Острохвостый тетерев - Sharp-tailed grouse
| Острохвостый тетерев | |
|---|---|
 | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Авес |
| Заказ: | Galliformes |
| Семья: | Phasianidae |
| Род: | Тимпанух |
| Разновидность: | Т. phasianellus |
| Биномиальное имя | |
| Tympanuchus phasianellus | |
 | |
| Синонимы | |
| |

В острохвостый тетерев (Tympanuchus phasianellus) (ранее: Tetrao phasianellus) - это прерия среднего размера тетерев. Он также известен как острый хвост или же тетерев.
Таксономия
В курица из больших прерий, цыпленок малой прерии, а куропатки составляют род Тимпанух, род тетерева, встречающийся только в Северная Америка. Полное научное название острохвостого тетерева - Tympanuchus phasianellus. Описаны шесть существующих и один вымерший подвид острохвостого тетерева:[2]
- Т. П. фазианелл: номинирующая раса или северный острохвостый тетерев встречается в Манитоба, северный Онтарио, и центральный Квебек. Частично мигрирующий.
- Т. П. Kennicotti: северо-западный острохвостый тетерев обитает в Mackenzie River к Большое Невольничье озеро в Северо-Западные территории, Канада.
- Т. П. Caurus: аляскинский острохвостый тетерев населяет северо-центральный регион. Аляска на восток на юг Юкон, северный британская Колумбия, и северный Альберта.
- Т. П. Columbianus: the Колумбийский острохвостый тетерев можно найти в изолированных карманах родного полынь и кустарниковые равнины Айдахо, Вайоминг, Колорадо, Юта, и Британская Колумбия.
- Т. П. Campestris: в степях обитает острохвостый тетерев Саскачеван, юго-восток Манитобы, юго-запад Онтарио и Верхний полуостров Мичиган в северную Миннесоту и северную Висконсин. Этот подвид сосуществует с равнинной расой вокруг северных Долина Красной реки и кустарникам предпочитает низколежащие стадии недавно переустроенных лесов.
- Т. П. джамеси: равнинный острохвостый тетерев поселился на севере Большие равнины в южной Альберте и Саскачеване, восточной Монтана, север и южная Дакота, Небраска и северо-восточный Вайоминг. Эта раса обитает в прериях со смешанной травой, предпочитая мозаику из местных лугов, пахотных земель и заросших кустарником / древесными прибрежными зарослями, ручьями и реками в качестве источника зимней пищи над снежным покровом, так как бутоны и ягоды.
- Т. П. Хуэйи: the Нью-Мексико острохвостый тетерев вымерший.
Описание

У взрослых особей относительно короткий хвост с двумя центральными (палубными) перьями с квадратными кончиками и несколько длиннее, чем их более светлые внешние рулевые перья, что дает птице свое отличительное название. Оперение пестрое, темное и светло-коричневое на белом фоне, они светлее на нижней части с белым брюшком, равномерно покрытым слабыми V-образными пятнами. Эти отметины отличают острохвостых куропаток от малых и больших прерийных кур, у которых на нижней части туловища имеется сильная решетка (Connelly et al. 1998). Взрослые самцы имеют желтый гребешок над глазами и фиолетовое пятно на шее. Это демонстрационное пятно является еще одной отличительной особенностью прерийных кур, поскольку самцы прерийных кур имеют желтые или оранжевые воздушные мешочки (Коннелли и др., 1998). Самка меньше самца, и ее можно отличить по правильным горизонтальным отметкам на палубных перьях, в отличие от неровных отметин на палубных перьях самцов, идущих параллельно стержню пера. У самок также, как правило, менее заметные гребни.
Измерения[3]:
- Длина: 15,0-19,0 дюймов (38,1-48,3 см)
- Масса: 21,0-31,0 унции (596-880 г)
- Размах крыльев: 24,4-25,6 дюймов (62-65 см)
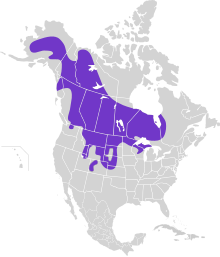
Распределение
Исторически острохвостый тетерев населял восемь канадских провинций и 21 штат США до европейских поселений (Johnsgard 2002). Они простирались от севера до Аляски, на юг до Калифорнии и Нью-Мексико и на восток до Квебека, Канада (Johnsgard 2002). После европейского заселения острохвостый тетерев был истреблен из Калифорния, Канзас, Иллинойс, Айова, Невада и Нью-Мексико (Johnsgard 1973; Connelly et al. 1998).
Поведение

Кормление
Летом эти птицы кормятся на земле, зимой - на деревьях. Они едят семена, почки, ягоды, разнотравье и листья, а также насекомые, особенно кузнечики, летом.

Разведение
Острохвостый тетерев - это вид птиц-тококов. Эти птицы появляются на открытых площадках, известных как токи с другими самцами от одного до двадцати самцов будет занимать один лек (в среднем 8-12). Лек определяется как место сбора, где животные демонстрируют свое поведение и ухаживают (Merriam-Webster Dictionary 2010). Весной самцы острохвостого тетерева посещают эти токи с марта по июль, причем пик посещаемости приходится на конец апреля - начало мая (Conelly et al. 1998). Эти даты меняются из года в год в зависимости от погоды. Johnsgard (2002) наблюдал в Северной Дакоте задержку протекания острохвостого тетерева на срок до двух недель. Самцы показывают на токе, быстро топая ногами, примерно 20 раз в секунду, и трясут хвостовыми перьями, вращаясь по кругу или танцуя вперед. Во время показа фиолетовые шейные мешочки надуваются и сдуваются. Самцы используют «воркование» также для привлечения самок и конкуренции за них (Sisson 1969; Bergerud and Gratson 1988; Conelly et al. 1998). Самки выбирают одного или двух наиболее доминирующих самцов в центре лека, совокупляются, а затем уходят в гнездо и выращивают детенышей в одиночку от самца. Иногда самец низкого ранга может действовать как женщина, приближаться к доминирующему самцу и драться с ним.
Выбор среды обитания
Острохвостый тетерев встречается в различных экосистемах прерий Северной Америки. Они населяют экосистемы от сосновых саванн на востоке верхнего Среднего Запада до низкотравных, среднетравных и кустарниковых степных прерий Великих равнин и Западных Скалистых гор (Aldrich, 1963; Johnsgard, 1973; Johnsgard, 2002). Выбор конкретных характеристик местообитаний и растительных сообществ различен для разных подвидов острохвостого тетерева. Выбор этих конкретных местообитаний зависит от качества среды обитания, доступной для тетерева (Johnsgard 1973; Kohn 1976; Swenson 1985; Roersma 2001; Goddard et al. 2009). Основные места обитания острохвостого тетерева, указанные в литературе, включают прерия в стиле саванны с преобладанием трав и смешанными участками кустарников, с минимальными участками деревьев (Hammerstrom 1963; Robel 1972; Moyles 1981; Johnsgard 2002; Goddard et al. 2009). Фактически, Хаммерстром (1963) утверждает, что чем выше древесная растительность, тем меньше ее должно быть в среде обитания. Среда обитания в стиле саванны в основном предпочтительна летом и в месяцы выращивания выводков до осени. Эта общая среда обитания используется в течение всех четырех сезонов для различных целей. Выбор и использование среды обитания зависит от сезона; по-разному выбираются и используются токи, гнездование, выращивание выводков и зимняя среда обитания.
Леккинг среда обитания
Лек, или танцплощадка, обычно состоит из короткой, относительно плоской местной растительности (Manske and Barker 1987; Hanowski et al. 2000). Другими типами местообитаний, используемых для токов, являются возделываемые земли, недавние пожары, скошенные участки, пастбища на вершинах холмов и влажные луга (Ammann 1957; Kobriger 1965; Johnsgard 1973; Johnsgard 2002). Manske и Barker (1987) сообщили о солнечной осоке (Осока инопс ), иглы и нитки травы (Hesperostipa comata ) и синяя грама (Bouteloua gracilis ) на водоёмах в Национальных пастбищах Шейенн в Северной Дакоте. Самцы также выбирают среду обитания на возвышенностях или в средней полосе на вершинах хребтов или холмов (Manske and Barker 1987). Лекы, окруженные высокой остаточной растительностью, наблюдались Kirsch et al. (1973). Они заметили, что на распределение лека повлияло количество высокой остаточной растительности, прилегающей к току. Участки Лека в конечном итоге стали заброшенными, если структура растительности стала слишком высокой. Вторжение древесной растительности и деревьев на арены течения также привело к тому, что самцы бросили токи (Moyles 1981; Swenson 1985). Moyles (1981) наблюдал обратную зависимость посещаемости лекаря мужчинами от увеличения числа трясущихся осин (Populus tremuloides ) в пределах 0,8 км от арен в парковой зоне Альберты. Бергер и Байдак (1992) также наблюдали аналогичную тенденцию в зарастании осин, когда 50% (7 из 14) токов были заброшены, когда покрытие осиной увеличивалось до более 56 процентов от общей площади в пределах 1 км от лека. Самцы выбирают для леков вершины холмов, хребтов или любое место с хорошим обзором. Таким образом, они могут видеть окружающих: самцов, приближающихся самок к танцплощадке и хищников (Sisson 1969; Manske and Barker1987; Johnsgard 2002).
Гнездовая среда
Гнездовой покров - один из важнейших типов среды обитания кур острохвостого тетерева. Среда гнездования сильно различается у разных подвидов острохвостого тетерева (Roersma 2001). Хамерстром-младший (1939) обнаружил, что большинство гнезд острохвостого тетерева (T.p.campestris) занимают густые заросли кустарников и леса по краям болот. Гизон и Коннелли (1993) сообщили, что колумбийский острохвостый тетерев (T.p.columbianus) выбирает густые заросли кустарников с более высокими и густыми кустами, расположенными в месте гнездования. Равнинный острохвостый тетерев (T.p. jamesii) выбирал места для гнезд с густой остаточной растительностью и кустарниковой составляющей (Kirby and Grosz 1995; Roersma 2001). Однако места гнездования обычно характеризуются густой высокой остаточной растительностью (прошлогодний рост) с наличием древесной растительности либо на месте гнездования, либо поблизости (Manske and Barker 1987; Prose et al. 2002). Годдард и др. (2009) утверждают, что использование местообитаний с преобладанием кустарников не было задокументировано многими другими исследователями. Годдард и др. (2009) обнаружили, что острохвостые куропатки в Альберте, Канада, выбирали больше для кустарниковых степных местообитаний при первых попытках гнездования из-за большей маскировки, обеспечиваемой кустарниками, чем остаточной травой в начале сезона размножения. Роерсма (2001) также обнаружил, что куропатки в южной части Альберты выбирают более высокую древесную растительность по сравнению со всеми другими оцениваемыми местообитаниями, а куропатки используют эту территорию в большей пропорции по сравнению с доступной древесной средой обитания. Эти данные противоречат Prose et al. (2002), который утверждает, что остаточная растительность имеет решающее значение для успеха гнездовий острохвостого тетерева из-за раннего сезонного характера гнездования тетерева.
Среда обитания выводка
Острохвостый тетерев - это скороспелый вид, это означает, что они вылупляются с открытыми глазами, самостоятельны и не требуют, чтобы мать их кормила. Вскоре после вылупления птенцы и мать покидают гнездо в поисках укрытия и корма. Места выращивания потомства острохвостого тетерева имеют много характеристик, включая кустарниковую растительность для укрытия, короткую растительность поблизости для кормления и большое количество разнотравья (Hamerstrom 1963; Kohn 1976; Manske and Barker 1987; Roersma 2001; Goddard et al. 2009) ). Это может объяснить, почему острохвостый тетерев гнездится в кустарниковых сообществах или поблизости от них. Компонент кустарника в среде обитания для насиживания обеспечивает хорошую защиту полога от прямых солнечных лучей и птичьих хищников (Roesmra 2001; Goddard 2009). Hamerstrom (1963) и Goddard et al. (2009) оба наблюдали наибольшее количество выводков острохвостого тетерева, обитающих на открытых, а не лесных ландшафтах. Оба предположили, что такое использование открытого ландшафта было связано с обилием насекомых для цыплят и зеленым травяным покровом для курицы, которым они могли питаться. Использование среды обитания выводками острохвостого тетерева зависит от времени суток, доступной среды обитания и погоды ( Амманн 1957; Кон 1976). Среда обитания выводка состоит из множества сложных типов среды обитания. Выводки могут использовать кустарники или места обитания дубовых лугов и саванн (Hamerstrom 1963). Выводки используют эти типы местообитаний для укрытия, оставаясь при этом близкими к основным местам кормления в виде более короткой растительности со смесью местной растительности.
Зимняя среда обитания
Использование острохвостого тетерева в зимнее время, похоже, смещается в сторону более плотного укрытия для теплоизоляции. Хаммерстром и Хаммерстром (1951) заметили, что в Мичигане и Висконсине зимой в Мичигане и Висконсине куропатки используют среду обитания с более толстыми краями чаще, чем открытый грунт. Хаммерстром и Хаммерстром (1951) также отметили, что птицы, обнаруженные в открытой среде обитания, находились не более чем в нескольких сотнях метров от более толстого покрова. Эти птицы обычно использовали зерновые поля. Свенсон (1985) наблюдал ту же тенденцию в Монтане. Хамерстром и Хаммерстром (1951) заявили, что использование лесных местообитаний острохвостыми тетеревами различается в зависимости от местоположения, отметив, что острохвостые тетерева в более полузасушливых и засушливых районах реже используют кусты зимой. Однако Хаммерстром и Хаммерстром (1951) сообщили, что острохвостые тетерева в Вашингтоне и Калифорнии чаще наблюдались в местах обитания краевого типа в зимние месяцы. Манске и Баркер (1987) заметили аналогичную тенденцию в использовании зимних местообитаний в Северной Дакоте, отметив, что острохвостые глухари в небольших стаях объединяются, образуя большие стаи в суровую погоду. Эти стаи перемещаются из открытых прерий в лесополосы и прилегающие пахотные земли с кукурузой и подсолнухами. Использование среды обитания зимой сильно зависит от высоты снежного покрова (Swenson 1985). По мере увеличения высоты снежного покрова выбор среды обитания смещается от пахотных земель и прерий к лесополосам и древесной растительности. Одно изменение среды обитания, замеченное Хамерстромом и Хамерстромом (1951), заключалось в том, что тетерева выбирали большие снежные насыпи, чтобы зарыться в них и согреться в холодные ночи. Использование нор было также отмечено Gratson (1988).
Фрагментация среды обитания
Фрагментация среды обитания был одним из факторов, способствовавших сокращению численности всех подвидов острохвостого тетерева на всем его ареале по всей Северной Америке (Silvy and Hagen 2004). Тип фрагментации среды обитания варьируется от экологической сукцессии, поскольку участки кустарников / пастбищ переходят в лесные массивы. Тушение пожаров, посадка деревьев, ограничение практики лесозаготовок и рост инвазивных древесных пород также привели к фрагментации среды обитания. Наибольший вклад в фрагментацию среды обитания вносит сельское хозяйство.
Закон о усадьбе 1862 года открыл для первых поселенцев просторы девственных прерий на западе. К 1905 году около 41 миллиона гектаров на западе были заселены (Olsen 1997). Большая часть этой земли находилась в полузасушливых пастбищах с субмаргинальными осадки для поддержки растениеводства (Olsen 1997). Вспашка этой земли представляла собой постоянное изменение ее характера. Еще один фактор, способствующий фрагментации среды обитания тетерева, - это бесконтрольный и чрезмерный выпас скота (Кирш и др., 1973; Гизен и Коннелли, 1993; Кирби и Гросс, 1995; Рис и др., 2001; Сидл, 2005). При правильном содержании крупный рогатый скот может быть важным инструментом управления структурой среды обитания острохвостого тетерева (Evens 1968; Kirby and Grosz 1995; Sidle 2005). Среда обитания острохвостого тетерева серьезно пострадала от первых поселенцев до того, как пастбища поняли, какое воздействие на окружающую среду оказывает чрезмерный выпас скота.
Вторичным эффектом раннего земледелия в годы Пыльной чаши и Великой депрессии в конце 1920-х и начале 1930-х годов было то, что поселенцы покидали непродуктивные земли (Olsen 1997). Правительство Соединенных Штатов скупило большую часть этой земли через Программу землепользования, управление которой в конечном итоге перешло под контроль Лесной службы США и Бюро землепользования (Wooten 1965; Olsen 1997). В течение засушливых лет 1930-х годов эти агентства восстановили некоторые из этих территорий с помощью чужеродной высококонкурентной растительности, такой как бром (Bromus inermis) и пырей (Agropyron cristatum) (USDA-US Forest service 2001). Эти растения служили своей цели, восстанавливая растительность и защищая почву. Но эти захватчики стали серьезными конкурентами и напрямую повлияли на местную растительность. В некоторых случаях пырей и кострец вытеснили местную растительность, создав среду обитания монокультур. Места обитания монокультур не подходят для острохвостого тетерева, поскольку они предпочитают участки с высокой неоднородностью. Хамерстром (1939) сказал: «Более важным, чем отдельные покровные растения, является тот факт, что большинство гнезд всех видов находились в смеси покровов, а не в чистых насаждениях».
Оценка среды обитания
Исследования по оценке местообитаний острохвостого тетерева, проводившиеся до 1950 г., проводились визуально. Хамерстром (1939) сообщил, что скудная растительность редко выбиралась для гнездования из-за отсутствия адекватного покрытия. Обобщения среды обитания были сформированы на основе количества особей, найденных в данном месте. Эти предположения были если больше птиц было в одном месте и меньше в другом, тогда во-первых, должна быть лучшая среда обитания. Хамерстром (1963) наблюдал 119 из 207 (57%) выводков тетерева, часто посещающих места обитания в стиле саванны. Он пришел к выводу, что среда обитания в стиле саванны - это среда, необходимая для лучшего управления. По мере развития исследований среды обитания видов тетерева, совершенствовались и методы оценки. Покрытия и столбы Robel были разработаны для измерения визуальных препятствий (VO) и создания индексов среды обитания. Покровные доски были разработаны Уайтом (1938) еще в 1938 году для изучения среды обитания белохвостого оленя. Покровная доска Уайта (1938) была 6 футов высотой, каждая была помечена и пронумерована. Видимые отметки подсчитывались для измерения препятствий, создаваемых растениями. Кобригер (1965) разработал доску размером 4 × 4 фута, размеченную с интервалом в 3 дюйма чередующимися белыми и черными квадратами. Он разместил камеру в центре нерестилища на высоте 3 фута. Затем он поместил защитную доску на расстоянии 30 футов и сфотографировал ее. После компиляции всех фотографий они были проанализированы с помощью ручного объектива, чтобы оценить количество видимых квадратов. Это число дало ему индекс растительности по классам покрова. Этот метод был модифицирован Limb et al. (2007). Вместо того, чтобы делать фотографии на расстоянии 30 футов, как Кобригер (1965), Лимб и др. (2007) сфотографировали растительность, сброшенную на защитную доску размером 1 × 1 метр на высоте 1 метр на расстоянии 4 метра. Эти цифровые фотографии были загружены в Adobe Acrobat и оцифрованы на заднем фоне размером 1 × 1 метр (Лимб и др., 2007). Robel et al. (1970) разработали шест для определения роста на основе коррелированного веса растительности. Полюс был должным образом назван Шест Робель. Robel et al. (1970) обнаружили, что измерения VO, сделанные на высоте 1 м и на расстоянии 4 м от столба, дали надежный показатель количества растительности в данном месте. Hamerstrom et al. (1957) были процитированы слова: «Высота и густота травы были явно более важны для прерийных кур, чем видовой состав», как сообщили Робель и др. (1970). Считалось, что это справедливо и для острохвостого тетерева. Эти ключевые аспекты теперь можно оценить с помощью шеста Robel, крышки Nudds и Limb et al. метод цифровой фотографии эффективно и рационально.
Управление
Очевидно, что эффекты фрагментации местообитаний по всем типам местообитаний, выбранных острохвостым тетеревом, влияют на этот вид. За прошедшие годы управление средой обитания острохвостого тетерева изменилось с наблюдательного (обеспечение сохранения нынешней среды обитания) на более практический подход. Управление местами обитания течи и зимними местообитаниями не так четко определено в литературе, как оценка и управление местами обитания для гнездования и разведения выводков. Разработка столбов Robel и укрытий стала ключевым инструментом в оценке среды обитания, предоставляя управляющим земельными ресурсами средство для инвентаризации и изучения предпочтений среды обитания на основе структуры и плотности растительности. В последние годы полюс Робеля стал более популярным из двух методов оценки среды обитания. Лесная служба США (USFS) использует показания визуальных препятствий (VOR) для определения плотности поголовья крупного рогатого скота на основе существующей остаточной растительности текущего года. Этот метод в настоящее время применяется на пастбищах USFS Little Missouri Grasslands, Sheyenne National Grasslands, Cedar River National Grassland и Grand River National Grassland, которые находятся на национальных пастбищах Dakota Prairie National Grasslands в Северной и Южной Дакоте (Лесная служба Северного региона (USDA). Полюс Робеля - это неразрушающий метод инвентаризации растительной биомассы (Робель, 1970; Бенкоби и др., 2000). Этот метод использовался для создания индекса пригодности среды обитания на основе визуального препятствия растительности (VO) в диапазоне от 0 до 30,5 см с рейтингом индекса пригодности от 0 до 1,0 (Prose 1987). Исследования гнездовой среды Prose et al. (2002) в Песчаных холмах Небраски обнаружили, что для гнездования острохвостого тетерева выбирались места для гнезд с показаниями визуального препятствия (VOR) более 4 см. Точно так же Клоусон и Роттелла (1998) наблюдали, что 58% гнезд (432 из 741) в Юго-Западной Монтане были расположены на участках со средним VOR 24 см. Остальные гнезда в этом исследовании были расположены на участках с VOR 11–18 см. Рис и др. (2001) отметили, что участки с VO менее 5 см рядом с возможными местами гнездования указывают на снижение качества гнездовой среды, так как средний VO снижается. Использование шеста Робеля для оценки среды обитания острохвостого тетерева дало менеджерам целевую высоту структуры растительности, которую необходимо иметь в конце пастбищного сезона. Это позволяет менеджерам устанавливать соответствующую норму поголовья для достижения желаемой высоты растительности. Как показывает практика, среднее значение VOR для подходящей среды гнездования тетерева составляет 8,89 см (3,5 дюйма). Средой обитания леккинга можно управлять путем сжигания, скашивания, сплошных рубок и выпаса скота на всем ареале подвида острохвостого тетерева. Амманн (1957) обнаружил, что токи, содержащие древесную растительность, не превышали 30% от общей площади течения. Точно так же Moyles (1989) обнаружил отрицательную корреляцию с увеличением количества осин (Populus tremuloides) на участках течения и количеством присутствующих самцов. Деревья могут служить насестами для птичьих хищников, но необходимо провести дополнительную работу по устранению последствий посягательство на деревья (Манзер и Хэннон, 2005).
Провинциальная птица
Острохвостый тетерев - это провинциальная птица из Саскачеван.
Статус и сохранение
Эти птицы сокращаются в численности и ареале из-за потери среды обитания, но в целом они не считаются угрожаемыми видами.

Примечания
- ^ BirdLife International (2012). "Tympanuchus phasianellus". Красный список видов, находящихся под угрозой исчезновения МСОП. 2012. Получено 26 ноября 2013.CS1 maint: ref = harv (связь)
- ^ Хоффман и другие. (2007), стр. 15.
- ^ "Идентификация острохвостых тетеревов, Все о птицах, Корнельская лаборатория орнитологии". www.allaboutbirds.org. Получено 2020-09-26.
Рекомендации
- Олдрич, Дж. 1963. Географическая ориентация американских Tetraonidae. Журнал управления дикой природой 27: 529-545.
- Амманн, Г.А. 1957. Мичиганский глухарь. Технический бюллетень Департамента охраны природы штата Мичиган.
- Benkobi, L., D.W. Уреск, Г. Шенбек и Р. М. Кинг. 2000. Протокол мониторинга стоячих культур на пастбищах с использованием визуальных препятствий. Журнал Range Management, Vol. 53, № 6, стр. 627–633
- Бергер, Р.П. и Р.К. Байдак. 1992 Влияние сукцессии осины на острохвостого тетерева, Tympanuchus phasianellus. В межозерном районе Манитобы. Канадский естествоиспытатель 106: 185-191
- Бергеруд, А. Т. 1988. Системы спаривания у тетерева. Страницы 439-470 в Адаптивные стратегии и популяционная экология тетерева. (Bergerud, A. T. и M. W. Gratson, Eds.) Univ. Миннесота Пресс, Миннеаполис.
- Bergerud, A. T. и M. W. Gratson 1988. Популяционная экология североамериканского тетерева. Страницы 578-685 в Адаптационные стратегии и популяционная экология тетерева. (Bergerud, A. T. и M. W. Gratson, Eds.) Univ. Миннесота Пресс, Миннеаполис
- Кларк, Джулия А. (2004): Морфология, филогенетическая таксономия и систематика Ихтиорнис и Апаторнис (Avialae: Ornithurae). Бюллетень Американского музея естественной истории 286: 1-179 Полный текст PDF
- Клоусон, М.Р., и Дж. Дж. Ротелла. 1998. Успех искусственных гнезд на полях CRP, естественной растительности и границах полей в Юго-Западной Монтане. Журнал полевой орнитологии, Vol. 69, № 2, с. 180–191
- Коннелли, Дж. У., М. В. Гратсон и К. П. Риз. 1998. Острохвостый тетерев (Tympanuchus phasianellus), Птицы Северной Америки в Интернете (А. Пул, ред.). Итака: Корнельская лаборатория орнитологии; Получено из Интернета птиц Северной Америки: http://bna.birds.cornell.edu/bna/species/354
- Коновер, М. Р. и Дж. Борго. 2009. Выбирают ли острохвостые тетерева места для бездельничанья, чтобы избежать зрительных или обонятельных хищников? Журнал управления дикой природой, Vol. 73, № 2, стр. 242–247
- Гизен, К. М. и Дж. У. Коннелли. 1993. Руководство по управлению местообитаниями колумбийского острохвостого тетерева. Бюллетень Общества дикой природы, Том. 21 Выпуск 3, стр. 325-333
- Годдард, А. Д .; Р. Д. Доусон; М.П. Джиллингем. 2009. Выбор местообитания путем гнездования и выведения потомства острозубых тетеревов. Канадский зоологический журнал, апрель 2009 г., Vol. 87 Выпуск 4, с326-336, 10п, 6 графиков, 2 графика; Дои:10.1139 / Z09-016; (AN 37580857)
- Гратсон, М. В. 1988. Пространственные модели, движения и выбор укрытия острохвостым тетеревом. Страницы 158-192 в Адаптивные стратегии и популяционная экология тетерева. (Bergerud, A. T. и M. W. Gratson, Eds.) Univ. Миннесота Пресс, Миннеаполис
- Hamerstrom Jr, F. N. и F. Hamerstrom. 1951. Подвижность острохвостого тетерева в связи с его экологией и распространением. Американский натуралист из Мидленда. Vol. 46, No. 1 (июль 1951 г.), стр. 174–226 Издатель: Университет Нотр-Дам
- Хамерстром-младший, Ф. Н. 1963. Места обитания острохвоста в северных сосновых степях штата Висконсин. Журнал управления дикой природой, Vol. 27, № 4. 793-802
- Hanowski, JAM, D.P. Кристиан и Г.Дж. Ниеми. 2000. Требования к ландшафту прерийного острохвостого тетерева Tympanuchus phasianellus campestris в Миннесоте, США. Биология дикой природы Vol. 6, вып. 4. С. 257–263.
- Хендерсон, Ф. Р., Ф. У. Брукс, Р. Э. Вуд и Р. Б. Дальгрен. 1967. Определение пола глухаря по рисунку перьев кроны. Журнал управления дикой природой. 31: 764-769.
- Hoffman, R.W .; И Томас, A.E. (17 августа 2007 г.): Колумбийский острохвостый тетерев (Tympanuchus phasianellus columbianus): техническая оценка сохранения. [В сети]. Лесная служба Министерства сельского хозяйства США, регион Скалистых гор. 132 стр. Полный текст PDF Проверено 28 декабря 2008.
- Johnsgard, P.A. 1973. Острохвостый тетерев. С. 300–319 в Тетерева и перепела Северной Америки. Университет Линкольна Небраски.
- Джонсгард, П.А., 2002. Танцовщицы зари на серой траве. Острохвостый тетерев и северные прерии и кустарники. Страницы 81–103 в Рекавере. (Johnsgard, P.A.) Smithsonian Institution Press. Вашингтон и Лондон
- Джонс, Р. Э. 1968. Доска для измерения укрытия, используемая Прерий Гроуз. Журнал управления дикой природой, Vol. 32, № 1, с. 28–31
- Кирш, Л.М., А.Т. Клетт и Х. В. Миллер. 1973 г.Землепользование и отношения населения луговых куропаток в Северной Дакоте. Журнал управления дикой природой, Vol. 37, No. 4 (октябрь 1973 г.), стр. 449–453
- Кобригер, Г. Д. 1965. Статус, перемещения, среды обитания и пища глухаря на заповеднике Sandhills. Журнал управления дикой природой, Vol. 29, No. 4 (октябрь 1965 г.), стр. 788–800
- Кон, С. С. 1976. Гнездование и насиженная среда острохвостого тетерева на юго-западе Северной Дакоты, Департамент охоты и рыболовства Северной Дакоты,
- Лек. (2010). В онлайн-словаре Merriam-Webster. Получено 16 апреля 2010 г. из http://www.merriam-webster.com/dictionary/lek
- Лимб, Р.Ф., К.Р. Хикман, Д.М. Энгл, Дж.Э. Норланд и С.Д. Фулендорф. 2007. Цифровая фотография: уменьшает вариации в измерениях визуальных препятствий для южных высокотравных прерий. Управление экологии земель пастбища. 60: pp. 548–552.
- Manske, L.L .; В. Т. Баркер. 1987. Использование местообитаний луговыми тетеревами на национальных пастбищах Шейенн. С. 8–20. В: A. J. Bjugstad, tech. Координатор Цыплята прерий на национальных пастбищах Шейенн. Gen. Tech. Реп. RM-159. Форт-Коллинз, Колорадо: Министерство сельского хозяйства США, Экспериментальная станция лесов и хребтов Скалистых гор. 73 с
- Moyles, DLJ. 1981. Сезонное и ежедневное использование сообществ растений острохвостым тетеревом (Pedioecetes phasianellus) в парковых зонах Альберты. Канадский естествоиспытатель. Оттава ON Vol. 95, нет. 3. С. 287–291.
- Nudds, T. D. 1977. Количественная оценка растительной структуры дикой природы. Бюллетень Общества дикой природы. Vol. 5, № 3, с. 113–117
- Олсен, E. 1997. Учебник по управлению национальными пастбищами. Управление природных ресурсов Генерального совета USDA. стр. 1–40
- Проза, Б. (1987): Модели индекса пригодности местообитаний: равнинный острохвостый тетерев. U.S. Fish Wildl. Серв. Биол. Rep. 82: 10.142. 31 стр. Полный текст PDF
- Проза, Б.Л., Б.С. Кейд и Д. Хайн. 2002. Выбор места гнездования острохвостого тетерева в песчаных холмах Небраски. Натуралист прерий. 34 (3/4): 85-105.
- Рис П.Е., Дж.Д. Волески и У. Schacht. 2001. Укрытие для дикой природы после летнего выпаса на пастбищах у песчаных холмов. Журнал Range Management. Vol. 54, № 2, с. 126–131.
- Робель, Р.Дж., Дж. Бриггс, А.Д. Дейтон и Л.С. Хульберта. 1970. Связь между измерениями визуальных препятствий и весом пастбищной растительности. Журнал Range Management. 23: 295-297.
- Робель, Р. Дж., Р. Ф. Хендерсон и У. Джексон. 1972. Некоторые статистические данные о популяции острохвостого тетерева из Южной Дакоты. Журнал управления дикой природой, Vol. 36, № 1 (январь 1972 г.), стр. 87–98.
- Роерсма, С.Дж. 2001. Экология гнездования и выращивания выводков равнинного острохвостого тетерева (Tympanuchus phasianellus jamesi) в смешанно-злаково-овсяничном экорегионе Южной Альберты. M.Sc. докторская диссертация, Университет Манитобы, Виннипег.
- Silvy, N.J. и C.A. Hagen. 2004 г. Введение: Управление видами лугового тетерева, находящимися под угрозой, и их средой обитания Бюллетень Общества дикой природы, март 2004 г .: Vol. 32, выпуск 1, стр. 2-5 Дои:10.2193 / 0091-7648 (2004) 32 [2: IMOIPG] 2.0.CO; 2
- Сиссон, Л. 1969. Изменения в землепользовании и племенное поведение острохвостых куропаток Белые книги Комиссии по охоте и паркам штата Небраска, презентации на конференциях и рукописи.
- Свенсон, Дж. Э. 1985. Сезонное использование среды обитания острохвостым тетеревом Tympanuchus phasianellus в прериях со смешанной травой в Монтане. Может. Поле-Нат. 99: 40-46. ЙОКОМ, К. Ф. 1952.
- План управления земельными ресурсами USFS для северного региона Dakota Prairie Grasslands, 2001 г.
- Уайт, Х. 1938. Полевые и лабораторные методы в природопользовании. Univ. из Michigan Press, Анн-Арбор. 108pp.
- Вутен, Х. Х. «Программа землепользования с 1934 по 1964 год: происхождение, развитие и нынешнее состояние», в Приложении C к Руководству по управлению национальными пастбищами (1965).
внешняя ссылка
- Драка острохвостого тетерева в суперзамедленном видео из Корнельской лаборатории орнитологии
- Острохвостый Грус (так в оригинале) - Джон Джеймс Одюбон Крупные планы высокого разрешения от "Птицы Америки"
- Учет видов острохвостых тетеревов - Корнельская лаборатория орнитологии
- Острохвостый тетерев Tympanuchus phasianellus - Информационный центр идентификации птиц USGS Patuxent
Дичь и стрельба в Северной Америке | ||
|---|---|---|
| Игровые птицы |  | |
| Водоплавающих птиц | ||
| Большая игра | ||
| Другой карьер | ||
| Смотрите также | ||