Бассейн Кокинетас - Cocinetas Basin
| Бассейн Кокинетас | |
|---|---|
| Cuenca Cocinetas | |
 Расположение бассейна в Колумбии | |
| Координаты | 11 ° 58′00 ″ с.ш. 71 ° 22′43 ″ з.д. / 11,96667 ° с.ш. 71,37861 ° з.д.Координаты: 11 ° 58′00 ″ с.ш. 71 ° 22′43 ″ з.д. / 11,96667 ° с.ш. 71,37861 ° з.д. |
| Этимология | Кокинетас Бэй |
| Область, край | Карибский бассейн Ксерический скраб Guajira-Barranquilla экорегион |
| Страна | Колумбия |
| Состояния) | Ла Гуахира |
| Города | Урибия |
| Характеристики | |
| On / Offshore | На берегу |
| Границы | Серрания-де-Харара, Серрания-де-Макуира, Залив Венесуэлы, Серрания Кочинас |
| Часть | Прикарибские бассейны |
| Площадь | ~ 1,000 км2 (390 кв. Миль) |
| Гидрология | |
| Моря) | Тортугас или залив Тукакас, залив Кокинетас Залив Венесуэлы Карибское море |
| Геология | |
| Тип бассейна | Раздвижная раковина |
| Пластина | Северные Анды |
| Орогенез | Андский |
| Возраст | Средний эоцен -Голоцен |
| Стратиграфия | Стратиграфия |
| Неисправности | Cuisa & Macuira (ограничивая) |
| Поле (я) | никто |
В Бассейн Кокинетас (испанский: Cuenca Cocinetas) - небольшой осадочный бассейн примерно 1000 квадратных километров (390 квадратных миль) на северо-востоке Колумбия. В береговой раздвижной бассейн расположен в отделение из Ла Гуахира на границе с Зулия, Венесуэла. Бассейн ограничен тремя группами холмов; Серрания-де-Харара, Серрания-Макуира и Серрания-Кочинас, с восточной границей, образованной Залив Венесуэлы, часть Карибское море.
Бассейн образовался во время Палеоген в результате движения на восток Карибская плита вдоль северного края Южноамериканская плита, что приводит к осаждению Средний эоцен к Средний плейстоцен осадочная толща конгломераты, песчаники, алевролиты, известняки и аргиллиты.
В бассейне Кокинетас несколько стратиграфические единицы, содержащие ископаемые были зарегистрированы, обеспечивая обилие сообществ ископаемой фауны морских и континентальных позвоночных и беспозвоночных. Формации Уитпа, Джимол, Кастиллетес и Вар содержат многочисленные окаменелости Неоген и Четвертичный возрастов, как до, так и после Великий американский биотический обмен (ГАБИ), представляя новые идеи в понимании изменчивости фауны Южной Америки, связанной с поднятием Панамского блока и соединением Северной Америки и Южной Америки.
Описание

Бассейн Кокинетас - небольшой береговой осадочный бассейн примерно 1000 квадратных километров (390 квадратных миль) в самой северо-восточной части Колумбия.[1] Самый северный береговой осадочный бассейн Южной Америки ограничен по часовой стрелке рекой. Серрания-де-Харара, то Macuira Fault отделяя бассейн от Серрания-де-Макуира, заливы Тукакас или Тортугас и Кокинетас как часть Залив Венесуэлы и Серрания-де-Кочинас (в более ранней литературе упоминается как Cosinas),[2] разделены Cuisa Fault.[3]
Ограничивающий разлом на юге, правый разлом Cuisa с боковым смещением 15 километров (9,3 мили),[4] имеет протяженность примерно на 80 километров (50 миль) по оси примерно восток-запад, а разлом продолжается на восток под Венесуэльским заливом. Разлом частично перекрыт олигоценовыми отложениями.[5] Северная граница раздвижной бассейн образован разломом Macuira, простирающимся примерно с северо-запада на юго-восток, с протяженностью около 40 километров (25 миль). Левосторонний косой разлом имеет смещение около 3 км (1,9 мили) и поднял северо-восточную часть Серрания-де-Макуира.[6]
Бассейн полностью расположен в муниципалитет Урибия из отделение из Ла Гуахира. Самая южная оконечность бассейна граничит с Венесуэльский состояние Зулия.[3] Деревни Уитпа, Кастильетес и Пуэрто-Лопес расположены внутри бассейна.[3][7][8]
Современный климат в бассейне очень сухой и жаркий (Köppen: БШ), находясь в пустыне Ла Гуахира. Температура в тропиках на севере Южной Америки не сильно меняется в течение года. Июль - самый жаркий месяц со средней температурой 30,6 ° C (87,1 ° F), а январь имеет среднее значение 27,9 ° C (82,2 ° F). Дождь редкий в течение года, за исключением октября, самого влажного месяца, когда выпадает 109 миллиметров (4,3 дюйма) осадков.[9] Альта-Гуахира - самый засушливый регион Колумбии, и здесь часто бывают продолжительные периоды засухи.[10]
Стратиграфия

Стратиграфия бассейна Кокинетас была впервые определена Ренцем в 1960 году.[3] В 2015 году Морено и др. детально пересмотрел стратиграфию бассейна, переопределив возраст на основе обширных полевых исследований, биостратиграфия и 87Sr /86Sr изотопные отношения. В подвал бассейна, вырезание на окружающих холмах образован метаморфический Джарара Сланец и палеозой Macuira Gneiss.[8][11][12][13][14]
Седиментация в бассейне началась в среднем и позднем эоцене. Формация Макарао и продолжился снова в позднем олигоцене, когда Сиаманская формация, толща конгломератов и мощных карбонатов. Над формацией Сиамана залегает ранний миоцен. Формация Уитпа, мощная толща известковых аргиллитов с прослоями каменные песчаники. Свита Уитпа покрыта миоценом. Jimol и Кастильетские образования, состоящий в основном из каменных песчаников и аргиллитов, которые на некоторых участках содержат большое количество окаменелостей. Последняя фаза сохранившейся седиментации представлена плиоценом. Формирование посуды, определенные в 2015 году Морено и др.[15]
Третичный (Палеоген к Поздний плейстоцен ) Южной Америки подразделяется на собственную стратиграфию; то Возраст наземных млекопитающих в Южной Америке, (САЛМА). Это подразделение используется по всему континенту для обозначения определенных геологических интервалов на основе встречаемости конкретных наземных млекопитающих.[16]
Среды осадконакопления и корреляции

Фауна беспозвоночных свиты Джимол похожа на фауну последних Ранний миоцен Формация кантаура Венесуэлы и Формация Кулебра из Панама.[22] Формация Джимол была отложена в мелководной морской среде. внутренняя полка глубина (глубина воды менее 50 метров (160 футов)). Этот блок соотносится с верхним Агуа Клара и ниже Формации Серро Пелао венесуэльского Бассейн Фалькон.[23]
Фауна беспозвоночных формации Castilletes очень похожа не только на фауну нижележащей формации Jimol, но и на формацию Cantaure в Венесуэле.[24] Формация Castilletes откладывалась в мелководной морской среде (эстуарий, лагуна и мелководье сублиторали) до флювиодельтовой среды с сильным речным влиянием. Формация Castilletes коррелирует с верхними слоями Серро Пеладо и Формации Кералес венесуэльского Бассейн Фалькон. Эта единица также соотносится с Формация кантаура полуострова Парагуана в Венесуэле.[25]
Фауна морских беспозвоночных формации Вар показывает большее сходство с современными сообществами у берегов полуострова Гуахира, чем с таковыми из нижележащих подразделений.[26] Основание формации Вэр было отложено в флювиодельтовой среде, в то время как сообщество морских беспозвоночных в верхней части единицы содержит таксоны, типичные для открытой береговой линии открытого океана и прибрежных территорий, но с близостью к местообитаниям коралловых рифов. Формирование керамики коррелирует с Формация Сан Грегорио в венесуэльском Бассейн Фалькон.[27]
Тектоническая история
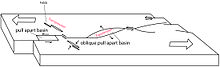
| Палеогеография Колумбии | |
 | 65 млн лет |
 | 50 млн лет |
 | 35 млн лет |
 | 20 млн лет |
 | Подарок |
Тектоническая история бассейна Кокинетас раздвижной бассейн сформированный на вершине Мезозойский подвал лежать в основе океаническая кора, возвращается к Палеоген.[28] Дальше на юг, в бассейне Сезар-Ранчерия, палеоцен, отсутствующий в бассейне Кокинетас, представлен первым в мире неотропным лесом с мегафауна как самая большая зарегистрированная змея, Titanoboa cerrejonensis, с предполагаемой длиной 14 метров (46 футов) и весом 1135 килограммов (2502 фунта), сегодня найденный в Формация Серрехон, датируемые 60-58 млн лет назад. Это было оценено на основе ископаемой флоры, пыльца и крупных рептилий, средняя годовая температура которых составляла от 28,5 до 33 ° C (83,3 и 91,4 ° F), а годовое количество осадков колебалось от 2 260 до 4640 миллиметров (89 - 183 дюйма) в год.[29]
К северу от Ока разлом, самая старая осадочная толща бассейна Кокинетас, формация Макарао, является старейшей осадочной единицей как более высокого района Ла-Гуахира (бассейн Кокинетас), так и нижняя часть Ла-Гуахира. Это образование отложилось в фазе, которая неофициально называется «рифтинг», из-за формирования половина грабенов в позднем эоцене, хотя реального рифтинга в этом районе не происходило.[30] Вскоре после осаждения, во время раннего олигоцена, формация Макарао была сильно деформирована, образовав серию холмов, которые сегодня все еще окружают бассейн Кокинетас.[31] Этот транстенсиональный деформация была вызвана движением на восток Карибская плита вдоль Южноамериканской плиты, когда Карибский бассейн вулканическая дуга был расположен к северу от бассейна Кокинетас.[32] Основное движение разломов, формирующих бассейн pull-apart Кокинетас, было вызвано правым разломом Cuisa, активным с палеоцена до эоцена.[28]
Транстенсионная фаза раннего олигоцена сменилась отложением несогласно перекрывающая сиаманская формация, характеризующаяся базальной последовательностью конгломератов, указывающая на тектоническое поднятие в происхождение области, за которыми следуют рифал известняки богат в кораллы и водоросли.[33] Новая фаза транстенсии произошла в раннем и среднем миоцене, когда были отложены формации Уитпа, Джимол и Кастильетес.[30] Начальная фаза подъема Панамский перешеек, датируется этим периодом около 12 млн лет назад.[34]
В Андская орогенез, представленный тектоническим поднятием Восточного хребта Колумбии и его северным продолжением, Серрания-дель-Периха, вызвало наклон и поднятие бассейна Кокинетас в позднем миоцене и раннем плиоцене, когда неогеновый перерыв отделял формацию Кастиллетес от формации Вар.[30] Конец неогена соответствовал постановке морской платформы, прерванной присутствием серии максимумов; современные горные массивы региона. Между этими возвышениями рифовые известняки откладывались в относительно спокойной морской среде.[35] Во время андской орогенной фазы палеотемпературы в бассейне значительно упали; в районе Баха Гуахира от 115 ° C (239 ° F) в раннем миоцене до 70 ° C (158 ° F) в позднем миоцене.[36] В период с позднего миоцена до плиоцена основные разломы к юго-западу от бассейна Кокинетас, Ока и Букараманга-Санта-Марта Разломы были тектонически активными.[37]
Палеонтология



Бассейн Кокинетас представил несколько новых видов и роды позвоночных и беспозвоночных, многие из которых до сих пор подробно изучены. В формациях Уитпа, Джимол, Кастиллетес и Вар сохранились как разнообразные континентальные позвоночные, так и богатая летопись окаменелостей морских беспозвоночных, которые документируют палеоэкологические изменения в течение неогена. Эта последовательность показывает широкую картину изменения среды осадконакопления, связанную с тектонической историей бассейна Кокинетас.[38] Палеобатиметрический и палеоэкологический анализ показывает, что нижняя формация Уитпа накапливалась на глубинах от 100 до 200 метров (от 330 до 660 футов), углубляясь по отношению к нижележащей формации Сиамана и приводя к обрушению коралловых рифов. Это контрастирует с наблюдаемым увеличением поступления наносов в экосистемы коралловых рифов Сан-Луис в северо-западной части бассейна Фалькон.[39]
Фауна черепах Castilletes разделяет два таксона со средним миоценом. Ла Вента Фауна (Chelus colombiana и Chelonoidis sp.) и один таксон с поздним миоценом Формация Урумако и районы Западной Амазонии. По крайней мере, две гипотезы могут быть предложены для объяснения географического распространения черепах в тропической Южной Америке в период от раннего до среднего миоцена. Первый основан на существовании Система водно-болотных угодий Pebas во время среднего миоцена, который мог служить маршрутом для распространения таксонов, чему способствовала связь между основными и второстепенными дренажами, даже с потенциальными перерывами в этой системе из-за возрастающего подъема Восточные хребты Колумбии. С другой стороны, географическое распространение тропических южноамериканских черепах в миоцене можно объяснить как пережиток обширного распространения, достигнутого в эоцене или олигоцене и впоследствии модифицированного поднятием Анд и изменениями в структуре основных речных систем.[40]
Возникновение Пуруссавр и Mourasuchus в раннем среднемиоцене пласты Патаяу и Кайтамана формации Кастильетес представляют собой ранние записи для линий, ранее известных из более молодых Лавентан и Huayquerian фауны. Эти записи расширяют временной диапазон высокоразнообразных ассоциаций гавиалоидов и кайманинов до раннего среднего миоцена. Летопись формации Кастильетес также расширяет географический диапазон миоценовых эндемичных фаун крокодилов до широт, эквивалентных широтам Центральной Америки.[41] Связи между гидрографическими бассейнами и развитием систем мега-водно-болотных угодий в экваториальной части Южной Америки позволили долгое время существовать нескольким родословным на обширном географическом ареале. Исчезновение гавиалоидов и специализированных кайманинов в экваториальной части Южной Америки, вероятно, было вызвано изоляцией и аридификацией периферийных бассейнов, а также исчезновением мега-водно-болотных угодий.[42]
Ленивцы из позднеплиоценовой керамической формации представлены 12 особями, относящимися к 5 таксонам из 4 семейств. наземные ленивцы. Размеры их тела превышают два порядка, и, скорее всего, они использовали различные стратегии кормления, что свидетельствует о очень разнообразном сообществе ленивцев из Неотропов. Хотя географически они близки к Панамскому перешейку и временно предшествуют первой фазе Великого американского биотического обмена всего на 200 000-400 000 лет, ленивцы, с которыми было обнаружено однозначное родство, не имеют тесного родства с ранними иммигрантами, обитавшими в Северной Америке до н. первый главный импульс Великого американского биотического обмена.[43] Зарегистрированный гигант Капибара Hydrochoeropsis wayuu это самая северная южноамериканская пластинка гидрохоринов плиоцена и ближайшая к Панамскому мосту. Присутствие этого гидрохорина вместе с флювиодельтовой средой формации Вар предполагает, что во время позднего плиоцена среда, которая доминировала на полуострове Гуахира, была более влажной и с постоянными водоемами, в отличие от современных пустынных местообитаний.[44]
Содержание ископаемых
Смотрите также
Рекомендации
- ^ Калькулятор площади Google Maps
- ^ Эрнандес Пардо и др., 2009, с.65.
- ^ а б c d Морено и др., 2015, стр.6.
- ^ Эрнандес Пардо и др., 2009, стр.66.
- ^ Афанадор, 2009, стр.38.
- ^ Родригес и Лондоньо, 2002, стр.177
- ^ а б Каррильо Брисеньо и др., 2016, стр.78.
- ^ а б c d Хенди и др., 2015, стр.47.
- ^ Климатические данные Урибия
- ^ (на испанском) Альта Гуахира, дос аньос син ллувия - El Espectador
- ^ а б c Plancha 3, 2008 г.
- ^ Plancha 5, 2008 г.
- ^ Plancha 10, 2008 г.
- ^ а б Планча 10бис, 2008 г.
- ^ Морено и др., 2015, стр.7.
- ^ Возраст наземных млекопитающих в Южной Америке в Окаменелости.org
- ^ Морено и др., 2015, стр. 32-34.
- ^ Морено и др., 2015, стр. 27-32.
- ^ Морено и др., 2015, стр. 18-27.
- ^ Родригес и Лондоньо, 2002, стр.132.
- ^ Родригес и Лондоньо, 2002, стр.128.
- ^ а б Морено и др., 2015, стр.23.
- ^ Морено и др., 2015, стр.27.
- ^ Морено и др., 2015, стр.31.
- ^ Морено и др., 2015, стр.32.
- ^ а б Морено и др., 2015, стр.33.
- ^ Морено и др., 2015, стр.34.
- ^ а б Эрнандес Пардо и др., 2009, стр.62.
- ^ Wing et al., 2009, с.18629.
- ^ а б c Эрнандес Пардо и др., 2009 г., стр. 60
- ^ Эрнандес Пардо и др., 2009, стр.48.
- ^ Эрнандес Пардо и др., 2009, стр.23.
- ^ Эрнандес Пардо и др., 2009, стр.50
- ^ Де Порта, 2003, стр.191
- ^ Эрнандес Пардо и др., 2009, стр.69.
- ^ Эрнандес Пардо и др., 2009, стр.122.
- ^ Эрнандес Пардо и др., 2009, стр.28.
- ^ Хенди и др., 2015, стр.72.
- ^ Каррильо Брисеньо и др., 2016, стр.92
- ^ Кадена и Харамилло, 2015, стр.199
- ^ Морено Бернал, 2014, стр.41
- ^ Морено Бернал, 2014, стр.42
- ^ Амсон и др., 2016, стр.16.
- ^ а б Перес и др., 2017
- ^ а б c d е ж грамм час я Морено и др., 2015, стр.35.
- ^ а б Амсон и др., 2016, стр.3
- ^ Амсон и др., 2016, стр.4.
- ^ а б Амсон и др., 2016, стр.7.
- ^ Амсон и др., 2016, стр.8
- ^ Амсон и др., 2016, стр.9
- ^ Амсон и др., 2016, стр.12.
- ^ Амсон и др., 2016, стр.15.
- ^ Полицейский участок 470062 в Окаменелости.org
- ^ Карилло, 2018, с.142
- ^ Суарес и др., 2015, стр.4.
- ^ Морено Бернал, 2014, стр.7
- ^ Морено Бернал, 2014, стр.11
- ^ Морено Бернал, 2014, стр.13
- ^ Морено Бернал, 2014, стр.15
- ^ Морено Бернал, 2014, стр.19
- ^ Морено Бернал, 2014, стр.26
- ^ Морено Бернал, 2014, стр.29.
- ^ Морено Бернал, 2014, стр.30
- ^ Кадена и Харамилло, 2015, с.190
- ^ а б Кадена и Харамилло, 2015, стр.194
- ^ Кадена и Харамилло, 2015, стр.196
- ^ Морено и др., 2015, стр.29.
- ^ а б Хенди и др., 2015, стр.52.
- ^ а б Хенди и др., 2015, стр.53.
- ^ а б Хенди и др., 2015, стр. 54
- ^ Каррильо Брисеньо и др., 2016, стр.91
- ^ а б Хенди и др., 2015, стр.50.
- ^ Формация Макарао
Библиография
- Афанадор, Ф. 2009. Caracterización físico-biótica del litoral del depamento de La Guajira, 33–66. Проверено 8 августа 2017 г.
- Амсон, Эли; Хуан Д. Каррильо, и Карлос Харамилло. 2016. Неогеновые сообщества ленивцев (Mammalia, Pilosa) в бассейне Кокинетас (Ла-Гуахира, Колумбия): последствия для великого американского биотического обмена. Палеонтология _. 1–20. По состоянию на 31 марта 2017 г.
- Кадена, Эдвин, и Карлос Харамилло. 2015. Черепахи раннего и среднего миоцена из самой северной оконечности Южной Америки: гигантские тестудиниды, хелиды и подокнемидиды из формации Кастильетес, Колумбия. Амегиниана 52. 188–203. Проверено 8 августа 2017 г.
- Каррильо Брисеньо, Хорхе Д.; Тодорис Аргириу; Владимир Запата; Рене Киндлиманн, и Карлос Харамилло. 2016. Новое раннемиоценовое (аквитанское) скопление Elasmobranchii с полуострова Ла Гуахира, Колумбия. Амегиниана 53. 77–99. Проверено 8 августа 2017 г.
- Карилло Санчес, Хуан Давид. 2018. Систематика коренных копытных животных Южной Америки и неогеновая эволюция млекопитающих Северной Южной Америки (кандидатская диссертация), 1–285. Цюрихский университет. Проверено 15 мая 2018 г.
- Хенди, Остин Дж. У.; Дуглас С. Джонс; Федерико Морено; Владимир Запата, и Карлос Харамилло. 2015. Неогеновые моллюски, мелководные морские палеосреды и хроностратиграфия полуострова Гуахира, Колумбия. Швейцарский журнал палеонтологии 134. 45–75. Проверено 8 августа 2017 г.
- Эрнандес Пардо, Орландо; Хосе Мария Харамильо; Маурисио Парра; Армандо Салазар; Раймонд Донелик, и Астрид Бландон. 2009. Reconstrucción de la Historia termal en el piedemonte occidental de la Serranía del Perijá entre Codazzi y La Jagua de Ibirico - Cuenca de Cesar-Ranchería, 1–85. Национальный университет Колумбии & ANH. Проверено 14 июня 2017 г.
- Морено, Ф.; A.J.W. Хенди; Л. Кирос; Н. Ойос; Д.С. Джонс; В. Сапата; С. Сапата; Г.А. Баллен, и Эдвин Кадена, A.L. Cárdenas, J.D. Carrillo Briceño, J.D. Carrillo, D. Delgado Sierra, J. Escobar, J.I. Мартинес, К. Мартинес, К. Монтес, Х. Морено, Н. Перес, Р. Санчес, К. Суарес, М.С. Вальехо Пареха, К. Харамильо. 2015. Пересмотренная стратиграфия пластов неогена в бассейне Кокинетас, Ла-Гуахира, Колумбия. Швейцарский журнал палеонтологии 134. 5–43. Проверено 8 августа 2017 г.
- Морено Берналь, Хорхе В.. 2014. Ископаемые крокодилы с полуострова Высокий Гуахира в Колумбии и история неогенового крокодилового разнообразия в тропической Южной Америке, 1–66. Университет Небраски. Проверено 8 августа 2017 г.
- Перес, Мария Э.; Мария К. Вальехо Пареха; Хуан Д. Каррильо, и Карлос Харамилло. 2017. Капибара нового плиоцена (Rodentia, Caviidae) из Северной Южной Америки (Гуахира, Колумбия) и ее значение для великого обмена биотиками Америки. Журнал эволюции млекопитающих 24. 111–125. Проверено 4 апреля 2017 г.
- Де Порта, Хайме. 2003. La formación del istmo de Panamá - su incidencia en Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales XXVII. 191–216.
- Родригес, Габриэль, и Ана Кристина Лондоньо. 2002. Геологическая карта департамента Ла-Гуахира - 1: 250 000, 1–259. ИНГЕОМИНЫ. Проверено 8 августа 2017 г.
- Варгас, Карлос А.; Луис А. Монтес, и Карлос Ортега. 2012. Geología estructural y estratigrafía del área Majayura (Guajira). Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 36. 385–398.
- Крыло, Скотт Л.; Фабиани Эррера; Карлос А. Харамилло; Каролина Гомес Наварро; Питер Уилф, и Конрад К. Лабандейра. 2009. Окаменелости позднего палеоцена из формации Серрехон, Колумбия ((sic)), являются самым ранним упоминанием о неотропических тропических лесах.. Труды Национальная Академия Наук 106. 18627–18632. Проверено 14 июня 2017 г.
Карты
- Сулуага, Карлос А.; Альберто Очоа; Карлос А. Муньос; Камило Э. Дорадо; Наталия М. Герреро; Ана М. Мартинес; Паула А. Медина; Эдгар Ф. Окампо, и Алехандро Пинилья и Паула А. Риос, Бибиана П. Родригес, Эдвард А. Салазар, Владимир Л. Сапата. 2008. Планча 3 - Пуэрто-Эстрелла - 1: 100 000, 1. ИНГЕОМИНЫ. По состоянию на 06.06.2017.
- Сулуага, Карлос А.; Альберто Очоа; Карлос А. Муньос; Камило Э. Дорадо; Наталия М. Герреро; Ана М. Мартинес; Паула А. Медина; Эдгар Ф. Окампо, и Алехандро Пинилья и Паула А. Риос, Бибиана П. Родригес, Эдвард А. Салазар, Владимир Л. Сапата. 2008. Планча 5 - Баия Портете - 1: 100 000, 1. ИНГЕОМИНЫ. По состоянию на 06.06.2017.
- Сулуага, Карлос А.; Альберто Очоа; Карлос А. Муньос; Камило Э. Дорадо; Наталия М. Герреро; Ана М. Мартинес; Паула А. Медина; Эдгар Ф. Окампо, и Алехандро Пинилья и Паула А. Риос, Бибиана П. Родригес, Эдвард А. Салазар, Владимир Л. Сапата. 2008. Планча 10 - Ранчо Гранде - 1: 100 000, 1. ИНГЕОМИНЫ. По состоянию на 06.06.2017.
- Сулуага, Карлос А.; Альберто Очоа; Карлос А. Муньос; Камило Э. Дорадо; Наталия М. Герреро; Ана М. Мартинес; Паула А. Медина; Эдгар Ф. Окампо, и Алехандро Пинилья и Паула А. Риос, Бибиана П. Родригес, Эдвард А. Салазар, Владимир Л. Сапата. 2008. Планча 10бис - Ранчо Гранде - 1: 100 000, 1. ИНГЕОМИНЫ. По состоянию на 06.06.2017.
дальнейшее чтение
- Балли, А., и С. Снельсон. 1980. Царства проседания. Мемуары Канадского общества нефтяной геологии 6. 9–94.
- Кингстон, Д.; C.P. Dishroon, и П.А. Уильямс. 1983. Глобальная система классификации бассейнов. Бюллетень AAPG 67. 2175–2193. Проверено 23 июня 2017 г.
- Клемме, Х.. 1980. Нефтяные бассейны - классификации и характеристики. Журнал нефтяной геологии 3. 187–207. Проверено 23 июня 2017 г.
Осадочные бассейны Колумбии | ||||||||
|---|---|---|---|---|---|---|---|---|
| На берегу |
|  | ||||||
| Офшор | ||||||||
| Источник | • Барреро, Дарио; Андрес Пардо; Карлос А. Варгас, и Хуан Ф. Мартинес. 2007. Колумбийские осадочные бассейны: номенклатура, границы и нефтегазовая геология, новое предложение, 1–92. ANH. | |||||||