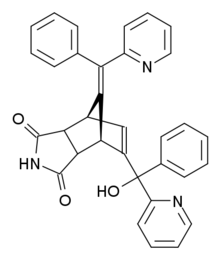
Норбормид - Norbormide
 | |
| Имена | |
|---|---|
| Предпочтительное название IUPAC 5- (α-Гидрокси-α-2-пиридилбензил) -7- (α-2-пиридилбензилиден) -5-норборнен-2,3-дикарбоксимид | |
| Систематическое название ИЮПАК (10E) -8- [гидрокси (фенил) пиридин-2-илметил] -10- [фенил (пиридин-2-ил) метилиден] -4-азатрицикло [5.2.1.02,6] дек-8-ен-3,5-дион | |
| Идентификаторы | |
3D модель (JSmol ) | |
| ChemSpider | |
| ECHA InfoCard | 100.012.354 |
| Номер ЕС |
|
| КЕГГ | |
PubChem CID | |
| UNII | |
| |
| |
| Характеристики | |
| C33ЧАС25N3О3 | |
| Молярная масса | 511.570 |
| Опасности | |
| Главный опасности | Токсичный |
Если не указано иное, данные для материалов приведены в их стандартное состояние (при 25 ° C [77 ° F], 100 кПа). | |
| Ссылки на инфобоксы | |
Норбормид (Raticate, Шоксин) - токсичное соединение, используемое в качестве родентицид. Он имеет несколько механизмов действия, действуя как сосудосуживающее средство и блокатор кальциевых каналов,[1] но избирательно токсичен для крысы и имеет относительно низкую токсичность для других видов из-за видоспецифического действия открытия проницаемость переходных пор в крысе митохондрии.[2]
Классифицируется как чрезвычайно опасное вещество в Соединенных Штатах, как определено в Разделе 302 США Закон о чрезвычайном планировании и праве общества на информацию (42 U.S.C. 11002), и к ним предъявляются строгие требования к отчетности со стороны предприятий, которые производят, хранят или используют его в значительных количествах.[3]
История
В начале 1960-х годов был разработан норбормид в качестве неантикоагулянтного крысиного яда. Однако в течение 1970-х годов использование этого родентицида снизилось, поскольку антикоагулянтные токсины оказались более эффективными против более широкого круга грызунов.[4] NRB убивает только грызунов из рода Rattus (R. norvegicus, R. exulans и R. rattus) и оказывается умеренно безвредным для других грызунов и млекопитающих.[5] Хотя многие считают его селективность недостатком, ученые Landcare Research в Новой Зеландии ищут способы улучшить этот родентицид и разрабатывают несколько аналогов.[6][7][8]
Структура и реакционная способность
Норбормид представляет собой органическое соединение со следующим систематическим названием: 5- (α-гидрокси-α-2-пиридилбензил) -7- (α-2-пиридилбензилиден) -5-норборнен-2,3-дикарбоксимид. Структура состоит из норборненового кольца, которое слито с имидным кольцом напротив двойной связи. Один из углеродных атомов этой двойной связи связан с другим углеродным атомом, который связан с гидроксильной, пиридильной и фенильной группами. Мостиковый углерод норборненового кольца связан двойной связью с атомом углерода, к которому присоединены пиридильная и фенильная группы.[9] Существует восемь возможных стереоизомеров норбормида. По экзоциклической двойной связи существует цис / транс-изомерия. Имидное кольцо может иметь эндо- или экзо-ориентацию, а для гидроксильной группы возможны эритро- и трео-изомеры.[10] Сосудосуживающие свойства норбормида оказались очень зависимыми от стереохимии. Только эндо-изомеры токсичны для крыс, а трео-изомеры в десять раз сильнее, чем эритро-изомеры.[11] Было обнаружено, что изомер цис-эндо-трео является изомером с наиболее сильными вазоконстрикторными свойствами.[12] В этой структуре существует водородная связь между гидроксильной группой и соседним пиридиновым кольцом.[5] Исследования показывают, что токсичность норбормида чувствительна к структурным изменениям, почти во всех случаях токсичность снижается из-за структурных изменений. Только замена протона NH имида определенными группами может дать токсическую активность, сравнимую с самим норбормидом.[13]
Синтез
Поскольку NRB вызывает у крыс и крыс робость приманки, поэтому часто принимают только сублетальные дозы, были проведены исследования по поиску производных NRB, которые более токсичны, чем сам NRB. В этом случае замены произошли только в имидной группе NRB. Общая структура для каждой из этих производных показана на рисунке 4.1, где R - изменяемая группа.
В положение R были помещены различные углеводородные группы. Ни один из них не был токсичнее NRB, поэтому была опробована другая стратегия. Во втором исследовании были изучены различные кольцевые аналоги NRB. Ни один из этих аналогов кольца не оказался более токсичным, чем NRB.[13]Другой тип реакции, который был изучен, - это производство пролекарств. Эти пролекарства были синтезированы с целью преодоления «застенчивости» приманки. Целью исследования было создание пролекарства с лучшим вкусом, чем NRB, которое после попадания в организм метаболизировалось до NRB. Были использованы три различные стартовые конструкции, показанные на рисунках 4.2, 4.3 и 4.4. На рисунках 4.5 и 4.6 показано, как исходные структуры были синтезированы из NRB. Только соединение 19 (рис. 4.7) было многообещающим, потому что оно задерживало появление симптомов и более приемлемо для крыс (показано на рис. 4.8), но необходимо провести дополнительные исследования этого соединения, прежде чем его можно будет использовать.[13]
Доступные формы
Различные стереоизомеры
Во время синтеза норбормида пять из восьми возможных стереоизомеров образуются в значительном количестве, а именно все эндостереоизомеры и цис-экзо-стереоизомер. Большая часть эффективности норбормида обусловлена транс-эндо-трео (LD50 = 0,50 мг / кг (крыса)) и цис-эндо-трео-изомерами (LD50 = 0,15 мг / кг (крыса)). Эти два изомера составляют примерно половину коммерческой смеси.[13]
Производные
Были проведены исследования, в которых проводился поиск более токсичных производных соединений норбормида. Добавление или замена разных групп никогда не приводило к получению значительно более токсичных соединений. В большинстве случаев были получены соединения, которые были значительно менее токсичны.[13]Проблемой использования норбормида в качестве родентицида является застенчивость приманки, это означает, что после того, как крыса съест немного, крыса заболеет и затем избегает токсина, также предполагается, что вкус будет плохим. Недавние исследования были направлены на поиск пролекарств норбормида, которые медленно высвобождают токсикант и тем самым замедляют токсические эффекты. Были обнаружены пролекарства, которые, по-видимому, обладают этими свойствами. Последующие исследования необходимо провести для уточнения, прежде чем в конечном итоге станет возможным использование.[11]
Механизм действия
Предполагается, что механизм сосудосуживающего эффекта опосредуется модуляцией притока кальция. Этот приток кальция может привести к сокращению миоцитов. Вероятно, приток кальция опосредуется рецепторами, связанными с фосфолипазой C (PLC), в миоцитах периферических артерий крыс.[14][15]
Другое исследование Серджио Бова и др. показал, что в гладких мышцах дыхательных путей, мочевыводящих путей и желудочно-кишечного тракта не было сокращения норбормидом. Симптомы норбормида были очень похожи на симптомы более известных блокаторов входа Ca2 +. Следовательно, норбормид не только видоспецифичен, но и тканеспецифичен.[16]
Норбормид оказывает сильное влияние на митохондрии в клетке. Следовательно, норбормид переносится через внешнюю митохондриальную мембрану (OMM) во внутреннее митохондриальное пространство. В этом месте или в матрице он вызывает поры перехода проницаемости (PTP). Этот PTP представляет собой канал внутренней митохондриальной мембраны (IMM), открытие которого приводит к увеличению проницаемости для ионов с размером исключения около 1500 Да.[17] Транспорт норборимида происходит за счет белка транслокации (TSPO), также известного как периферический бензодиазепиновый рецептор. TSPO является селективным по отношению к транспорту норбормида у крыс. На рисунке 6.1 показано ключевое выравнивание LPSO крысы, мыши и морской свинки. Есть несколько различий в аминокислотах, но позиция 113 очень похожа между видами и другими видами, такими как собаки, люди и куры. Если крыса имеет в положении 113 метионин (M), у других видов есть аминокислота лейцин. Вероятно, это происходит из-за различного количества транспорта между крысами и другими видами.[17]
Метаболизм
Токсикологические свойства соединения связаны с метаболическими путями, которые часто различаются у разных видов. По этой причине наблюдалась корреляция между токсичностью и метаболизмом, чтобы получить четкое представление о клеточном метаболизме как в условиях in vitro, так и in vivo.[18][19][20]
In vitro эксперимент
Исследования in vitro препаратов печени крыс и других грызунов показали, что гидроксилирование является основным процессом метаболизма NRB. Более того, метаболиты у крыс разных полов, по-видимому, отличались друг от друга.[5] Прежде всего, когда NRB инкубировали с фракцией S9 печени, было обнаружено несколько продуктов метаболизма. Фракция S9 была определена как «фракция супернатанта, полученная из гомогената органа (обычно печени) центрифугированием при 9000 g в течение 20 минут в подходящей среде; эта фракция содержит цитозоль и микросомы».[21] Фракция S9 состоит из двух компонентов: микросомного компонента, который включает изоформы цитохрома P450 (фаза метаболизма I).[22] и цитозольный компонент, содержащий трансферазы (метаболизм фазы II).[23]Смесь четырех активных эндо-изомеров NRB (U, V, W en Y) образовала четыре основных метаболита в S9 печени крысы и цитозольных препаратах после инкубации. Изомеры по отдельности не являются чистыми соединениями, но исследования с чистым изомером V и смесью изомеров U и V показали, что каждый изомер подвергается метаболизму с образованием у крысы одного продукта. Масс-спектрометрия подтвердила, что все метаболиты имеют массу, соответствующую гидроксилированному метаболиту NRB. Поскольку самки крыс более чувствительны к NRB, чем самцы, уровень метаболитов у самок выше, чем у самцов.[5]Те же метаболиты были обнаружены у морских свинок, хотя уровни метаболизма у этих грызунов значительно ниже, чем у крыс. У мышей были обнаружены два метаболита: один метаболит V-изомера (V1), который также был обнаружен в препаратах крыс, и другой, новый метаболит V-изомера (V2). (Рисунок 7.1) Эти данные подтверждают связь между чувствительностью к NRB и производством различных типов и различных уровней метаболитов.
In vivo эксперимент
После сбора крови у крыс через 10 и 30 минут после перорального введения фракции плазмы и эритроцитов анализировали в поисках либо исходных соединений, либо гидроксилированных метаболитов. Однако ни того, ни другого не было обнаружено, что либо означает, что абсорбция не происходит, либо соединения быстро выводятся из организма. Образцы цельной крови определенно показали следы исходного соединения и метаболита (M3) с молекулярной массой 226 в образцах как самок, так и самцов крыс. Уровень, в котором присутствует этот метаболит, снова оказался выше у самок крыс, чем у самцов. M3 не был обнаружен ни в каких других тканях крыс или мышей. Это говорит о том, что M3 может образовываться микрофлорой кишечника у крыс.[24]После перорального введения NRB исследование печени крысы и мыши выявило следы исходного соединения. Сравнивая уровни метаболитов у обоих полов и видов, у самок крыс было обнаружено значительно большее количество, чем у самцов крыс или мышей. Хроматограммы ВЭЖХ препаратов печени самок и самцов крыс показывают обнаружение четырех метаболитов (U1, V1, M1, M2). (Рисунок 7.2) Для их идентификации использовалась масс-спектрометрия, и это было легко, потому что гидроксилированные метаболиты были на +16 а.е.м. выше, чем NRB. Эти исследования подтвердили, что метаболиты идентичны тем, которые обнаруживаются в препаратах in vitro.[5][24]В препаратах печени мышей метаболиты не обнаружены. Более того, ни исходное соединение NRB, ни его метаболиты не были обнаружены в сердце после перорального введения крысам, а также мышам. Отсутствие обнаруживаемых метаболитов означает, что NRB способен вызывать летальный эффект при чрезвычайно низких уровнях.[24]
Побочные эффекты и воздействие на животных
Побочные эффекты
Эффекты норбормида вызваны эндо-изомерами, но экзо-изомеры R, T и X не показали сократительного действия на гладкие мышцы артериальных сосудов крысы. Было только расслабляющее действие на мышцы.[5] В исследованиях Розковски на животных он показал, что эффекты, вызываемые NRB, необратимы.[25] Другое исследование показало, что у крыс, получавших норбормид in vivo, наблюдались угнетения дыхания после сердечно-сосудистых эффектов.[26]
Воздействие на животных
NRB особенно токсичен для крыс, но относительно безвреден для других грызунов и млекопитающих. У всех протестированных животных, а также в аорте крысы и внесосудистой гладкой мышечной ткани NRB проявляет сосудорасширяющие свойства в артериях.[14]Другой эффект NRB - стимуляция выработки кортикостерона и альдостерона в надпочечниках крыс и мышей за счет усиления поздних стадий синтеза стероидных гормонов.[15]
Токсичность
Уникальная токсичность NRB была определена путем проведения экспериментов на животных с использованием нескольких видов грызунов. Токсическая доза вводилась перорально.[27] В таблице определенные данные LD50 иллюстрируют специфическую токсичность соединения. Крысы, особенно самки, даже после введения небольшого количества NRB испытывают действие токсина. NRB токсичен также для морских свинок и мышей, но в меньшей степени. Достоверной информации о значениях LD50 у человека нет. Хотя NRB является родентицидом и особенно токсичен для коричневых крыс, люди могут подвергаться воздействию NRB при вдыхании и контакте с кожей.
Воздействие Уровень острой токсичностиПероральный LD50 (мг / кг)
- Крыса (самец) 15
- Крыса (самка) 5
- Морская свинка 620
- Мышь 2250
Исследования острой токсичности на крысах
Смертельная доза NRB для крыс вызывает изменения в поведении, которые очень похожи на признаки, связанные с токсичностью цианида. Первые признаки токсичности NRB проявляются примерно через десять минут после приема. Сначала у крыс появляются признаки повышенной двигательной активности и нарушения координации движений. В дальнейшем задние конечности крыс ослабевают и становятся пепельными. Их дыхание становится затрудненным, и через короткий промежуток времени крысы начинают страдать от серии судорожных движений. За спазмом следует смерть, которая наступает в течение 30 минут у лабораторных крыс-альбиносов и в течение двух часов у диких животных.[25]После исследования изолированных сердец крыс в препаратах Лангендорфа, часто используемом в лабораторных условиях, NRB также оказывал сильное токсическое воздействие на сердце крысы.[25] В коронарный сосуд вводили 2-10 мкг NRB, что приводило к уменьшению кровотока, сопровождавшемуся уменьшением сердечных сокращений и уменьшением частоты сердечных сокращений. Крысиные сердца начали биться нерегулярно. Эти необратимые эффекты были отмечены не в сердечной мышце (миокарде), как, возможно, ожидалось, а в коронарных артериях.[25] Кроме того, было подтверждено, что крысы, которые подвергались лечению NRB, страдали гипервентиляцией с последующими сердечно-сосудистыми эффектами.[26]Недавние исследования показывают способность NRB активировать поры перехода митохондриальной проницаемости (MPTP) в изолированных препаратах крыс. Апоптоз и некроз клеток (среди прочего) регулируются порами MPT. Понятно, что такая аномалия митохондриальных мембран вызывает проблемы в клеточном метаболизме отравленного животного.[8]
Исследования токсичности у млекопитающих и птиц
Представители рода Rattus очень чувствительны к NRB, но другие животные не испытывают токсических эффектов. При более внимательном рассмотрении доза норбормида 1000 мг / кг не выявила никаких токсических признаков при пероральном введении кошкам, цыплятам, собакам, обезьянам, мышам, свиньям или птицам. Это подтверждено в таблице ниже. Таким образом, можно с уверенностью сказать, что NRB относительно безвреден для видов, не являющихся крысами.[25]
Рекомендации
- ^ Реннисон Д., Бова С., Кавалли М., Ричелли Ф., Зулиан А., Хопкинс Б., Бримбл М.А. (апрель 2007 г.). «Синтез и изучение активности аналогов селективного токсиканта норбормида крыс». Биоорганическая и медицинская химия. 15 (8): 2963–74. Дои:10.1016 / j.bmc.2007.02.012. PMID 17321141.
- ^ Зулиан А., Петронилли В., Бова С., Даббени-Сала Ф., Карнелли Г., Кавалли М. и др. (Июль 2007 г.). «Оценка молекулярной основы для крыс-селективной индукции перехода митохондриальной проницаемости норбормидом». Biochimica et Biophysica Acta (BBA) - Биоэнергетика. 1767 (7): 980–8. Дои:10.1016 / j.bbabio.2007.04.002. PMID 17509521.
- ^ «40 C.F.R .: Приложение A к Части 355 - Список особо опасных веществ и их планируемые пороговые количества» (PDF) (1 июля 2008 г. ред.). Государственная типография. Архивировано из оригинал (PDF) 25 февраля 2012 г.. Получено 29 октября, 2011.
- ^ Телле HJ (1967). «Некоторые полевые наблюдения по наблюдениям за приемлемостью и эффективностью норбормида для Rattus norvegicus». Кто / VBC. 67 (39): 10.
- ^ а б c d е ж Равиндран С., Хопкинс Б., Бова С., Реннисон Д., Бримл М., Тингл М. (январь 2009 г.). «Метаболизм норбормида in vitro в препаратах печени крыс, мышей и морских свинок». Экологическая токсикология и фармакология. 27 (1): 144–8. Дои:10.1016 / j.etap.2008.09.007. PMID 21783932.
- ^ Бова С., Тревизи Л., Чима Л., Лучиани С., Головина В., Карнелли Г. (февраль 2001 г.). «Сигнальные механизмы селективного сосудосуживающего действия норбормида на мелкие артерии крыс». Журнал фармакологии и экспериментальной терапии. 296 (2): 458–63. PMID 11160631.
- ^ Кавалли М., Омичиуоло Л., Карнелли Дж., Чима Л., Хопкинс Б., Бова С. (сентябрь 2004 г.). «Распределение сосудосуживающего и сосудорасширяющего действия норбормида вдоль сосудистого дерева крысы». Науки о жизни. 75 (18): 2157–65. Дои:10.1016 / j.lfs.2004.04.022. PMID 15325842.
- ^ а б Ричелли Ф., Даббени-Сала Ф., Петронилли В., Бернарди П., Хопкинс Б., Бова С. (июнь 2005 г.). «Видоспецифическая модуляция перехода митохондриальной проницаемости норбормидом». Biochimica et Biophysica Acta (BBA) - Биоэнергетика. 1708 (2): 178–86. Дои:10.1016 / j.bbabio.2005.03.002. PMID 15953474.
- ^ Нильссон Б. Стерохимия неактивного рацемата норбормида - селективного токсиканта для крыс. Acta Chemica Scandinavica, 1968. 22 (2).
- ^ Steel PJ, Brimble MA, Hopkins B, Rennison D (май 2004 г.). «Два стереоизомера крысиного токсиканта норбормида». Acta Crystallographica Раздел C. 60 (Пт 5): o374-6. Дои:10.1107 / S0108270104006845. HDL:10092/365. PMID 15131397.
- ^ а б Реннисон Д., Бова С., Кавалли М., Ричелли Ф., Зулиан А., Хопкинс Б., Бримбл М.А. (апрель 2007 г.). «Синтез и изучение активности аналогов селективного токсиканта норбормида крыс». Биоорганическая и медицинская химия. 15 (8): 2963–74. Дои:10.1016 / j.bmc.2007.02.012. PMID 17321141.
- ^ Бримл М.А., э.а., 2004.
- ^ а б c d е Поос Г.И., Морбахер Р.Дж., Карсон Э.Л., Парагамиан В., Пума Б.М., Расмуссен С.Р., Рошковски А.П. (июль 1966 г.). «Исследования структуры-активности с селективным токсикантом для крыс норбормидом». Журнал медицинской химии. 9 (4): 537–40. Дои:10.1021 / jm00322a021. PMID 5968018.
- ^ а б Реннисон Д., Лайта О, Бова С., Кавалли М., Хопкинс Б., Линтикум Д.С., Бримбл М.А. (июль 2012 г.). «Дизайн и синтез пролекарств селективного токсиканта норбормида для крыс». Биоорганическая и медицинская химия. 20 (13): 3997–4011. Дои:10.1016 / j.bmc.2012.05.014. PMID 22658693.
- ^ а б Нери Дж., Торторелла С., Андреис П. Г., Бова С., Малендович Л. К., Циолковска А., Нуссдорфер Г. Г. (март 2003 г.). «Норбормид усиливает поздние стадии синтеза стероидных гормонов в коре надпочечников крыс и мышей». Журнал стероидной биохимии и молекулярной биологии. 84 (4): 479–83. Дои:10.1016 / s0960-0760 (03) 00060-8. PMID 12732293. S2CID 10840313.
- ^ Bova S, Cavalli M, Cima L, Luciani S, Saponara S, Sgaragli G, Cargnelli G, Fusi F (июнь 2003 г.). «Расслабляющие и блокирующие Ca2 + каналы свойства норбормида на несосудистых гладких мышцах крыс». Европейский журнал фармакологии. 470 (3): 185–91. Дои:10.1016 / s0014-2999 (03) 01797-7. PMID 12798957.
- ^ а б Зулиан А., Силейките Дж., Петронилли В., Бова С., Даббени-Сала Ф., Карнелли Дж., Реннисон Д., Бримбл М.А., Хопкинс Б., Бернарди П., Ричелли Ф. (декабрь 2011 г.). «Белок-транслокатор (периферический бензодиазепиновый рецептор) опосредует селективную для крыс активацию перехода митохондриальной проницаемости норбормидом». Biochimica et Biophysica Acta (BBA) - Биоэнергетика. 1807 (12): 1600–5. Дои:10.1016 / j.bbabio.2011.08.007. PMID 21889488.
- ^ Митчелл-младший, Jollow DJ, Поттер WZ, Дэвис, округ Колумбия, Gillette JR, Броди BB (октябрь 1973). «Ацетаминофен-индуцированный некроз печени. I. Роль метаболизма лекарств». Журнал фармакологии и экспериментальной терапии. 187 (1): 185–94. PMID 4746326.
- ^ Хендерсон РФ (декабрь 1996 г.). «Видовые различия в метаболизме бензола». Перспективы гигиены окружающей среды. 104 Дополнение 6: 1173–5. Дои:10.1289 / ehp.961041173. ЧВК 1469720. PMID 9118889.
- ^ Тингл, доктор медицины, Махмуд Р., Маггс Дж. Л., Пирмохамед М., Парк Б.К. (ноябрь 1997 г.). «Сравнение метаболизма и токсичности дапсона у крыс, мышей и человека». Журнал фармакологии и экспериментальной терапии. 283 (2): 817–23. PMID 9353403.
- ^ Даффус Дж. Х., Нордберг М., Темплтон Д. М. (январь 2007 г.). «Глоссарий терминов, используемых в токсикологии, 2-е издание». Чистая и прикладная химия. 79 (7): 1153–344. Дои:10.1351 / pac200779071153. S2CID 98296965.
- ^ Грейм Х, Снайдер Р (2008). Токсикология и оценка рисков: всестороннее введение. Wiley-Interscience. п. 387.
- ^ Фогель GH (2006). Открытие и оценка лекарств: безопасность и фармакокинетические анализы. Springer. п. 509.
- ^ а б c Равиндран С., Хопкинс Б., Бова С., Тингл М. (июль 2009 г.). «Метаболизм норбормида in vivo у крыс и мышей». Экологическая токсикология и фармакология. 28 (1): 147–51. Дои:10.1016 / j.etap.2009.03.013. PMID 21783995.
- ^ а б c d е Рошковский А.П. (август 1965 г.). «Фармакологические свойства норбормида, селективного токсиканта для крыс». Журнал фармакологии и экспериментальной терапии. 149 (2): 288–99. PMID 4953462.
- ^ а б Ельноски Дж., Лоулор Р. (сентябрь 1971 г.). «Сердечно-сосудистые эффекты норбормида». Европейский журнал фармакологии. 16 (1): 117–9. Дои:10.1016/0014-2999(71)90065-3. PMID 5157526.
- ^ Рассел RU. «Норбормид - токсичный агент, специфичный для Rattus». J. Forensic Sci. 1965: 80–83.