Мезенозавр - Mesenosaurus
| Мезенозавр | |
|---|---|
 | |
| Мезенозавр romeri | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Семья: | †Varanopidae |
| Подсемейство: | †Mycterosaurinae |
| Род: | †Мезенозавр Ефремов, 1938 г. |
| Типовой вид | |
| †Мезенозавр romeri, Ефремов, 1938 г. | |
| Разновидность | |
| |
Мезенозавр вымерший род не-млекопитающее синапсиды. Он принадлежит семье Varanopseidae (также известный как Varanopidae). Это семейство пейклозавров (парафилетическая группа нетерапсидных синапсидов) имеет самую длинную летопись окаменелостей и самое широкое географическое распространение.
Этот род включает два вида: типовой вид Мезенозавр romeri из средней перми Река Мезень Бассейн северной Россия, и Мезенозавр ефремови с ранней перми Ричардс Спур местонахождение (Оклахома, Соединенные Штаты ).[1] М. РомериСтратиграфический диапазон - от среднего до позднего гваделупского, в то время как М. ефремовиСтратиграфический диапазон страны - Приуралье.[1]
Этимология
Известный русский палеонтолог, Иван Ефремов, установленный Мезенозавр как род, и назвал его по корням с (дословный перевод «ящерица из Мезени»).
Мезенозавр ефремови назван в честь Ивана Ефремова, воздвигнувшего род.[1]
Описание
Мезенозавр представляют собой синапсиды варанопсеидов небольшого размера. Для них характерны преимущественно черепные черты.[2] Многие посткраниальные особенности этого рода не были полностью проанализированы из-за отсутствия свидетельств окаменелостей.
Череп
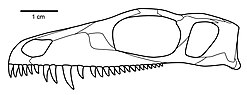
Черты черепа, характеризующие Мезенозавр находятся:
- Стройная предчелюстная кость образует узкую прямоугольную морду (вид сверху и снизу)
- Дорсальный отросток предчелюстной кости длинный, образует переднюю половину дорсального края наружного яруса.
- Расширенная повязка, доходящая почти до кончика морды
- Небный отросток предчелюстной кости длинный с длинным срединным швом
- Хорошо развитое углубление на боковой поверхности носа, простирающееся кзади от нариальной границы почти до переднего конца префронтальной
- Небольшая боковая припухлость верхней челюсти на уровне клыковидного зуба
- Короткий задний отросток верхней челюсти не достигает уровня заглазничной перемычки.
- Первый предчелюстной зуб меньше второго и третьего. (На предчелюстной кости не более пяти зубов. Первый зуб меньше второго и третьего, но больше двух последних. Эти пять зубов расположены близко друг к другу, сильно изогнуты, заострены и имеют острый режущий край. по дистальной половине заднего края.)
- Один средний зубной ряд сошника
- Заглазничная щечная область черепа необычайно широкая и низкая, с почти вертикальным задним краем.
- Задний край поперечного фланца крыловидный слегка наклонен кпереди от базального сочленения.
- Стремени тонкие, короткие, стержневидные, со скромно развитой подошвой и расширенными дистально квадратный процесс. (Ступени Мезенозавр более похож на таковые в ранних терапсидах Перми.)
- Очень заметный узловой орнамент присутствует на орбитальных краях префронтальной, заглазничной и скуловой областей. (Эта особенность ярко выражена в хорошо сохранившихся черепах Мезенозавр, настолько, что Ефремов первоначально считал это аутапоморфией Мезенозавр. Однако тщательное изучение и анализ показали, что эта особенность присутствует и у других варанопсеидов.)[2]
Открытие
Мезенозавр romeri
Бассейн реки Мезень расположен на севере России и имеет обширные выходы среднепермских отложений по берегам полноводных рек реки Мезень. В этих отложениях образовалось множество скелетных останков различных амниот, но, что наиболее важно, частичный череп «небольшого синапсида сродства варанопсеидов», Мезенозавр romeri.[2] М. Ромери был открыт в 1938 году Иваном Ефремовым и стал первым видом мезенозавра из-за отсутствия у него черепных сходств с другими. Кроме того, это была первая описанная рептилия из российских регионов, которую считали «хорошим пеликозавром», поскольку у нее были зубы верхней челюсти, которые соответствовали другим известным пеликозаврам (тонкие, загнутые, увеличенные резцы, единственный клыковидный зуб). Ромер и Прайс предположили, что М. Ромери принадлежали к Varanopseidae.[3] Эта гипотеза была подтверждена в 2001 году на основании следующих синапоморфий:
- Дорсовентральное расширенное височное окно занимает большую часть высоты височной области, что приводит к узкой субвисочной полосе
- Краевой зубной ряд состоит из сильно загнутых, сжатых с боков остроконечных передних и задних режущих кромок зубов, ограниченных дистальной половиной зуба.
- Хорошо развитая предчелюстная субнариальная полка, внешняя поверхность которой в поперечном сечении широко закруглена.
- Передний срединный отросток теменной кости заходит в надглазничную область стола черепа.
- Небольшая постфронтальная кайма медиально узкая, задний отросток лобной
- Табличные сморщенные размеры, которые становятся небольшими узкими элементами, которые контактируют с медиальным краем заднебокового крыла теменной кости.
- Отсутствие медиального, затылочного фланца заднего края чешуйчатой кости, закрывающего задний край квадратной кости
- Парасфеноидная пластинка широкая, базисфеноидальные туберы крыловидные, далеко отходят латеральнее и кзади от основания культиформного отростка.
- Выраженный узловой орнамент присутствует на краях глазницы префронтальной, заглазничной и скуловой областей.[2]
Мезенозавр ефремови
Второй вид этого рода - Mesenosaurus efremovi. Его почти полный череп и нижняя челюсть были обнаружены в районе Ричардс-Спур в ряду заполненных карстовых трещин в ордовикском известняке Арбакл в Оклахоме, который является одним из самых многочисленных участков для раннего изучения. Пермский период окаменелости четвероногих.[4]
С точки зрения классификации М. ефремови, он имеет общие черты черепа с миктерозавринами (группа на основе стебля, которая включает Mycterosaurus longiceps и все варанопсеиды более тесно связаны с ним, чем с Варанодон агилис), такие как «исключение слезной жидкости из наружного носа и переднезадней широкой дорсальной пластинки верхней челюсти, которая лежит в основе носа и контактирует с префронтальной костью». Однако, М. ефремови делится другими функциями с М. Ромери из России. Некоторые из этих общих черт включают относительный размер и форму височного отверстия, латеральный отек верхней челюсти в клыковидной области и пять предчелюстных зубов (не сообщается у других миктерозавринов).[1]
Хотя М. ефремови и М. Ромери имеют много отличительных черт, между этими образцами есть четыре основных морфологических различия, которые определяют таксономические различия на уровне видов (различия недостаточны для различения выше уровня видов):
- Наличие коротких дорсальных предчелюстных отростков (которые не доходят ни до заднего лицевого края, ни до заднего отделения предчелюстных костей от носа)
- Более обширная кзади верхняя челюсть
- Меньшее количество позиций зубов на верхней челюсти
- Контакт между заглазничной и надвисочной костями
М. ефремови также больше, чем самый большой известный образец M. romeri. [1]
Классификация
Ниже приводится кладограмма изменено на основе анализа Бенсона, после исключения Базеранодон:
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеосреда
Между этими двумя видами существует разрыв в 20 млн лет, что превышает временной диапазон большинства вымерших четвероногих. Хотя этот пробел вызывает сомнения, радиоизотопное датирование образований, восстановленное для Richards Spur, использованное для идентификации этого 20-миллионного пробела, является надежным методом, который использовался для идентификации других больших пробелов / длинных временных диапазонов.[6] Этот разрыв является значительным, поскольку он указывает на эволюционный застой (сохраняющийся в различных средах Пангеи, а также на круговорот фауны на протяжении всего пермского периода). Разрыв Олсона ). Этот эволюционный застой может быть объяснен сохранением занятия ниши во всех временных и географических пределах. Требуются дальнейшие исследования посткраниальных особенностей, чтобы определить, верна ли эта гипотеза, поскольку нам нужно будет наблюдать аналогичную степень застоя во всем скелете.[1]
Палеобиология
Мезенозавр - маленькое подвижное существо, способное лазить по камням / деревьям.[7] Их небольшой размер позволил им занять и процветать в нишах в качестве мелких фауноядных животных, в то время как терапсиды доминировали в большинстве наземных сред. Только в конце перми, когда маленький диапсиды появившийся[8] и обеспечили конкуренцию, что привело к снижению Мезенозавр. Мезенозавр представляет гильдию очень ловких подчиненных хищников[1][9] в их сообществах из-за их больших, тонких, изогнутых зубов, которые могли нанести серьезные раны при прокалывании своей добычи.[10]
У них были согнутые и сплющенные ногти, что наводило на мысль, что они были землекопами. Неизвестно, были ли их копающие когти приспособлены для рытья нор или копания исключительно для еды.[10] Многие варанопсеиды были древесными, однако хорошо развитый олекранон (костный выступ локтя) мезенозавра указывает на наличие трехглавой мышцы и мышцы анонуса,[11][12][13] оба из них обеспечат мощное разгибание предплечья. Эта сила разгибания предплечья в сочетании с небольшими / средними размерами тела поддерживает идею роющего образа жизни.[10] Также предлагается, чтобы факультативное двуногие произошел у мезенозавра. Это основано на наличии смещения центра массы тела назад (тонкие туловища, удлиненные задние конечности и короткие передние конечности), что необходимо для факультативного двуногого мышления.[10]
Смотрите также
Рекомендации
- ^ а б c d е ж грамм Сиги Махо; Брайан М. Джи; Роберт Р. Рейс (2019). «Новый синапсид варанопидов из ранней перми Оклахомы и эволюционный застой в этой кладе». Королевское общество открытой науки. 6 (10): Идентификатор статьи 191297. Дои:10.1098 / RSOS.191297. ЧВК 6837192. PMID 31824730.
- ^ а б c d Рейс, Роберт Р. (2001). Череп Мезенозавр romeri, небольшой варанопсеид (Synapsida: Eupelycosauria) из верхней перми бассейна реки Мезень, север России. OCLC 633882190.
- ^ ROMER, A. S .; PRICE, L. W. (1940), "Обзор пеликозавров", 28: Обзор пеликозаврии, Специальные статьи Геологического общества Америки, 28, Геологическое общество Америки, стр. 1–534, Дои:10.1130 / spe28-p1
- ^ MacDougall, Mark J .; Табор, Нил Дж .; Вудхед, Джон; Дауст, Эндрю Р .; Рейс, Роберт Р. (июнь 2017 г.). «Уникальная природоохранная среда раннепермских (цисуральских) пещерных отложений, содержащих окаменелости, в местности Ричардс Спур, Оклахома». Палеогеография, палеоклиматология, палеоэкология. 475: 1–11. Дои:10.1016 / j.palaeo.2017.02.019. ISSN 0031-0182.
- ^ Бенсон, Р.Дж. (2012). «Взаимоотношения базальных синапсидов: черепные и посткраниальные морфологические перегородки предполагают разную топологию». Журнал систематической палеонтологии. 10 (4): 601–624. Дои:10.1080/14772019.2011.631042.
- ^ Вудхед, Джон; Рейс, Роберт; Фокс, Дэвид; Дрисдейл, Рассел; Хеллстрем, Джон; Маас, Роланд; Ченг, Хай; Эдвардс, Р. Лоуренс (май 2010 г.). «Климатические записи Speleothem из глубоких времен? Изучение потенциала на примере пермского периода». Геология. 38 (5): 455–458. Дои:10.1130 / g30354.1. ISSN 1943-2682.
- ^ Шпиндлер, Фредерик; Вернебург, Ральф; Schneider, Joerg W .; Luthardt, Людвиг; Аннакер, Фолькер; Рёсслер, Ронни (15 марта 2018 г.). «Первые древесные« пеликозавры »(Synapsida: Varanopidae) из ранней перми Chemnitz Fossil Lagerstätte, SE Германия, с обзором филогении варанопидов». PalZ. 92 (2): 315–364. Дои:10.1007 / s12542-018-0405-9. ISSN 0031-0220.
- ^ Андерсон, Джейсон С .; Рейс, Роберт Р. (2004-03-25). «Pyozia mesenensis, новый малый варанопид (Synapsida, Eupelycosauria) из России: разнообразие« пеликозавров »в средней перми». Журнал палеонтологии позвоночных. 24 (1): 173–179. Дои:10.1671/1940-13. ISSN 0272-4634.
- ^ Модесто, Кристиан А. Сидор, Брюс, Шон (2001-12-01). «Второй череп варанопсеида из верхней перми в Южной Африке: значение для позднепермской эволюции« пеликозавра »». Lethaia. 34 (4): 249–259. Дои:10.1080/002411601753292971. ISSN 0024-1164.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ а б c d Шпиндлер, Фредерик; Вернебург, Ральф; Шнайдер, Йорг В. (19 января 2019 г.). «Новый мезенозаврин из нижней перми Германии и посткрании мезенозавра: значение для ранней сравнительной остеологии амниот». PalZ. 93 (2): 303–344. Дои:10.1007 / s12542-018-0439-z. ISSN 0031-0220.
- ^ Haines, R.W. (1950). «Сгибатели предплечья и кисти у ящериц и млекопитающих». Журнал анатомии. 84 (Pt 1) (Pt 1): 13–29. ЧВК 1273351. PMID 17105089.
- ^ Haines, R.W. (1939). «Ревизия мышц-разгибателей предплечья у четвероногих». Журнал анатомии. 73 (2) (Pt 1): 211–233. ЧВК 1273351. PMID 17105089.
- ^ Сэвидж, Джей М .; Ромер, Альфред Шервуд (1957-07-15). «Остеология рептилий». Копея. 1957 (2): 162. Дои:10.2307/1439431. ISSN 0045-8511. JSTOR 1439431.
внешняя ссылка
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||



