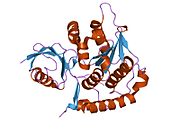
NEDD8 - NEDD8


NEDD8 это белок что у людей кодируется NEDD8 ген.[4][5] (В Saccharomyces cerevisiae этот белок известен как Руб1.) Этот убиквитиноподобный белок (ULP), становится ковалентно конъюгированным с ограниченным количеством клеточных белков аналогично убиквитинирование. NEDD8 человека имеет 60% идентичность аминокислотной последовательности убиквитину. Основными известными подложками модификации NEDD8 являются Каллин субъединицы убиквитинлигаз E3 на основе Cullin, которые активны только при неддилировании. Их NEDDylation является критическим для рекрутирования E2 в лигазный комплекс, тем самым облегчая конъюгацию убиквитина. Таким образом, модификация NEDD8 участвует в развитии клеточного цикла и регуляции цитоскелета.
Активация и конъюгация
Как и в случае с убиквитином и SUMO, NEDD8 конъюгируется с клеточными белками после того, как его C-концевой хвост обработан. В NEDD8, активирующий фермент E1 гетеродимер, состоящий из APPBP1 и субъединицы UBA3.[6] В APPBP1 / Фермент UBA3 гомологичен N- и C-концевой половинкам фермента убиквитина E1 соответственно. Субъединица UBA3 содержит каталитический центр и активирует NEDD8 в АТФ-зависимой реакции, образуя высокоэнергетический тиолэфирный промежуточный продукт. Активированный NEDD8 впоследствии переносится на фермент UbcH12 E2, а затем конъюгируется со специфическими субстратами в присутствии соответствующих лигаз E3.
Подложки для NEDD8
Согласно обзору Brown et al.,[7] наиболее охарактеризованными субстратами активированного NEDD8 являются Cullins (CUL1, 2, 3, 4A, 4B, 5 и 7 и PARC в клетках человека), которые служат в качестве молекулярных каркасов для cullin-ЗВЕНЕТЬ убиквитинлигазы (CRL). Неддилирование приводит к ковалентный конъюгирование фрагмента NEDD8 с консервированный Cullin лизин остаток.[8] Неддилирование куллина увеличивает активность убиквитилирования CRL за счет конформационных изменений, которые оптимизируют перенос убиквитина к целевым белкам
Удаление
Существует несколько различных протеаз, которые могут удалять NEDD8 из белковых конъюгатов. Протеазы UCHL1, UCHL3 и USP21 обладают двойной специфичностью к NEDD8 и убиквитину. Специфическими для удаления NEDD8 протеазами являются сигнаносома COP9, которая удаляет NEDD8 из субъединицы CUL1 убиквитинлигаз SCF, и NEDP1 (или DEN1, SENP8).[9]
Роль в восстановлении ДНК
Как показали Браун и др.,[7] Накопление NEDD8 в местах повреждения ДНК - очень динамичный процесс. Неддилирование необходимо в течение короткого периода суб-пути глобальной репарации генома (GGR) ДНК. эксцизионная репарация нуклеотидов (NER). В GGR NER после того, как повреждение ДНК вызвано УФ-облучением, Cul4A в белке, связывающем повреждение ДНК 2 (DDB2 ) комплекс активируется NEDD8, и это позволяет GGR-NER приступить к устранению повреждения.[10]
Неддилирование также играет роль в репарации двухцепочечных разрывов.[7] Негомологичное соединение концов (NHEJ) - это путь репарации ДНК, часто используемый для восстановления двунитевых разрывов ДНК. Первый шаг на этом пути зависит от Ku70 / Ku80 гетеродимер который образует высокостабильную кольцевую структуру, охватывающую концы ДНК.[11] Но гетеродимер Ku необходимо удалить, когда NHEJ завершится, иначе он блокирует транскрипцию или репликацию. Гетеродимер Ku убиквитилирован в зависимости от повреждения ДНК и неддилирования, чтобы способствовать высвобождению Ku и других факторов NHEJ из сайта репарации после завершения процесса.[7]
В химиотерапии рака
Как обсуждают Джин и Роберстон в своем обзоре,[12] подавление гена репарации ДНК гиперметилирование своего промоутер может быть очень ранним шагом к развитию рака. Предполагается, что молчание гена репарации ДНК на уровне транскрипции действует аналогично мутации зародышевой линии гена репарации ДНК. Потеря способности к репарации ДНК любым механизмом приводит к нестабильность генома и предрасполагает клетку и ее потомков к развитию рака. Эпигенетически гены репарации молчаливой ДНК часто встречаются при 17 наиболее распространенных формах рака (см. Частота гиперметилирования генов репарации ДНК при раке ).[12]
Как обсуждалось выше, активированный NEDD8 необходим в двух путях репарации ДНК: NER и NHEJ. Если активация NEDD8 ингибируется, клетки с индуцированным дефицитом NER или NHEJ могут погибнуть из-за недостаточной репарации ДНК, приводящей к накоплению повреждений ДНК. Эффект ингибирования NEDD8 может быть больше для раковых клеток, чем для нормальных клеток, если раковые клетки независимо испытывают дефицит репарации ДНК из-за предшествующего эпигенетического молчания генов репарации ДНК, активных в альтернативных путях (см. синтетическая летальность ).
Певонедистат (MLN4924), препарат, ингибирующий активацию NEDD8, продемонстрировал значительный терапевтический эффект в четырех Фаза I клинические исследования рака в 2015-2016 гг. К ним относятся испытания певонедистата в отношении острого миелоидного лейкоза и миелодиспластических синдромов,[13] рецидивирующая / рефрактерная множественная миелома или лимфома,[14] метастатическая меланома,[15] и запущенные солидные опухоли.[16]
В доклинических исследованиях
Неддилирование PPARγ
PPARγ играет решающую роль в адипогенез и накопление липидов в адипоциты (жировые клетки).[17] Активированный NEDD8 стабилизирует PPARγ, способствуя усилению адипогенеза. В экспериментах с мышами Певонедистат, препарат, ингибирующий активацию NEDD8, предотвращал ожирение и непереносимость глюкозы, вызванное диетой с высоким содержанием жиров.[17]
NF-κB и NEDD8
Транскрипционная активность NF-κB в первую очередь регулируется физическим взаимодействием с ингибирующими белками IκB (IκBα и IκBβ), что предотвращает его ядерную транслокацию.[18] Распад субъединицы IκBα в IκB опосредуется убиквитинированием, и это убиквитинирование зависит от неддилирования.[19] Певонедистат (MLN4924) ингибирует активацию NEDD8, который затем ингибирует убиквитинирование IκBα, и это ингибирует транслокацию NF-κB в ядро.[18]
Певонедистат, воздействуя на NF-κB и мишень для NF-κB (микроРНК-155), продлевал выживаемость мышей с привитыми лейкемическими клетками.[18]
Колоректальный рак
Было обнаружено, что ингибирование активации NEDD8 с помощью певонедистата вызывает задержку роста и апоптоз у 16/122 (13%) колоректальный рак (CRC) клеточные линии. Дальнейший анализ в ксенотрансплантаты опухоли, полученные от пациентов показали, что певонедистат эффективен в отношении низкодифференцированного муцинозного CRC высокой степени злокачественности. [20]
Взаимодействия
NEDD8 был показан взаимодействовать с:
Рекомендации
- ^ а б c ENSG00000285246 GRCh38: Ensembl release 89: ENSG00000129559, ENSG00000285246 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ «Ссылка на Mouse PubMed:». Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ Камитани Т., Кито К., Нгуен Х.П., Йе ET (ноябрь 1997 г.). «Характеристика NEDD8, убиквитин-подобного белка, подавляемого в процессе развития». Журнал биологической химии. 272 (45): 28557–62. Дои:10.1074 / jbc.272.45.28557. PMID 9353319.
- ^ «Ген Entrez: экспрессия нервных клеток-предшественников NEDD8, подавленная в процессе развития 8».
- ^ Walden H, Podgorski MS, Huang DT, Miller DW, Howard RJ, Minor DL, Holton JM, Schulman BA (декабрь 2003 г.). «Структура комплекса APPBP1-UBA3-NEDD8-ATP раскрывает основу для избирательной активации убиквитиноподобного белка с помощью E1». Молекулярная клетка. 12 (6): 1427–37. Дои:10.1016 / с1097-2765 (03) 00452-0. PMID 14690597.
- ^ а б c d Браун Дж. С., Лукащук Н., Сцанецка-Клифт М., Бриттон С., Ле Сейдж С., Калсу П., Бели П., Галанты Ю., Джексон С. П. (май 2015 г.). «Неддилирование способствует убиквитилированию и высвобождению Ku из участков повреждения ДНК». Отчеты по ячейкам. 11 (5): 704–14. Дои:10.1016 / j.celrep.2015.03.058. ЧВК 4431666. PMID 25921528.
- ^ Пан З. К., Кенцис А., Диас, округ Колумбия, Ямоа К., Ву К. (март 2004 г.). "Nedd8 на cullin: строительство скоростной дороги к разрушению белков". Онкоген. 23 (11): 1985–97. Дои:10.1038 / sj.onc.1207414. PMID 15021886.
- ^ "Обзор реагентов Boston Biochem NEDD8". Архивировано из оригинал на 2008-05-02. Получено 2008-04-29.
- ^ Гройсман Р., Полановска Дж., Кураока И., Савада Дж., Сайджо М., Драпкин Р., Киселев А.Ф., Танака К., Накатани Ю. (май 2003 г.). «Активность убиквитинлигазы в комплексах DDB2 и CSA по-разному регулируется сигнаносомой COP9 в ответ на повреждение ДНК». Клетка. 113 (3): 357–67. Дои:10.1016 / s0092-8674 (03) 00316-7. PMID 12732143.
- ^ Уокер JR, Corpina RA, Goldberg J (август 2001 г.). «Структура гетеродимера Ku, связанного с ДНК, и его значение для репарации двухцепочечных разрывов». Природа. 412 (6847): 607–14. Bibcode:2001 Натур 412..607Вт. Дои:10.1038/35088000. PMID 11493912.
- ^ а б Джин Б., Робертсон К.Д. (2013). ДНК-метилтрансферазы, восстановление повреждений ДНК и рак. Достижения экспериментальной медицины и биологии. 754. С. 3–29. Дои:10.1007/978-1-4419-9967-2_1. ISBN 978-1-4419-9966-5. ЧВК 3707278. PMID 22956494.
- ^ Swords RT, Erba HP, DeAngelo DJ, Bixby DL, Altman JK, Maris M, Hua Z, Blakemore SJ, Faessel H, Sedarati F, Dezube BJ, Giles FJ, Medeiros BC (май 2015 г.). «Певонедистат (MLN4924), первый в своем классе ингибитор фермента, активирующего NEDD8, у пациентов с острым миелоидным лейкозом и миелодиспластическими синдромами: исследование фазы 1» (PDF). Британский журнал гематологии. 169 (4): 534–43. Дои:10.1111 / bjh.13323. PMID 25733005.
- ^ Shah JJ, Jakubowiak AJ, O'Connor OA, Orlowski RZ, Harvey RD, Smith MR, Lebovic D, Diefenbach C, Kelly K, Hua Z, Berger AJ, Mulligan G, Faessel HM, Tirrell S, Dezube BJ, Lonial S ( Январь 2016 г.). «Фаза I исследования нового исследуемого ингибитора фермента, активирующего NEDD8, певонедистата (MLN4924) у пациентов с рецидивирующей / рефрактерной множественной миеломой или лимфомой». Клинические исследования рака. 22 (1): 34–43. Дои:10.1158 / 1078-0432.CCR-15-1237. ЧВК 5694347. PMID 26561559.
- ^ Бхатиа С., Павлик А.С., Боасберг П., Томпсон Дж. А., Маллиган Дж., Пикард М.Д., Фессель Н., Дезубе Б.Дж., Хамид О. (август 2016 г.). «Фаза I исследования исследуемого ингибитора NEDD8-активирующего фермента певонедистата (TAK-924 / MLN4924) у пациентов с метастатической меланомой». Новые исследуемые препараты. 34 (4): 439–49. Дои:10.1007 / s10637-016-0348-5. ЧВК 4919369. PMID 27056178.
- ^ Сарантопулос Дж., Шапиро Дж. И., Коэн РБ, Кларк Дж. У., Каух Дж. С., Вайс Дж. Дж., Клири Дж. М., Махалингам Д., Пикард, Мэриленд, Фессель Х. М., Бергер А. Дж., Берк К., Маллиган Дж., Дезубе Б. Дж., Харви РД (февраль 2016 г.). «Фаза I исследования исследуемого ингибитора фермента, активирующего NEDD8, певонедистата (TAK-924 / MLN4924) у пациентов с развитыми солидными опухолями». Клинические исследования рака. 22 (4): 847–57. Дои:10.1158 / 1078-0432.CCR-15-1338. PMID 26423795.
- ^ а б Park HS, Ju UI, Park JW, Song JY, Shin DH, Lee KH, Jeong LS, Yu J, Lee HW, Cho JY, Kim SY, Kim SW, Kim JB, Park KS, Chun YS (август 2016 г.). «Неддилирование PPARγ, необходимое для адипогенеза, является потенциальной мишенью для лечения ожирения». Гибель клеток и дифференциация. 23 (8): 1296–311. Дои:10.1038 / cdd.2016.6. ЧВК 4947677. PMID 26990658.
- ^ а б c Khalife J, Radomska HS, Santhanam R, Huang X, Neviani P, Saultz J, Wang H, Wu YZ, Alachkar H, Anghelina M, Dorrance A, Curfman J, Bloomfield CD, Medeiros BC, Perrotti D, Lee LJ, Lee RJ , Калиджури М.А., Пичиорри Ф., Кроче С.М., Гарсон Р., Гусман М.Л., Мендлер Дж. Х., Маркучи Дж. (Октябрь 2015 г.). «Фармакологическое воздействие на miR-155 с помощью ингибитора NEDD8-активирующего фермента MLN4924 (Pevonedistat) при остром миелоидном лейкозе FLT3-ITD». Лейкемия. 29 (10): 1981–92. Дои:10.1038 / leu.2015.106. ЧВК 4868182. PMID 25971362.
- ^ Фролова М.А., Гудкова Р.Г., Большухина Л.А., Новоселова В.Н. (октябрь 1978 г.). «[Феномен усиления у мышей с гетеротопической трансплантацией сердца]». Журнал Микробиологии, Эпидемиологии, И Иммунобиологии. 20 (10): 36–40. PMID 85397.
- ^ Picco G, Petti C, Sassi F, Grillone K, Migliardi G, Rossi T, Isella C, Di Nicolantonio F, Sarotto I, Sapino A, Bardelli A, Trusolino L, Bertotti A, Medico E (февраль 2017 г.). «Эффективность ингибирования пути NEDD8 в доклинических моделях плохо дифференцированного, клинически агрессивного колоректального рака». Журнал Национального института рака. 109 (2): djw209. Дои:10.1093 / jnci / djw209. PMID 27771609.
- ^ Антенос М., Каспер РФ, Браун Т.Дж. (ноябрь 2002 г.). «Взаимодействие с Nedd8, убиквитин-подобным белком, усиливает транскрипционную активность арилуглеводородного рецептора». Журнал биологической химии. 277 (46): 44028–34. Дои:10.1074 / jbc.M202413200. PMID 12215427.
- ^ Hipp MS, Raasi S, Groettrup M, Schmidtke G (апрель 2004 г.). «NEDD8 ultimate buster-1L взаимодействует с убиквитин-подобным белком FAT10 и ускоряет его распад». Журнал биологической химии. 279 (16): 16503–10. Дои:10.1074 / jbc.M310114200. PMID 14757770.
- ^ Камитани Т., Кито К., Фукуда-Камитани Т., Йе Э.Т. (декабрь 2001 г.). «Нацеливание NEDD8 и его конъюгатов на протеасомную деградацию с помощью NUB1». Журнал биологической химии. 276 (49): 46655–60. Дои:10.1074 / jbc.M108636200. PMID 11585840.
- ^ а б Gong L, Yeh ET (апрель 1999 г.). «Идентификация активирующих и конъюгирующих ферментов пути конъюгации NEDD8». Журнал биологической химии. 274 (17): 12036–42. Дои:10.1074 / jbc.274.17.12036. PMID 10207026.
- ^ Вада Х, Кито К., Каски Л.С., Йе Е.Т., Камитани Т. (октябрь 1998 г.). «Расщепление C-конца NEDD8 посредством UCH-L3». Сообщения о биохимических и биофизических исследованиях. 251 (3): 688–92. Дои:10.1006 / bbrc.1998.9532. PMID 9790970.
дальнейшее чтение
- Xirodimas DP (октябрь 2008 г.). «Новые субстраты и функции для убиквитиноподобной молекулы NEDD8». Сделки Биохимического Общества. 36 (Часть 5): 802–6. Дои:10.1042 / BST0360802. PMID 18793140.
- Кумар С., Томока Ю., Нода М. (июнь 1992 г.). «Идентификация набора генов с подавленной экспрессией в мозге мышей». Сообщения о биохимических и биофизических исследованиях. 185 (3): 1155–61. Дои:10.1016 / 0006-291X (92) 91747-E. PMID 1378265.
- Кумар С., Йошида Ю., Нода М. (август 1993 г.). «Клонирование кДНК, которая кодирует новый убиквитиноподобный белок». Сообщения о биохимических и биофизических исследованиях. 195 (1): 393–9. Дои:10.1006 / bbrc.1993.2056. PMID 8395831.
- Боналдо М.Ф., Леннон Г., Соарес МБ (сентябрь 1996 г.). «Нормализация и вычитание: два подхода для облегчения открытия генов». Геномные исследования. 6 (9): 791–806. Дои:10.1101 / гр.6.9.791. PMID 8889548.
- Осака Ф., Кавасаки Х., Аида Н., Саеки М., Чиба Т., Кавасима С., Танака К., Като С. (август 1998 г.). «Новая система лигирования NEDD8 для cullin-4A». Гены и развитие. 12 (15): 2263–8. Дои:10.1101 / gad.12.15.2263. ЧВК 317039. PMID 9694792.
- Вада Х, Кито К., Каски Л.С., Йе Е.Т., Камитани Т. (октябрь 1998 г.). «Расщепление C-конца NEDD8 посредством UCH-L3». Сообщения о биохимических и биофизических исследованиях. 251 (3): 688–92. Дои:10.1006 / bbrc.1998.9532. PMID 9790970.
- Whitby FG, Xia G, Pickart CM, Hill CP (декабрь 1998 г.). «Кристаллическая структура человеческого убиквитиноподобного белка NEDD8 и взаимодействия с ферментами пути убиквитина». Журнал биологической химии. 273 (52): 34983–91. Дои:10.1074 / jbc.273.52.34983. PMID 9857030.
- Gong L, Yeh ET (апрель 1999 г.). «Идентификация активирующих и конъюгирующих ферментов пути конъюгации NEDD8». Журнал биологической химии. 274 (17): 12036–42. Дои:10.1074 / jbc.274.17.12036. PMID 10207026.
- Гонг Л., Ли Б., Миллас С., Йе Э. Т. (апрель 1999 г.). «Молекулярное клонирование и характеристика человеческих AOS1 и UBA2, компонентов комплекса ферментов, активирующих сентрин». Письма FEBS. 448 (1): 185–9. Дои:10.1016 / S0014-5793 (99) 00367-1. PMID 10217437.
- Лиакопулос Д., Бюсген Т., Брихзи А., Йенч С., Пауза А. (май 1999 г.). «Конъюгирование убиквитиноподобного белка NEDD8 с кулином-2 связано с функцией супрессора опухоли фон Хиппеля-Линдау». Труды Национальной академии наук Соединенных Штатов Америки. 96 (10): 5510–5. Bibcode:1999ПНАС ... 96.5510Л. Дои:10.1073 / пнас.96.10.5510. ЧВК 21890. PMID 10318914.
- Певец JD, Gurian-West M, Clurman B, Робертс JM (сентябрь 1999 г.). «Cullin-3 нацелен на циклин E для убиквитинирования и контролирует S-фазу в клетках млекопитающих». Гены и развитие. 13 (18): 2375–87. Дои:10.1101 / gad.13.18.2375. ЧВК 317026. PMID 10500095.
- Симеони С., Манчини М.А., Стенойен Д.Л., Марчелли М., Вейгель Н.Л., Заниси М., Мартини Л., Полетти А. (январь 2000 г.). «Гибель мотонейрональных клеток не коррелирует с образованием агрегатов андрогенных рецепторов, содержащих удлиненный полиглутаминовый тракт» (PDF). Молекулярная генетика человека. 9 (1): 133–44. Дои:10.1093 / hmg / 9.1.133. PMID 10587588.
- Хори Т., Осака Ф., Чиба Т., Миямото К., Окабаяси К., Шимбара Н., Като С., Танака К. (ноябрь 1999 г.). «Ковалентная модификация всех членов белков семейства кулинов человека с помощью NEDD8». Онкоген. 18 (48): 6829–34. Дои:10.1038 / sj.onc.1203093. PMID 10597293.
- Рид М.А., Браунелл Дж. Э., Гладышева Т. Б., Хоттелет М., Родитель Л. А., Коггинс М. Б., Пирс Дж. В., Подуст В. Н., Луо Р. С., Чау В., Паломбелла В. Дж. (Апрель 2000 г.). «Nedd8-модификация cul-1 активирует SCF (бета (TrCP)) - зависимое убиквитинирование IkappaBalpha». Молекулярная и клеточная биология. 20 (7): 2326–33. Дои:10.1128 / MCB.20.7.2326-2333.2000. ЧВК 85397. PMID 10713156.
- Моримото М., Нисида Т., Хонда Р., Ясуда Х. (апрель 2000 г.). «Модификация cullin-1 убиквитин-подобным белком Nedd8 усиливает активность SCF (skp2) по отношению к p27 (kip1)». Сообщения о биохимических и биофизических исследованиях. 270 (3): 1093–6. Дои:10.1006 / bbrc.2000.2576. PMID 10772955.
- Wada H, Yeh ET, Kamitani T. (июнь 2000 г.). «Доминантно-отрицательный мутант UBC12 изолирует NEDD8 и ингибирует конъюгацию NEDD8 in vivo». Журнал биологической химии. 275 (22): 17008–15. Дои:10.1074 / jbc.275.22.17008. PMID 10828074.
- Кито К., Йе Е.Т., Камитани Т. (июнь 2001 г.). «NUB1, белок, взаимодействующий с NEDD8, индуцируется интерфероном и подавляет экспрессию NEDD8». Журнал биологической химии. 276 (23): 20603–9. Дои:10.1074 / jbc.M100920200. PMID 11259415.
- Камитани Т., Кито К., Фукуда-Камитани Т., Йе Э.Т. (декабрь 2001 г.). «Нацеливание NEDD8 и его конъюгатов на протеасомную деградацию с помощью NUB1». Журнал биологической химии. 276 (49): 46655–60. Дои:10.1074 / jbc.M108636200. PMID 11585840.
- Ву К., Чен А., Тан П., Пан З. К. (январь 2002 г.) «Конъюгированная с Nedd8 коровая убиквитинлигаза ROC1-CUL1 использует заряженные поверхностные остатки Nedd8 для эффективной сборки полиубиквитиновой цепи, катализируемой Cdc34». Журнал биологической химии. 277 (1): 516–27. Дои:10.1074 / jbc.M108008200. PMID 11675391.
PDB галерея | |
|---|---|
|