Оливокохлеарная система - Olivocochlear system
В оливокохлеарная система является составной частью слуховая система связан с нисходящим контролем над улитка. Его нервные волокна, оливокохлеарный пучок (ОКБ), образуют часть вестибулокохлеарный нерв (VIII черепной нерв, также известный как слухово-вестибулярный нерв), и выступает из высший оливарный комплекс в мозговой ствол (мосты ) в улитку.
Анатомия оливокохлеарной системы
Тела происхождения клеток

Оливокохлеарный пучок (OCB) берет начало в высший оливарный комплекс в стволе мозга. Вестибулокохлеарный анастомоз несет эфферент аксоны в улитку, где они иннервируют орган Корти (OC). OCB содержит волокна, выходящие как на ипсилатеральный и контралатеральный cochleae, что побудило начальное разделение на скрещенные (COCB) и неперекрещенные (UCOCB) системы.[1] Однако в последнее время деление ОКВ основано на месте происхождения клеточных тел в стволе мозга относительно медиальной верхней оливы (MSO). Медиовентральная периоливная область (MVPO), также известная как вентральное ядро трапециевидного тела, диффузная область нейронов, расположенных медиальнее MSO, дает начало медиальной оливокохлеарной системе (MOCS). Латеральная верхняя олива (LSO), отдельное ядро нейронов, расположенных латеральнее MSO, дает начало латеральной оливокохлеарной системе (LOCS).[2][3] Нейроны MOCS представляют собой большие мультиполярные клетки, в то время как LOCS классически определяется как состоящий из небольших сферических клеток. Это разделение считается более значимым с точки зрения физиологии ОКБ.[4] В дополнение к этим классически определенным оливокохлеарным нейронам, успехи в методах отслеживания трактов помогли выявить третий класс оливокохлеарных нейронов, называемых нейронами оболочки, которые окружают LSO.[5] Таким образом, тела клеток класса LOCS в LSO называются внутренними нейронами LOCS, а те, что окружают LSO, называются нейронами оболочки или внешними нейронами LOCS. Оболочечные нейроны, как правило, большие и морфологически очень похожи на нейроны MOCS.
Оливокохлеарные волокна
LOCS (происходящий как из внутренних нейронов, так и из нейронов оболочки) содержит немиелинизированные волокна, которые синапсируют с дендритами I типа. спиральный узел клетки, выступающие внутрь волосковые клетки. Хотя внутренние нейроны LOCS имеют тенденцию быть небольшими (от ~ 10 до 15 мкм в диаметре), а нейроны оболочки OC больше (~ 25 мкм в диаметре), именно внутренние нейроны OC обладают более крупными аксонами (0,77 мкм по сравнению с Диаметр 0,37 мкм для нейронов оболочки). Напротив, MOCS содержит миелинизированные нервные волокна, которые напрямую иннервируют внешние волосковые клетки.[6] Хотя и LOCS, и MOCS содержат пересекающиеся (контралатеральные) и неперекрещенные (ипсилатеральные) волокна, у большинства видов млекопитающих большинство волокон LOCS проецируются в ипсилатеральную улитку, в то время как большинство волокон MOCS проецируются в контралатеральную улитку.[2][7] Доля волокон в MOCS и LOCS также варьируется между видами, но в большинстве случаев волокна LOCS более многочисленны.[8][9][10] У людей насчитывается примерно 1000 волокон LOCS и 360 волокон MOCS.[11][12] однако цифры варьируются от человека к человеку. MOCS вызывает частотно-зависимую иннервацию улитки, поскольку волокна MOC оканчиваются на внешних волосковых клетках в том месте в улитке, которое определяется на основе характерной частоты волокон, и, таким образом, тонотопически организованы таким же образом, как и первичные волосковые клетки. афферентные нейроны.[6][13] Волокна LOCS также располагаются в виде тонотопный мода.[14] Однако неизвестно, совпадают ли характеристические частоты волокон LOCS с характеристическими частотами первичных афферентных нейронов, поскольку попытки избирательно стимулировать волокна LOCS были в значительной степени безуспешными.[15] Внутренние аксоны, происходящие из LOCS, перемещаются только примерно на 1 мкм в кортиевом органе, обычно спиралевидно апикально. Они испускают небольшой пучок синаптических бутонов, компактный по своей протяженности, часто включающий менее 10 IHC. Для сравнения, нейроны оболочки спиралевидны как апикально, так и базально, и могут покрывать большие территории внутри кортиевого органа. Аксоны оболочки часто покрывают 1-2 октавы тонотопической длины.[16] Однако их конечная беседка довольно редка.
Физиология оливокохлеарной системы
Нейрофизиология
Вся известная в настоящее время активность оливокохлеарной системы осуществляется через рецепторный комплекс нейромедиаторов никотинового класса, который связан с калиевым каналом, активируемым кальцием. Вместе эти системы создают необычный синаптический ответ на стимуляцию со стороны мозга. Оливокохлеарные синаптические терминалы содержат различные нейротрансмиттеры и нейроактивные пептиды. Основным нейротрансмиттером, используемым в оливокохлеарной системе, является ацетилхолин (АХ), несмотря на то что гамма-аминомасляная кислота (ГАМК) также локализован в терминалах. Высвобождение АХ из оливокохлеарных окончаний активирует эволюционно древний холинэргический рецептор комплекс, состоящий из никотиновый альфа9[17] и альфа10 субъединицы.[18] Эти субъединицы создают управляемый лигандом ионный канал, который особенно проницаем для кальций и моновалентный катионы[19] то сотовый ответ внешнего волосковые клетки в АЧ активация есть гиперполяризующий, а не ожидаемый деполяризующий ответ. Это происходит из-за быстрой активации связанного калий канал. Этот канал, апамин чувствительный, малая проводимость Калиевый канал SK2, активируется кальцием, который, вероятно, попадает в цитоплазма через кальций-индуцированное высвобождение кальция из запасов кальция в подсинаптический цистерны в ответ на поступление кальция из никотинового комплекса.[20] Однако не исключено, что некоторое количество кальция, поступающего через никотиновый канал alpha9alpha10, также может напрямую активировать SK канал. Электрофизиологические ответы, зарегистрированные от наружных волосковых клеток после стимуляции ACh, поэтому показывают небольшой внутренний ток (переносимый в основном кальцием через рецептор ацетилхолина ), за которым сразу следует большой выходящий ток, ток калия, который гиперполяризует внешнюю волосковую клетку.
Когда оливокохлеарный пучок рассекают хирургическим путем до появления слуха, слуховая чувствительность нарушается.[21] Однако при генетическом удалении генов альфа9 или альфа10 такие эффекты не наблюдаются. Это может быть связано с различной природой поражений - хирургическое поражение приводит к полной потере всей оливокохлеарной иннервации волосковых клеток, тогда как генетические манипуляции приводят к гораздо более избирательной функциональной потере - только целевого гена. Любые оставшиеся нейроактивные вещества, которые могут высвобождаться неповрежденными синаптическими окончаниями, могут активировать волосковые клетки. Действительно, после генетического удаления одного из нейроактивных пептидов, присутствующих в терминалах LOCS,[22] Наблюдались последствия, аналогичные последствиям после хирургического поражения, демонстрируя, что эффекты хирургического вмешательства, наиболее вероятно, были вызваны потерей этого пептида, а не ACh, присутствующего в синаптических окончаниях.
Эффекты электростимуляции
У животных физиология MOCS изучена гораздо более широко, чем физиология LOCS. Это связано с тем, что миелинизированные волокна MOCS легче электрически стимулировать и регистрировать.[15] Следовательно, относительно мало известно о физиологии LOCS.[23]
Многие исследования проводились на животных in vivo стимулировали оливокохлеарный пучок (ОКБ), используя шоковые стимулы, доставляемые электродами, помещенными на нервный пучок. В этих исследованиях измерялась мощность слухового нерва (АН) со стимуляцией ОКВ и без нее. В 1956 году Галамбос активировал эфферентные волокна кошки, посылая шоковые стимулы на дно четвертого желудочка (на перекресте COCB). Галамбос наблюдал подавление сложных потенциалов действия АН (называемых потенциалом N1), вызванных щелчками низкой интенсивности.[24] Этот основной вывод неоднократно подтверждался (Desmedt and Monaco, 1961; Fex, 1962; Desmedt, 1962; Wiederhold, 1970). Эфферентное подавление N1 также наблюдалось при стимуляции тел клеток MOCS в медиальном SOC,[25] подтверждая, что подавление N1 было результатом стимуляции MOC (не LOC). Совсем недавно несколько исследователей наблюдали подавление нейронного выхода улитки во время стимуляции нижнего бугорка (IC) в среднем мозге, который проецируется на верхний оливарный комплекс (SOC) (Rajan, 1990; Mulders and Robertson, 2000; Ota et al. ., 2004; Чжан, Долан, 2006). Ота и др. (2004) также показали, что подавление N1 в улитке было наибольшим на частоте, соответствующей частотному расположению электрода в IC, что является дополнительным доказательством тонотопической организации эфферентных путей.
Эти находки привели к нынешнему пониманию того, что активность MOC снижает активный процесс OHC, что приводит к частотно-зависимому снижению кохлеарного усиления.
Акустически вызванные ответы MOCS

Электрическая стимуляция ствола головного мозга может привести к (i) стимуляции всего MOCS, (ii) скорости разряда (до 400 с-1), намного большей, чем обычно вызывается звуком (до 60 с-1), и ( iii) электрическая стимуляция нейронов, отличных от волокон MOCS. Следовательно, электрическая стимуляция MOCS может не дать точного указания на его биологическую функцию или естественную величину его воздействия.
Ответ MOCS на звук опосредуется акустическим рефлексом MOC (см. Вставку), который ранее был исследован с использованием методов антероградной и ретроградной маркировки (Aschoff et al., 1988; Robertson and Winter, 1988). Акустическая стимуляция внутренних волосковых клеток посылает нейронный сигнал в задневентральное ядро кохлеара (PVCN), и аксоны нейронов PVCN пересекают ствол мозга, чтобы иннервировать контралатеральные нейроны MOC. У большинства млекопитающих нейроны МОК преимущественно проецируются на контралатеральную сторону (формируя ипсилатеральный рефлекс), а остальные проецируются на ипсилатеральную сторону (формируя контралатеральный рефлекс).
Сила рефлекса является самой слабой для чистых тонов и становится сильнее по мере увеличения полосы пропускания звука (Berlin et al., 1993), поэтому максимальный отклик MOCS наблюдается для широкополосного шума (Guinan et al., 2003). Исследователи измерили эффект стимуляции MOCS звуком. У кошек Либерман (1989) показал, что контралатеральный звук (приводящий к стимуляции MOCS) снижает потенциал N1, подавление которого устраняется при пересечении оливокохлеарного пучка (OCB). У людей наибольшее количество доказательств действия эфферентов было получено из подавления отоакустической эмиссии (ОАЭ) после акустической стимуляции.
Используя акустические стимулы для активации рефлекторного пути MOC, были сделаны записи от отдельных эфферентных волокон у морских свинок.[13] и кошки.[6] Оба исследования подтвердили, что нейроны МОК резко настроены на частоту, как ранее предполагали Коди и Джонстон (1982) и Робертсон (1984). Они также показали, что частота возбуждения нейронов МОС увеличивается по мере увеличения интенсивности звука от 0 до 100 дБ SPL и имеет пороги, сопоставимые (в пределах ~ 15 дБ) с афферентными нейронами. Кроме того, оба исследования показали, что большинство нейронов MOC реагировали на звук, представленный в ипсилатеральном ухе, что согласуется с тем, что большинство нейронов MOC млекопитающих расположены на противоположной стороне.[2][7] Никаких записей от волокон MOC у людей не производилось. потому что инвазивный in vivo эксперименты невозможны. Однако у других видов приматов было показано, что около 50-60% волокон MOC пересекаются (Bodian and Gucer, 1980; Thompson and Thompson, 1986).
Предлагаемые функции MOCS
Предполагаемые функции MOCS делятся на три основные категории; (i) защита улитки от громких звуков, (ii) развитие функции улитки и (iii) обнаружение и различение звуков в шуме.
Защита улитки от громких звуков
Коди и Джонстон (1982) и Раджан и Джонстон (1988a; 1988b) показали, что постоянная акустическая стимуляция (которая вызывает сильную реакцию MOCS (Brown et al., 1998)) снижает тяжесть акустической травмы. Эта защита была сведена на нет в присутствии химического вещества, которое, как известно, подавляет действие оливокохлеарного пучка (OCB) (стрихнин), предполагая, что действие MOCS защищает улитку от громких звуков. Дополнительные доказательства того, что слуховые эфференты играют защитную роль, были предоставлены Раджаном (1995a) и Куджава и Либерман (1997). Оба исследования показали, что потеря слуха, полученная животными из-за воздействия бинаурального звука, была более серьезной при разрыве ПОК. Раджан (1995b) также показал частотную зависимость защиты от МОС, примерно соответствующую распределению волокон МОС в улитке. Другие исследования, подтверждающие эту функцию MOCS, показали, что стимуляция MOC снижает временный сдвиг порога (TTS) и постоянный сдвиг порога (PTS), связанные с длительным воздействием шума (Handrock and Zeisberg, 1982; Rajan, 1988b; Reiter and Liberman, 1995) и что животные с самым сильным рефлексом МОС меньше страдают слухом от громких звуков (Maison and Liberman, 2000). Предложенная биологическая роль MOCS, защита от громких звуков, была оспорена Кирком и Смитом (2003), которые утверждали, что интенсивность звуков, используемых в экспериментах (≥105 дБ SPL), редко или никогда не встречается в природе, и, следовательно, защитный механизм для звуков такой интенсивности не мог возникнуть. Это утверждение (что MOC-опосредованная защита улитки является эпифеноменом) недавно было оспорено Darrow et al. (2007), которые предположили, что LOCS обладает антиэксайтотоксическим действием, косвенно защищая улитку от повреждений.
Развитие функции улитки
Также существуют доказательства роли оливокохлеарного пучка (ОСБ) в развитии функции улитки. Либерман (1990) измерил ответы отдельных волокон АН взрослых кошек в течение 6 месяцев после разрыва ОКВ. Либерман не обнаружил никаких изменений пороговых значений волокон AN, кривых настройки и функций ввода-вывода. Walsh et al. (1998) провели аналогичный эксперимент, однако исследователи вырезали ОКБ новорожденных кошек и записали данные с волокон АН через год. У кошек без эфферентного входа в улитку регистрировались повышенные пороги AN, пониженная резкость кривых настройки и пониженные SR. Walsh et al. (1998) предположили, что неонатальная дефферентация мешает нормальному развитию и функционированию половых органов, следовательно, вовлекает ОКБ в развитие активных процессов в улитке.
Обнаружение и различение звуков в шуме
Обсуждаемые до сих пор эффекты, вызванные МОС, наблюдались в экспериментах, проводимых в тишине (обычно в звукоизолирующих кабинах или комнатах). Однако измерение реакции улитки на звуки в этих условиях может не выявить истинную биологическую функцию MOCS, поскольку эволюционирующие млекопитающие редко находятся в безмолвных ситуациях, а MOCS особенно чувствительна к шуму (Guinan et al., 2003). Первые эксперименты по изучению эффектов стимуляции MOC в присутствии шума были проведены на морских свинках Nieder и Nieder (1970a, 1970b, 1970c), которые измерили улитковую продукцию, вызванную щелчками, представленными в постоянном фоновом шуме (BGN). В этом состоянии они обнаружили, что потенциал N1, вызванный щелчками, усиливался в течение периода стимуляции MOC. Это открытие было подтверждено с использованием как электростимуляции (Dolan, Nuttall, 1988; Winslow and Sachs, 1987), так и акустической активации (Kawase et al., 1993, Kawase and Liberman, 1993) MOCS млекопитающих. Уинслоу и Сакс (1987) обнаружили, что стимуляция ОКБ:
«... позволяет волокнам слухового нерва сигнализировать об изменениях уровня тона с изменениями скорости разряда при более низком соотношении сигнал / шум, чем это было бы возможно в противном случае». (Страница 2002)
Одна из интерпретаций этих результатов заключается в том, что стимуляция МОС избирательно снижает реакцию слухового нерва на постоянный фоновый шум, что позволяет лучше реагировать на кратковременный звук.[15] Таким образом, стимуляция МОС уменьшит эффект подавляющей и адаптивной маскировки, и по этой причине процесс получил название «разоблачения» или «антимаскирования» (Kawase et al., 1993, Kawase and Liberman, 1993) . Было высказано предположение, что антимаскирование происходит аналогичным образом у людей (Kawase and Takasaka, 1995) и имеет последствия для избирательного прослушивания, поскольку быстрое разоблачение звука, возникающего в результате активации MOC, увеличивает общее отношение сигнал / шум (SNR). , что способствует лучшему обнаружению звука цели.
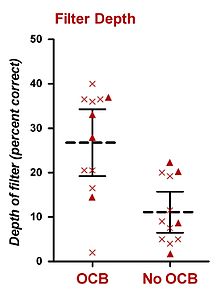
У людей психофизические эксперименты, проводимые при постоянном BGN, также показали, что оливокохлеарный пучок (OCB) участвует в избирательном прослушивании. Исследование, возможно, наиболее подходящее для этого тезиса, было проведено Шарфом и его коллегами. В 1993 году Шарф и др. представили данные восьми пациентов, перенесших одностороннюю вестибулярную неврэктомию для лечения болезни Меньера, процедуры, при которой рассекается ОКБ (предположительно, как MOCS, так и LOCS). Шарф и др. (1993) не обнаружили каких-либо явных различий в пороговых значениях тонов шума у испытуемых до и после операции. Вскоре после этого открытия Scharf et al. (1994, 1997) выполнили комплексный набор психофизических экспериментов на шестнадцати пациентах, перенесших одностороннюю вестибулярную неврэктомию (включая восемь исходных пациентов).[26][27] Они измерили производительность в психофизических задачах прослушивания до и после операции и не обнаружили существенной разницы в производительности для (i) обнаружения тонов, (ii) различения интенсивности тонов, (iii) частотного различения тонов, (iv) адаптации громкости, и (v) обнаружение тонов в режекторном шуме.[26][27] Их единственный положительный результат заключался в том, что большинство пациентов обнаруживали неожиданные звуки в прооперированном ухе лучше, чем в здоровом ухе или в том же ухе до операции. Этот результат был получен с использованием процедуры усеченного зондового сигнала, которая заставляла пациента ожидать определенной частоты в каждом испытании. Двенадцать испытуемых завершили этот эксперимент.[26][27] Их процедура была аналогична процедуре Гринберга и Ларкина (1968), за исключением того, что только 50% испытаний (не 77%) содержали цель, частота которой совпадала с частотой слухового сигнала. Остальные 50% испытаний содержат зонд, частота которого отличается от частоты сигнала. Кроме того, использовались только две частоты зонда, одна из которых была выше целевой, а другая - ниже целевой. Все испытания содержали слуховой сигнал (на целевой частоте) до первого интервала наблюдения. Результаты были использованы для построения основного фильтра внимания, который отображал уровень обнаружения ожидаемой (и полученной) целевой частоты и двух неожиданных частот зонда.[26][27] Из двух опубликованных отчетов (Scharf et al., 1994, 1997) уши, у которых был поврежден ОКБ, показали фильтр внимания со средней глубиной примерно на 15%, что меньше, чем те уши, у которых ОКБ не был поврежден.[26][27] Хотя нет возможности эмпирически преобразовать это значение в дБ, грубая оценка, основанная на психометрических функциях, представленных Грин и Светс (1966), дает значение 2–3 дБ. Их результаты представлены на вставке.[26]
Шарф и его коллеги утверждали, что секционирование ОКБ у этих пациентов привело к подавлению неожиданных частот. Этот эффект присутствовал не у всех субъектов, и наблюдались большие различия между субъектами. Тем не менее, никакие другие психофизические характеристики слуха не пострадали после секционирования OCB. Шарф и др. (1997) пришли к выводу, что OCB-опосредованное подавление звуков в улитке отвечает за подавление неожиданных звуков и, таким образом, играет роль в избирательном внимании при нормальном слухе.[26] В отличие от теории Шарфа, Tan et al. (2008) утверждали, что роль OCB в избирательном прослушивании относится к усилению подаваемого или ожидаемого тона. Это усиление может быть вызвано действием MOCS на внешние волосковые клетки, что приводит к антимаскированию.[28]
Хотя эксперименты Шарфа и др. (1993, 1994, 1997) не смогли выявить каких-либо явных различий в основных психофизических характеристиках слуха (кроме обнаружения неожиданных звуков), многие другие исследования с участием как животных, так и людей указали на наличие OCB. в задачах «слушать в шуме» с использованием более сложных стимулов. При постоянном BGN макаки-резусы с интактными ОКВ лучше справляются с задачами распознавания гласных, чем без них (Dewson, 1968). У кошек интактный ОКБ ассоциируется с лучшей идентификацией гласных (Heinz et al., 1998), локализацией звука (May et al., 2004) и различением интенсивности (May and McQuone, 1995). Все эти исследования проводились в постоянных левах. У людей измерения различения речи в шуме проводились у лиц, перенесших одностороннюю вестибулярную неврэктомию (в результате секционирования ОКВ). Giraud et al. (1997) наблюдали небольшое преимущество здорового уха над оперированным ухом в распознавании фонем и разборчивости речи в BGN. Шарф и др. (1988) ранее исследовали роль слухового внимания во время восприятия речи и предположили, что различению речи в шуме помогает фокусировка внимания на частотных диапазонах. В 2000 году Цзэн и др. Сообщили, что вестибулярная нейрэктомия напрямую не влияет на пороги чистого тона или различение интенсивности,[29] подтверждая более ранние выводы Scharf et al. 1994; 1997 г.[26][27] Для задач «Слушать в шуме» они обнаружили ряд несоответствий между здоровым и оперированным ухом. В соответствии с более ранними выводами May и McQuone (1995), различение интенсивности шума было немного хуже в ухе без ввода оливокохлеарного пучка (OCB). Тем не менее, главный вывод Цзэн и др. Касался эффекта «перерегулирования», который был значительно снижен (~ 50%) в оперированных ушах.[29] Этот эффект был впервые обнаружен Цвикером (1965) и охарактеризован как повышенный порог обнаружения тона, когда он присутствует в начале шума, по сравнению с тем, когда он представлен в постоянном установившемся шуме.[30] Zeng et al. предположил, что это открытие согласуется с антимаскированием, вызванным MOCS; то есть, вызванная MOCS антимаскировка отсутствует в начале шума, но становится активной во время установившегося шума. Эта теория подтверждается динамикой активации MOC;[6][31] будучи похожим на динамику эффекта выброса (Zwicker, 1965),[30] а также нарушение эффекта перерегулирования у субъектов с нейросенсорной тугоухостью, для которых MOCS, скорее всего, будет неэффективным (Bacon and Takahashi, 1992).
Рекомендации
- ^ Расумссен, Г. Л. (1960). «Глава 8: Эфферентные волокна улиткового нерва и кохлеарного ядра». In Rasmussen, G.L .; Windle, W. F. (ред.). Нейронные механизмы слуховой и вестибулярной системы. Спрингфилд, Иллинойс: Чарльз С. Томас. С. 105–115.
- ^ а б c Warr, WB .; Guinan, JJ. (Сентябрь 1979 г.). «Эфферентная иннервация кортиевого органа: две отдельные системы». Brain Res. 173 (1): 152–5. Дои:10.1016/0006-8993(79)91104-1. PMID 487078.
- ^ WARR, W.B .; ГУИНАН, J.J. Jr .; БЕЛЫЙ, Дж. (1986). Ричард Альтшулер; Ричард П. Боббин; Дуглас В. Хоффман (ред.). Организация эфферентных волокон: латеральная и медиальная оливокохлеарные системы.. Нейробиология слуха: улитка. Нью-Йорк: Raven Press. ISBN 978-0-89004-925-9. OCLC 14243197.
- ^ Guinan, JJ .; Warr, WB .; Норрис, BE. (Декабрь 1983 г.). «Дифференциальные оливокохлеарные проекции латеральных и медиальных зон верхнего оливкового комплекса». J Comp Neurol. 221 (3): 358–70. Дои:10.1002 / cne.902210310. PMID 6655089.
- ^ Vetter, DE .; Мугнаини, Э. (1992). «Распределение и дендритные особенности трех групп оливокохлеарных нейронов крысы: исследование с двумя ретроградными индикаторами токсина холеры». Анат. Эмбриол. 185: 1–16. Дои:10.1007 / bf00213596. PMID 1736680.
- ^ а б c d Liberman, MC .; Браун, MC. (1986). «Физиология и анатомия одиночных оливокохлеарных нейронов кошки». Слушайте Res. 24 (1): 17–36. Дои:10.1016/0378-5955(86)90003-1. PMID 3759672.
- ^ а б У. Б. Варр (сентябрь – октябрь 1980 г.). «Эфферентные компоненты слуховой системы». Анналы отологии, ринологии и ларингологии. Добавка. 89 (5, Пет 2): 114–120. Дои:10.1177 / 00034894800890S527. PMID 6786165.
- ^ Thompson, GC .; Томпсон, AM. (Декабрь 1986). «Olivocochlear нейроны в стволе мозга белки обезьяны». J Comp Neurol. 254 (2): 246–58. Дои:10.1002 / cne.902540208. PMID 3540042.
- ^ Робертсон и др., 1989 г.
- ^ Azeredo, WJ .; Климент, МЛ .; Морли, Б.Дж .; Релкин, Е .; Слепецкий, Н.Б .; Sterns, A .; Warr, WB .; Weekly, JM .; Woods, CI. (Август 1999 г.). «Olivocochlear нейроны в шиншилле: исследование ретроградной флуоресцентной маркировки». Слушайте Res. 134 (1–2): 57–70. Дои:10.1016 / S0378-5955 (99) 00069-6. PMID 10452376.
- ^ Арнесен, АР. (1984). «Популяция волокон вестибулокохлеарного анастомоза у человека». Акта Отоларингол. 98 (5–6): 501–18. Дои:10.3109/00016488409107591. PMID 6524346.
- ^ Арнесен, АР. (1985). «Численные оценки структур в ядрах улитки, афферентах и эфферентах улитки». Акта отоларингол Suppl. 423: 81–4. Дои:10.3109/00016488509122916. PMID 3864352.
- ^ а б Робертсон, Д .; Гаммер, М. (1985). «Физиологическая и морфологическая характеристика эфферентных нейронов в улитке морской свинки». Слушайте Res. 20 (1): 63–77. Дои:10.1016/0378-5955(85)90059-0. PMID 2416730.
- ^ Робертсон, Д; Андерсон, С .; Коул, К. С. (1987). «Разделение эфферентных проекций на разные витки улитки морской свинки». Слуховые исследования. 25 (1): 69–76. Дои:10.1016/0378-5955(87)90080-3. PMID 3804858.
- ^ а б c Гуинан, Джон Дж. Младший (1996). Питер Даллос; Артур Н. Поппер; Ричард Р. Фэй (ред.). Физиология оливокохлеарных эфферентов. Улитка. Нью-Йорк: Спрингер. С. 435–502. ISBN 978-0-387-94449-4. OCLC 33243443.
- ^ Warr, WB .; Beck, JE .; Neely, ST. (1997). «Эфферентная иннервация внутренней области волосковых клеток: истоки и окончания двух латеральных оливокохлеарных систем». Слушайте Res. 108 (1): 89–111. Дои:10.1016 / S0378-5955 (97) 00044-0. PMID 9213126.
- ^ Elgoyhen, AB .; Johnson, DS .; Boulter, J .; Vetter, DE .; Хайнеманн, С. (ноябрь 1994 г.). «Альфа 9: рецептор ацетилхолина с новыми фармакологическими свойствами, выраженный в волосковых клетках улитки крысы». Клетка. 79 (4): 705–15. Дои:10.1016 / 0092-8674 (94) 90555-Х. PMID 7954834.
- ^ Elgoyhen, AB .; Vetter, DE .; Katz, E .; Rothlin, CV .; Heinemann, SF; Боултер, Дж. (Март 2001 г.). «альфа10: детерминанта функции никотинового холинергического рецептора в вестибулярных и механосенсорных волосковых клетках млекопитающих». Proc Natl Acad Sci U S A. 98 (6): 3501–6. Дои:10.1073 / pnas.051622798. ЧВК 30682. PMID 11248107.
- ^ Katz, E .; Вербицкий, М .; Rothlin, CV .; Vetter, DE .; Heinemann, SF; Элгойхен, АБ. (Март 2000 г.). «Высокая проницаемость для кальция и кальциевый блок никотинового рецептора ацетилхолина альфа9». Слушайте Res. 141 (1–2): 117–28. Дои:10.1016 / S0378-5955 (99) 00214-2. PMID 10713500.
- ^ Lioudyno; и другие. (2004). «А» Синаптоплазматическая цистерна «опосредует быстрое ингибирование волосковых клеток улитки». Журнал неврологии. 24 (49): 11160–4. Дои:10.1523 / JNEUROSCI.3674-04.2004. ЧВК 6730265. PMID 15590932.
- ^ Уолш; и другие. (1998). «Долгосрочные эффекты разделения оливокохлеарной связки у новорожденных кошек». Журнал неврологии. 18 (10): 3859–69. Дои:10.1523 / JNEUROSCI.18-10-03859.1998. ЧВК 6793155. PMID 9570815.
- ^ Веттер; и другие. (2002). «У мышей с дефицитом урокортина наблюдается нарушение слуха и повышенное тревожное поведение». Природа Генетика. 31 (4): 363–9. Дои:10,1038 / ng914. PMID 12091910.
- ^ Groff, JA .; Либерман, М.С. (Ноябрь 2003 г.). «Модуляция афферентного ответа улитки латеральной оливокохлеарной системой: активация посредством электрической стимуляции нижнего холмика» (PDF). J Нейрофизиол. 90 (5): 3178–200. Дои:10.1152 / ян.00537.2003. HDL:1721.1/28596. PMID 14615429.
- ^ ГАЛАМБОС, Р. (сентябрь 1956 г.). «Подавление активности слухового нерва путем стимуляции эфферентных волокон улитки» (PDF). J Нейрофизиол. 19 (5): 424–37. Дои:10.1152 / jn.1956.19.5.424. PMID 13367873.
- ^ Гиффорд, ML .; Guinan, JJ. (1987). «Влияние электрической стимуляции медиальных оливокохлеарных нейронов на ипсилатеральные и контралатеральные улитковые реакции». Слушайте Res. 29 (2–3): 179–94. Дои:10.1016/0378-5955(87)90166-3. PMID 3624082.
- ^ а б c d е ж грамм час я Шарф, В .; Magnan, J .; Чайс, А. (январь 1997 г.). «О роли оливокохлеарной связки в слухе: 16 тематических исследований». Слушайте Res. 103 (1–2): 101–22. Дои:10.1016 / S0378-5955 (96) 00168-2. PMID 9007578.
- ^ а б c d е ж Шарф, В .; Magnan, J .; Цанга, L .; Ulmer, E .; Чайс, А. (май 1994 г.). «О роли оливокохлеарной связки в слухе: тематическое исследование». Слушайте Res. 75 (1–2): 11–26. Дои:10.1016/0378-5955(94)90051-5. PMID 8071137.
- ^ Tan, MN .; Робертсон, Д .; Хаммонд, гр. (Июль 2008 г.). «Отдельные вклады повышенной и подавленной чувствительности на слуховой фильтр внимания». Слуховые исследования. 241 (1–2): 18–25. Дои:10.1016 / j.heares.2008.04.003. PMID 18524512.
- ^ а б Zeng, FG .; Мартино, КМ .; Linthicum, FH .; Соли, СД. (Апрель 2000 г.). «Слуховое восприятие у испытуемых вестибулярной неврэктомии». Слушайте Res. 142 (1–2): 102–12. Дои:10.1016 / S0378-5955 (00) 00011-3. PMID 10748333.
- ^ а б Цвикер, Э. (июль 1965 г.). «Временные эффекты при одновременной маскировке и громкости» (PDF). J Acoust Soc Am. 38: 132–41. Дои:10.1121/1.1909588. PMID 14347604.
- ^ Backus, BC .; Guinan, JJ. (Май 2006 г.). «Динамика медиального оливокохлеарного рефлекса человека». J Acoust Soc Am. 119 (5, Пет. 1): 2889–904. Дои:10.1121/1.2169918. PMID 16708947.