SK канал - SK channel
| Калиевый канал SK, активированный кальцием | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 SK Channel | |||||||||
| Идентификаторы | |||||||||
| Символ | SK_channel | ||||||||
| Pfam | PF03530 | ||||||||
| ИнтерПро | IPR015449 | ||||||||
| |||||||||
SK каналы (активируемые кальцием калиевые каналы с малой проводимостью) являются подсемейством Ca2+-активированный K+ каналы.[1] Их называют так из-за их небольшой проводимости одного канала, порядка 10 pS.[2] Каналы SK являются разновидностью ионный канал позволяя калий катионы проникают через клеточную мембрану и активируются (открываются) при увеличении концентрации внутриклеточного кальция через Кальциевые каналы N-типа. Их активация ограничивает частоту срабатывания потенциалы действия и важно для регулирования постгиперполяризация в нейронах Центральная нервная система а также многие другие типы электрически возбудимых клеток. Это достигается за счет гиперполяризационной утечки положительно заряженных ионов калия по градиенту их концентрации во внеклеточное пространство. Этот гиперполяризация вызывает мембранный потенциал стать более негативным.[3] Считается, что каналы SK участвуют в синаптическая пластичность и поэтому играют важную роль в обучении и памяти.[4]
Функция
Каналы SK выражаются через Центральная нервная система. Они очень консервативны у млекопитающих, а также у других организмов, таких как Drosophila melanogaster и Caenorhabditis elegans.[5] Каналы SK специфически вовлечены в средний постгиперполяризующий потенциал (mAHP). Они влияют как на внутреннюю возбудимость нейронов, так и на синаптическую передачу. Они также участвуют в передаче сигналов кальция.[6] Каналы SK контролируют частоту разряда потенциала действия в нейронах гиппокампа, дофаминергических нейронах среднего мозга, дорсальных нейронах блуждающего нерва, симпатических нейронах, ретикулярных нейронах таламуса, нейронах нижних олив, спинномозговых и подъязычных мотонейронах, митральных клетках в обонятельной луковице и корковых нейронах.[3]
Структура
Калиевые каналы SK имеют ту же базовую архитектуру, что и Shaker-like потенциалзависимые калиевые каналы.[7] Четыре субъединицы объединяются, образуя тетрамер. Каждая из субъединиц имеет шесть трансмембранных гидрофобных альфа-спиральных доменов (S1-S6). Петля между S5 и S6, называемая P-петля - образует порообразующую область, которая всегда обращена к центру канала.[8] В каждом из подразделений по шесть гидрофобный альфа спиральный домены, которые вставляются в клеточную мембрану. Петля между пятым и шестым трансмембранными доменами образует ион калия. фильтр селективности. SK-каналы могут собираться как гомотетрамерные каналы или как гетеротетрамерные каналы, состоящие из более чем одного подтипа SK-каналов. Кроме того, калиевые каналы SK тесно связаны с белком. кальмодулин, что объясняет чувствительность этих каналов к кальцию.[7][9] Кальмодулин участвует как субъединица самого канала, связанная с цитоплазматической С-концевой областью пептида, называемой кальмодулин-связывающий домен (CaMBD).[10]
Дополнительная ассоциация фосфорилирующей киназы CK2 и дефосфорилирующая фосфатаза PP2A на цитоплазматической стороне белка позволяют обогащенный Ca2+-чувствительность и, следовательно, модуляция кинетики.[11] CK2 служит для фосфорилирования СаМ, связанного с SKCa, по остатку Т80, а не самих спиралей каналов, для снижения чувствительности к кальцию. Это может быть достигнуто только тогда, когда поры канала закрыты. PP2A дефосфорилирует этот остаток при ингибировании CK2.[10] Фильтр селективности всех подтипов каналов SK - будь то SK1, SK2, SK3 или SK4 - очень консервативен и отражает избирательность, наблюдаемую в любом калиевый канал, последовательность аминокислотных остатков GYGD на порообразующей петле.[12] Эти каналы считаются независимыми от напряжения, поскольку они содержат только два из семи положительно заряженных аминокислотных остатков, которые обычно наблюдаются в прототипах. потенциалзависимый калиевый канал.[8]
Классификация
Семейство каналов SK состоит из 4 членов - SK1, SK2, SK3, и SK4. SK4 часто называют IK (Промежуточная проводимость) из-за его более высокой проводимости 20 - 80 пс.[13]
| Канал | Ген | Псевдонимы | Ассоциированные субъединицы |
| SK1 | KCNN1 | Kок2.1 | кальмодулин, PP2A, CK2 |
| SK2 | KCNN2 | Kок2.2 | кальмодулин, PP2A, CK2 |
| SK3 | KCNN3 | Kок2.3 | кальмодулин, PP2A, CK2 |
| SK4 | KCNN4 | Kок3.1 | кальмодулин, PP2A, CK2 |
Механизм стробирования
Механизм затвора канала SK управляется внутриклеточный уровень кальция.[5] Кальций попадает в клетку через кальциевые каналы, активируемые напряжением, а также через рецепторы NMDA.[3] Кальций не связывается напрямую с каналом SK. Даже в отсутствие кальция канал SK связывается с C-долей белка кальмодулина (CaM). Когда N-доля связывает кальций, она захватывает линкер S4-S5 на внутриклеточной субъединице SK-канала. Когда каждый из четырех линкеров S4-S5 связан с N-долей кальмодулина, канал SK изменяет конформацию. Кальмодулин подталкивает линкер S4-S5, чтобы позволить расширение пересечения пучка S6, что приводит к открытию поры. Идея о том, что это переводит канал из тетрамер из мономеры в сложенный димер димеров, которое приводит к вращению СаМ-связывающих доменов, в настоящее время отказано, и самые последние наблюдения несовместимы с предположением, что это вращение вызывает механическое открытие ворот канала.[5] Постоянная времени активации канала SK составляет примерно 5 мс. Когда уровень кальция истощен, постоянная времени для деактивации канала находится в пределах 15–60 мс.[14]
Блокираторы
Все каналы SK могут быть фармакологически заблокированы солями четвертичного аммония нейротоксина растительного происхождения. бикукуллин.[15] Кроме того, каналы SK (SK1-SK3), но не SK4 (IK), чувствительны к блокаде пчелиным токсином. апамин,[16] и скорпион яды тамапин и харибдотоксин (ChTx), все через конкурентный антагонизм для доступа к устью порообразования.[17] Все известные блокаторы конкурируют примерно за один и тот же сайт связывания, поры, во всех подтипах. Это обеспечивает физическую блокировку поры канала.[18] Поскольку все блокираторы универсальны для всех трех типов каналов SK, существует невероятно узкий терапевтическое окно что не позволяет блокировать определенный подтип канала SK.[11] Соли четвертичного аммония, такие как бикукуллин и тетраэтиламмоний (ТЭА) проникают в поры через фильтр селективности, действуя как имитатор калия на стадии дегидратации при проникновении в поры.[18]
Следующие молекулы являются другими токсины и органические соединения которые также подавляют все три малых подтипа SK каналов в любой (даже минимальной) степени:[11]
- Деквалиний
- d-тубокурарин
- UCL-1684
- UCL-1848
- Ципрогептадин
- Флуоксетин, активный ингредиент в Прозак
- NS8593
- Сциллатоксин (Лейуротоксин-I)
- Лей-Даб7
- N-метил-лауданозин
- N-Me-бикукуллин
- Панкуроний
- Атракуриум
- 1-этил-1H-бензо [d] имидазол-2 (3H) -он
- 6,7-дихлор-3- (гидроксиимино) индолин-2-он
- N-циклогексил-2- (3,5-диметил-1H-пиразол-1-ил) -6-метилпиримидин-4-амин
- (R) -N- (1,2,3,4-тетрагидронафталин-1-ил) -1H-бензо [d] имидазол-2-амин
Модуляторы
Аллостерические модуляторы малых SK-каналов работают за счет изменения кажущейся кальциевой чувствительности каналов. Примеры включают:
- Рилузол
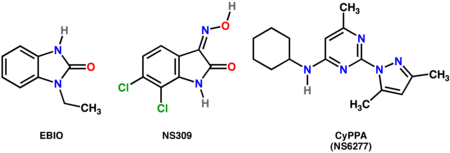
- Неселективные положительные модуляторы SK каналов: EBIO (1-Eтил-2-BэнзимядазолинОne),[19] NS309 (6,7-дихлор-1H-индол-2,3-дион 3-оксим)[20]
- Селективные положительные модуляторы СК-2 и СК-3: CyPPA (NS6277; Сайклогексил- (2- (3,5-диметил-пиразол-1-ил) -6-метил-пиримидин-4-ил) -Амоя)[21]
Синаптическая пластичность и долгосрочное потенцирование
В дендритных шипах каналы SK напрямую связаны с NMDA рецепторы. Помимо активации потоком кальция через потенциалзависимые кальциевые каналы, каналы SK могут активироваться кальцием, протекающим через рецепторы NMDA, что происходит после деполяризации постсинаптической мембраны.[10] Эксперименты с апамином показали, что именно блокирование SK-каналов может улучшить обучение и долгосрочное потенцирование. Кроме того, нейротрофический фактор головного мозга (BDNF) вызывает подавление SK-каналов, что облегчает долгосрочное потенцирование. Повышение активности SK-каналов имеет противоположный эффект и ухудшает учусь.[5] Увеличение активности SK-каналов, происходящее с течением времени, может быть связано со снижением пластичности и памяти, которое наблюдается при старении.[22]
Роль в болезни Паркинсона
Считается, что дисфункция калиевых каналов, в том числе SK-каналов, играет роль в патогенезе болезнь Паркинсона (PD), прогрессивный нейродегенеративное расстройство.
Блокираторы каналов СК контролируют скорость стрельбы (количество потенциалы действия произведенный нейрон в заданное время) и паттерна активации (способ распределения потенциалов действия во времени) через их производство м-AHP. Активаторы SK-каналов снижают частоту возбуждения и чувствительность нейронов к возбуждающим стимулам, тогда как блокаторы SK-каналов увеличивают скорость возбуждения и чувствительность к возбуждающим стимулам. Это имеет важное значение для функции дофаминергических нейронов.[23] Например, количество дофамина, выделяемого дофаминергическими нейронами среднего мозга, намного выше, когда частота активации увеличивается, чем когда они стреляют с постоянной скоростью.
Каналы SK широко экспрессируются в среднем мозге. дофаминергический нейроны. Для регулирования аффинности SK к ионам кальция использовались различные фармакологические методы, тем самым модулируя возбудимость черная субстанция дофаминергические нейроны. Блокада SK-каналов in vivo увеличивает скорость возбуждения клеток черной субстанции, что увеличивает количество дофамина, высвобождаемого из синаптических окончаний.[23] Когда в цитозоле накапливается большое количество дофамина, индуцируется повреждение клеток из-за накопления свободных радикалов и повреждения митохондрий. Кроме того, были использованы методы модуляции SK-каналов с целью изменения дофамина. фенотип нейронов. После потери TH + (тирозингидроксилаза -положительных) нейронов компактной черной субстанции (SNc) из-за нейродегенерации, вызванной паркинсонизмом, количество этих нейронов может частично восстановиться за счет «сдвига» клеточного фенотипа с TH- (тирозингидроксилаза-отрицательный) на TH +. Количество TH + нейронов может быть изменено путем модуляции канала SK; Чтобы быть конкретным, инфузия агонистов SK в черную субстанцию увеличивает количество TH + нейронов, тогда как инфузия антагониста SK снижает количество TH + нейронов. Причина такой связи между SK-каналами и экспрессией TH может быть связана с нейропротекцией от дофамина. токсичность.[23]
В качестве терапевтических вариантов для улучшения симптомов БП были предложены два противоречащих друг другу метода:
Подавление SK каналов
- Ингибирование каналов SK, а именно блокирование каналов SK3, увеличивает частоту возбуждения дофаминергических нейронов, тем самым увеличивая высвобождение дофамин. Поэтому считается, что применение блокаторов каналов SK3 у пациентов с БП может облегчить краткосрочные двигательные симптомы.
- Однако ингибирование также приводит к уменьшению количества нейронов TH + компактной черной субстанции (SNc) в клетке, что приводит к снижению синтеза дофамина в долгосрочной перспективе.
Фасилитация каналов SK
- Усиление функции SK-каналов увеличивает количество TH + нейронов компактной черной субстанции (SNc) в клетке, тем самым поддерживая синтез дофамина в течение длительного времени.
- Однако облегчение SK-каналов снижает частоту возбуждения дофаминергических нейронов в краткосрочной перспективе.
Рекомендации
- ^ Бонд CT, Мэйли Дж, Адельман ДжП (1999). «Каналы калия, активированные кальцием малой проводимости». Анна. Акад. Наука. 868 (1): 370–8. Дои:10.1111 / j.1749-6632.1999.tb11298.x. PMID 10414306.
- ^ Кёлер М., Хиршберг Б., Бонд К.Т., Кинзи Дж. М., Маррион Н. В., Мэйли Дж., Адельман Дж. П. (1996). «Активированные кальцием калиевые каналы малой проводимости из мозга млекопитающих». Наука. 273 (5282): 1709–14. Дои:10.1126 / science.273.5282.1709. PMID 8781233.
- ^ а б c Faber ES, Sah P (2007). «Функции SK каналов в центральных нейронах». Clin. Exp. Pharmacol. Физиол. 34 (10): 1077–83. Дои:10.1111 / j.1440-1681.2007.04725.x. PMID 17714097.
- ^ Стакман Р.В., Хаммонд Р.С., Линардатос Э., Герлах А., Мэйли Дж., Адельман Дж. П., Цунопулос Т. (2002). "Малая проводимость Ca2+-активированный K+ каналы модулируют синаптическую пластичность и кодирование памяти ». J. Neurosci. 22 (23): 10163–71. Дои:10.1523 / JNEUROSCI.22-23-10163.2002. PMID 12451117.
- ^ а б c d Адельман Дж. П., Мэйли Дж, Сах П (2012). "Малая проводимость Ca2+-активированный K+ каналы: форма и функции ». Анну. Преподобный Physiol. 74: 245–69. Дои:10.1146 / аннурев-физиол-020911-153336. PMID 21942705.
- ^ Долга А.М., Терполилли Н., Кепура Ф., Нейхольт И.М., Кнаус Х.Г., Д'Орси Б., Прен Дж. Х., Эйзель У.М., Плант Т, Плеснила Н., Калмзее С. (апрель 2011 г.). «Активация каналов KCa2 предотвращает нарушение регуляции [Ca2 +] i и снижает гибель нейронов после токсичности глутамата и церебральной ишемии». Cell Death Dis. 2 (e147): e147. Дои:10.1038 / cddis.2011.30. ЧВК 3122061. PMID 21509037.
- ^ а б Мэйли Дж, Бонд CT, Херсон PS, Ли WS, Адельман JP (2004). "Малая проводимость Ca2+-активированный K+ каналы и кальмодулин ». J. Physiol. 554 (Pt 2): 255–61. Дои:10.1113 / jphysiol.2003.049072. ЧВК 1664776. PMID 14500775.
- ^ а б Стокер М. (октябрь 2004 г.). "Ca2+-активированный K+ каналы: молекулярные детерминанты и функции семейства SK ». Обзоры природы Неврология. 5 (10): 758–70. Дои:10.1038 / номер 1516. PMID 15378036.
- ^ Шумахер М.А., Ривард А.Ф., Bächinger HP, Adelman JP (2001). "Структура стробирующей области Ca2+-активированный K+ канал в комплексе с Ca2+/ кальмодулин ». Природа. 410 (6832): 1120–4. Дои:10.1038/35074145. PMID 11323678.
- ^ а б c Лухан Р., Мэйли Дж, Адельман Дж. П. (июль 2009 г.). «Новые площадки действия для каналов ГИРК и СК». Обзоры природы Неврология. 10 (7): 475–80. Дои:10.1038 / номер 2668. PMID 19543219.
- ^ а б c Weatherall KL, Goodchild SJ, Jane DE, Marrion NV (июль 2010 г.). «Кальций-активируемые калиевые каналы малой проводимости: от структуры к функции». Прог. Нейробиол. 91 (3): 242–55. Дои:10.1016 / j.pneurobio.2010.03.002. PMID 20359520.
- ^ Бернеш С., Ру Б. (апрель 2005 г.). «Вентиль в фильтре селективности калиевых каналов». Структура. 13 (4): 591–600. Дои:10.1016 / j.str.2004.12.019. PMID 15837197.
- ^ Вергара С., Латорре Р., Маррион Н.В., Адельман Дж. П. (1998). «Калиевые каналы, активируемые кальцием». Текущее мнение в нейробиологии. 8 (Пт 3): 321–9. Дои:10.1016 / S0959-4388 (98) 80056-1. PMID 9687354.
- ^ Беркфельд Х, Факлер Б., Шульте У. (октябрь 2010 г.). "Ca2+-активированный K+ каналы: от белковых комплексов к функции ». Physiol. Rev. 90 (4): 1437–59. Дои:10.1152 / Physrev.00049.2009. PMID 20959620.
- ^ Хавалед Р., Брюнинг-Райт А., Адельман Дж. П., Мэйли Дж. (1999). «Бикукуллиновый блок кальций-активируемых калиевых каналов малой проводимости». Арка Пфлюгерса. 438 (3): 314–21. Дои:10.1007 / s004240050915. PMID 10398861.
- ^ Блац А.Л., Маглеби К.Л. (1986). "Одиночный блокированный апамином Ca-активированный K+ каналы малой проводимости в культивируемых скелетных мышцах крысы ». Природа. 323 (6090): 718–20. Дои:10.1038 / 323718a0. PMID 2430185.
- ^ Pedarzani P, D'hoedt D, Doorty KB, Wadsworth JD, Joseph JS, Jeyaseelan K, Kini RM, Gadre SV, Sapatnekar SM, Stocker M, Strong PN (2002). "Тамапин, пептид яда индийского красного скорпиона (Mesobuthus tamulus), который нацелен на низкую проводимость Ca2+-активированный K+ каналы и токи постгиперполяризации в центральных нейронах ». J. Biol. Chem. 277 (48): 46101–9. Дои:10.1074 / jbc.M206465200. PMID 12239213.
- ^ а б Дилли С., Лами С., Маррион Н.В., Льежуа Дж. Ф., Сойтин В. (август 2011 г.). «Модуляторы ионных каналов: больше разнообразия, чем считалось ранее». ChemBioChem. 12 (12): 1808–12. Дои:10.1002 / cbic.201100236. PMID 21726033.
- ^ Педарзани П., Мосбахер Дж., Ривард А., Чинголани Л.А., Оливер Д., Стокер М., Адельман Дж. П., Факлер Б. (2001). "Контроль электрической активности в центральных нейронах путем модуляции стробирования небольшой проводимости Ca2+-активированный K+ каналы". J. Biol. Chem. 276 (13): 9762–9. Дои:10.1074 / jbc.M010001200. PMID 11134030.
- ^ Стробаек Д., Тойбер Л., Йоргенсен Т.Д., Аринг П.К., Кьяер К., Хансен Р.С., Олесен С.П., Кристоферсен П., Скаанинг-Йенсен Б. (2004). "Активация человеческих IK и SK Ca2+-активированный K+ каналы по NS309 (6,7-дихлор-1H-индол-2,3-дион 3-оксим) ». Биохим. Биофиз. Acta. 1665 (1–2): 1–5. Дои:10.1016 / j.bbamem.2004.07.006. PMID 15471565.
- ^ Hougaard C, Eriksen BL, Jørgensen S, Johansen TH, Dyhring T, Madsen LS, Strøbaek D, Christophersen P (2007). «Селективная положительная модуляция подтипов SK3 и SK2 малой проводимости Ca2+-активированный K+ каналы". Br. J. Pharmacol. 151 (5): 655–65. Дои:10.1038 / sj.bjp.0707281. ЧВК 2014002. PMID 17486140.
- ^ Tzounopoulos T, Stackman R (декабрь 2003 г.). «Повышение синаптической пластичности и памяти: роль Са малой проводимости2+-активированный K+ каналы". Нейробиолог. 9 (6): 434–9. Дои:10.1177/1073858403259282. PMID 14678575.
- ^ а б c Лю XK, Ван Г, Чен С.Д. (июнь 2010 г.). «Модуляция активности дофаминергических нейронов с помощью SK-каналов: потенциальная цель для лечения болезни Паркинсона?». Невролог Бык. 26 (3): 265–71. Дои:10.1007 / s12264-010-1217-4. ЧВК 5560298. PMID 20502506.
внешняя ссылка
- SK + калий + каналы в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- «Активированные кальцием калиевые каналы». База данных рецепторов и ионных каналов IUPHAR. Международный союз фундаментальной и клинической фармакологии.
- Джон Адельман. «Область научных интересов: кальций-активируемые калиевые каналы малой проводимости (SK-каналы)».. Орегонский университет здоровья и науки. Архивировано из оригинал на 2007-09-30. Получено 2008-01-22.