расстоянии, происходит резонансная передача энергии Фёрстера (FRET). После слияния резонансная передача энергии (FRET) уменьшается при увеличении среднего расстояния между зондами, в то время как флуоресценция NBD увеличивается.
- Образование эксимерного пирена: Пирен мономер и эксимер длины волн излучения различны. Длина волны излучения мономера составляет около 400 нм, а эксимера - около 470 нм. В этом методе мембрана, меченная пиреном, сочетается с немеченой мембраной. Пирен самоассоцируется в мембране, а затем возбужденный пирен возбуждает другой пирен. До синтеза большая часть излучения - это эксимерное излучение. После слияния расстояние между зондами увеличивается, а коэффициент эксимерной эмиссии уменьшается.[нужна цитата ]
- Октадецил родамин B Самозатухающий:[21] Этот анализ основан на самогашении октадецилродамина B. Самотушение октадецилродамина B происходит, когда зонд включается в липиды мембран в концентрациях 1–10 мольных процентов.[22] потому что димеры родамина подавляют флуоресценцию. В этом методе мембрана, помеченная родамином, сочетается с немеченой мембраной. Слияние с немечеными мембранами приводит к разбавлению зонда, которое сопровождается увеличением флуоресценции.[23][24] Основная проблема этого теста - спонтанный перенос.
Анализы для измерения смешивания содержимого
Смешивание водного содержимого с пузырьками в результате лизиса, слияния или физиологической проницаемости может быть обнаружено флуорометрически с использованием растворимых индикаторов с низким молекулярным весом.


- Анализы тушения флуоресценции с ANTS / DPX:[25][26] ANTS - полианионный флуорофор, а DPX - катионный тушитель. Анализ основан на их столкновительном тушении. В отдельные популяции везикул загружают ANTS или DPX соответственно. Когда происходит смешивание содержимого, ANTS и DPX сталкиваются, и флуоресценция ANTS, контролируемая на 530 нм, с возбуждением на 360 нм гаснет. Этот метод выполняется при кислом pH и высокой концентрации.
- Анализы усиления флуоресценции с Tb3+/ DPA:[27][28] Этот метод основан на том, что хелат Tb3+/ DPA в 10 000 раз более флуоресцентный, чем Tb3+ один. В ТБ3+/ Анализ DPA, отдельные популяции везикул загружены TbCl3 или DPA. Образование туберкулеза3+/ DPA-хелат может использоваться для обозначения слияния везикул. Этот метод хорош для мембран, не содержащих белка.[нужна цитата ]
- Анализ одиночной молекулы ДНК.[29] Шпилька ДНК, состоящая из стержня из 5 пар оснований и политимидиновой петли, меченная донором (Cy3) и акцептором (Cy5) на концах стержня, была инкапсулирована в везикулу v-SNARE. Мы отдельно инкапсулировали несколько немеченых цепей полиаденозиновой ДНК в везикуле t-SNARE. Если две везикулы, оба диаметром ~ 100 нм, состыковываются и между ними образуется достаточно большая пора слияния, две молекулы ДНК должны гибридизоваться, открывая область стержня шпильки и переключая эффективность резонансной передачи энергии Фёрстера (FRET) ( E) между Cy3 и Cy5 от высокого до низкого значения.
Смотрите также
Рекомендации
- ^ а б Йигл, П. Л. (1993). Мембраны клеток (2-е изд.). Сан-Диего: Academic Press.[страница нужна ]
- ^ Папахаджопулос, Деметриос; Нир, Шломо; Düzgünes, Nejat (1990). «Молекулярные механизмы индуцированного кальцием слияния мембран». Журнал биоэнергетики и биомембран. 22 (2): 157–79. Дои:10.1007 / BF00762944. PMID 2139437. S2CID 1465571.
- ^ Левентис, Рания; Ганье, Жаннин; Фуллер, Нола; Rand, R .; Сильвиус, Дж. (1986). «Двухвалентное катион-индуцированное слияние и боковая сегрегация липидов в пузырьках фосфатидилхолин-фосфатидная кислота». Биохимия. 25 (22): 6978–87. Дои:10.1021 / bi00370a600. PMID 3801406.
- ^ Вильшут, Ян; Duezguenes, Nejat; Папахаджопулос, Деметриос (1981). «Специфичность кальция / магния в слиянии мембран: кинетика агрегации и слияния везикул фосфатидилсерина и роль изгиба бислоя». Биохимия. 20 (11): 3126–33. Дои:10.1021 / bi00514a022. PMID 7248275.
- ^ Чантурия, А; Scaria, P; Вудл, MC (2000). "Роль бокового натяжения мембраны в индуцированном кальцием слиянии мембран". Журнал мембранной биологии. 176 (1): 67–75. Дои:10.1007 / s00232001076. PMID 10882429. S2CID 2209769.
- ^ Паннуццо, Мартина; Джонг, Де; Djurre, H .; Раудино, Антонио; Марринк Сиверт, Дж. (2014). «Моделирование слияния полиэтиленгликоля и кальций-опосредованной мембраны» (PDF). J. Chem. Phys. 140 (12): 124905. Bibcode:2014ЖЧФ.140л4905П. Дои:10.1063/1.4869176. PMID 24697479.
- ^ Papahadjopoulos, D .; Poste, G .; Schaeffer, B.E .; Вейл, W.J. (1974). «Слияние мембран и молекулярная сегрегация в фосфолипидных везикулах». Biochimica et Biophysica Acta (BBA) - Биомембраны. 352 (1): 10–28. Дои:10.1016/0005-2736(74)90175-8. PMID 4859411.
- ^ Дюзгюнес, Неджат; Вильшут, Ян; Фрейли, Роберт; Папахаджопулос, Деметриос (1981). «Исследования механизма слияния мембран. Роль состава головной группы в индуцированном кальцием и магнием слиянии смешанных фосфолипидных везикул». Biochimica et Biophysica Acta (BBA) - Биомембраны. 642 (1): 182–95. Дои:10.1016/0005-2736(81)90148-6. PMID 7225377.
- ^ Маркин В.С.; Козлов, ММ; Боровягин, ВЛ (1984). «К теории слияния мембран. Стеблевой механизм» (PDF). Общая физиология и биофизика. 3 (5): 361–77. PMID 6510702.
- ^ Черномордик, Леонид В .; Козлов, Михаил М. (2003). «Взаимодействие белков и липидов при слиянии и делении биологических мембран». Ежегодный обзор биохимии. 72: 175–207. Дои:10.1146 / annurev.biochem.72.121801.161504. PMID 14527322.
- ^ Nir, S .; Bentz, J .; Wilschut, J .; Дузгунес, Н. (1983). «Агрегация и слияние фосфолипидных везикул». Прогресс в науке о поверхности. 13 (1): 1–124. Bibcode:1983ПрСС ... 13 .... 1Н. Дои:10.1016/0079-6816(83)90010-2.
- ^ Элленс, Харма; Бенц, Джо; Szoka, Фрэнсис К. (1986). «Слияние липосом, содержащих фосфатидилэтаноламин, и механизм фазового перехода L.alpha.-HII». Биохимия. 25 (14): 4141–7. Дои:10.1021 / bi00362a023. PMID 3741846.
- ^ Holopainen, Juha M .; Lehtonen, Jukka Y.A .; Киннунен, Пааво К.Дж. (1999). «Доказательства расширенной конформации фосфолипидов при слиянии мембран и гемифузии». Биофизический журнал. 76 (4): 2111–20. Bibcode:1999BpJ .... 76.2111H. Дои:10.1016 / S0006-3495 (99) 77367-4. ЧВК 1300184. PMID 10096906.
- ^ Георгиев, Данко Д .; Глейзбрук, Джеймс Ф. (2007). «Субнейронная обработка информации уединенными волнами и случайными процессами». В Лышевском, Сергей Эдуард (ред.). Справочник по нано- и молекулярной электронике. Серия нано- и микротехники. CRC Press. С. 17–1–17–41. Дои:10.1201 / 9781420008142.ch17 (неактивно 01.09.2020). ISBN 978-0-8493-8528-5.CS1 maint: DOI неактивен по состоянию на сентябрь 2020 г. (связь)
- ^ Chen, Yu A .; Шеллер, Ричард Х. (2001). «SNARE-опосредованное слияние мембран». Обзоры природы Молекулярная клеточная биология. 2 (2): 98–106. Дои:10.1038/35052017. PMID 11252968. S2CID 205012830.
- ^ Уайт, Дж. М. (1990). «Белки слияния вирусных и клеточных мембран». Ежегодный обзор физиологии. 52: 675–97. Дои:10.1146 / annurev.ph.52.030190.003331. PMID 2184772.
- ^ Köhler, G .; Мильштейн, К. (1975). «Непрерывные культуры слитых клеток, секретирующих антитела с заранее определенной специфичностью». Природа. 256 (5517): 495–7. Bibcode:1975Натура.256..495K. Дои:10.1038 / 256495a0. PMID 1172191. S2CID 4161444.
- ^ Ленц, Барри Р. (1994). «Полимер-индуцированное слияние мембран: потенциальный механизм и связь с событиями слияния клеток». Химия и физика липидов. 73 (1–2): 91–106. Дои:10.1016/0009-3084(94)90176-7. PMID 8001186.
- ^ Jordan, C.A .; Neumann, E .; Сауэрс, А. Э., ред. (1989). Электропорация и электрослияние в клеточной биологии. Springer. ISBN 978-0-306-43043-5.[страница нужна ]
- ^ Ударил, Дуглас К .; Хукстра, Дик; Пагано, Ричард Э. (1981). «Использование резонансной передачи энергии для контроля слияния мембран». Биохимия. 20 (14): 4093–9. Дои:10.1021 / bi00517a023. PMID 7284312.
- ^ Хукстра, Дик; Де Бур, Крошечный; Клаппе, Карин; Вильшут, Ян (1984). «Флуоресцентный метод измерения кинетики слияния биологических мембран». Биохимия. 23 (24): 5675–81. Дои:10.1021 / bi00319a002. PMID 6098295.
- ^ Макдональд, Рубин I (1990). «Характеристики самотушения флуоресценции липид-конъюгированного родамина в мембранах». Журнал биологической химии. 265 (23): 13533–9. PMID 2380172.
- ^ Rubin, R.J .; Чен, Ю. (1990). «Диффузия и перераспределение липидоподобных молекул между мембранами в системах слияния вирус-клетка и клетка-клетка». Биофизический журнал. 58 (5): 1157–67. Bibcode:1990БпДж .... 58.1157R. Дои:10.1016 / S0006-3495 (90) 82457-7. ЧВК 1281061. PMID 2291940.
- ^ Chen, Y.D .; Rubin, R.J .; Сабо, А. (1993). «Кинетика ослабления флуоресценции одноклеточных слитых комплексов». Биофизический журнал. 65 (1): 325–33. Bibcode:1993BpJ .... 65..325C. Дои:10.1016 / S0006-3495 (93) 81076-2. ЧВК 1225727. PMID 8369440.
- ^ Смолярский, Моше; Тейтельбаум, Двора; Села, Майкл; Гитлер, Карлос (1977). «Простой флуоресцентный метод определения опосредованного комплементом иммунного лизиса липосом». Журнал иммунологических методов. 15 (3): 255–65. Дои:10.1016/0022-1759(77)90063-1. PMID 323363.
- ^ Ellens, H; Бенц, Дж; Сока, ФК (1985). «H + - и Ca2 + -индуцированное слияние и дестабилизация липосом». Биохимия. 24 (13): 3099–106. Дои:10.1021 / bi00334a005. PMID 4027232.
- ^ Вильшут, Ян; Папахаджопулос, Деметриос (1979). «Ca2 + -индуцированное слияние фосфолипидных везикул, отслеживаемое смешиванием водного содержимого». Природа. 281 (5733): 690–2. Bibcode:1979Натура.281..690Вт. Дои:10.1038 / 281690a0. PMID 551288. S2CID 4353081.
- ^ Вильшут, Ян; Дузгунес, Неджат; Фрейли, Роберт; Папахаджопулос, Деметриос (1980). «Исследования механизма слияния мембран: кинетика слияния фосфатидилсериновых везикул, индуцированного ионами кальция, с последующим новым анализом смешивания содержимого водных везикул». Биохимия. 19 (26): 6011–21. Дои:10.1021 / bi00567a011. PMID 7470445.
- ^ Дяо, Цзяцзе; Су, Цзэнлю; Ишицука, Юдзи; Лу, Бин; Ли, Кён Сок; Лай, Инь; Шин, Ён-Кюн; Ха, Таэкджип (2010). «Анализ смешивания содержимого одиночных везикул для слияния мембран, опосредованного SNARE». Nature Communications. 1 (5): 1–6. Bibcode:2010 НатКо ... 1E..54D. Дои:10.1038 / ncomms1054. ЧВК 3518844. PMID 20975723.