Кортико-базальные ганглии-таламо-кортикальная петля - Cortico-basal ganglia-thalamo-cortical loop
Эта статья может быть слишком техническим для большинства читателей, чтобы понять. Пожалуйста помогите улучшить это к Сделайте это понятным для неспециалистов, не снимая технических деталей. (Апрель 2019) (Узнайте, как и когда удалить этот шаблон сообщения) |
| Кортико-базальные ганглии-таламо-кортикальная петля | |
|---|---|
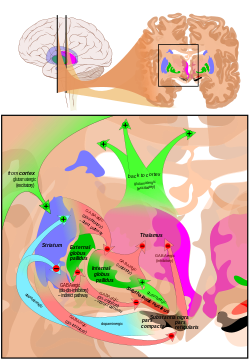 Связи базальный ганглий. | |
| Подробности | |
| Часть | Головной мозг |
| Анатомические термины нейроанатомии | |
В кортико-базальные ганглии-таламо-кортикальная петля (CBGTC петля) это система из нейронные цепи в мозг. Петля включает в себя связи между кора, то базальный ганглий, то таламус, и обратно в кору. Это особенно актуально для гиперкинетический и гипокинетический двигательные расстройства, Такие как болезнь Паркинсона и болезнь Хантингтона,[1] а также психические расстройства контроля, например Синдром дефицита внимания и гиперактивности (СДВГ),[2] обсессивно-компульсивное расстройство (ОКР),[3] и синдром Туретта.[4]
Цикл CBGTC в основном состоит из модулирующих дофаминергические проекции от pars compacta из черная субстанция, и вентральная тегментальная область а также возбуждающий глутаматергический прогнозы из кора к полосатое тело, где эти проекции образуют синапсы с возбуждающими и тормозными путями, которые возвращаются в кору. Изначально петля была предложена как часть модели базальный ганглий называется модель параллельной обработки, который подвергся критике и преобразован в другую модель, названную центральная модель объемного звучания.[5]
Современные схемы организации характеризуют взаимодействия кортико-базальных ганглиев как сегрегированные параллельные процессы, означающие, что в базальных ганглиях существует небольшая конвергенция отдельных кортикальных областей. Считается, что это объясняет топографически организованную функциональность полосатого тела.[4] Стриатум организован по ростро-каудальной оси, причем ростральная часть скорлупа и хвостатый обслуживает ассоциативные и когнитивные функции, а каудальные области - сенсомоторную функцию.[6] Иногда, когда выраженной мишенью является полосатое тело, петлю называют кортико-полосато-таламо-кортикальная петля.[7]
Нейроанатомия

Двумя основными входными структурами схемы являются полосатое тело и субталамическое ядро (СТН). Стриатум получает входы как от коры, так и от pars compacta из черная субстанция (SNc), в то время как STN получает только корковые сигналы.
Из полосатого тела отходят два пути. Один путь называется непрямой (или нет) путь и является тормозящим. Это проецирует и сдерживает бледный глобус внешний (GPe), что приводит к растормаживанию бледный глобус внутренний (GPi), что приводит к угнетению таламуса. Этот путь также, в результате ингибирования GPe, растормаживает субталамическое ядро, что приводит к возбуждению GPi и, следовательно, к ингибированию таламуса.
Второй путь называется прямой (или идущий) путь и возбуждает. Этот путь ингибирует GPi, что приводит к растормаживанию таламуса. Прямой путь в основном состоит из моносинаптических связей, управляемых дофаминовый рецептор D1, аденозиновый рецептор A1, и мускариновый рецептор ацетилхолина M4, в то время как косвенный путь основан на связях, управляемых дофаминовый рецептор D2, аденозиновый рецептор A2A, и мускариновый рецептор ацетилхолина M1.[1][8]
Параллельные петли CBGTC были разделены в соответствии с функциями связанных областей коры. Одна схема включает разделение на лимбическую и моторную петли, при этом моторные петли содержат непрямые и прямые пути, которые, в свою очередь, связаны с лимбической петлей, которая выступает в вентральное полосатое тело.[9] Петля также подразделяется на лимбическую, ассоциативную, глазодвигательную и моторную.[4] объяснить роль дофамина в базальных ганглиях на мотивационные состояния.[10] Разделение на пять петель, основанное на основных корковых мишенях, было описано следующим образом:[11]
- Цепь двигателя, возникающая в дополнительная моторная зона, моторная кора, и соматосенсорная кора, который, в свою очередь, проецируется на скорлупу, которая проецируется на вентролатеральный GPi и каудолатеральный SNr, прежде чем вернуться в кору через ventralis lateralis pars oralis и ventralis lateralis pars medialis.
- Глазодвигательный контур, возникающий в лобные поля глаз выступает к телу хвостатого тела и возвращается через каудальный дорсомедиальный GPi / вентромедиальный SNr, а затем латеральную вентральную переднюю pars magnocellularis и medialis dorsalis pars paralarnellaris.
- Дорсолатеральный префронтальный контур с проекциями из dlPFC и заднего теменная кора, которая проецируется на дорсолатеральную головку хвостатого тела, которая в свою очередь проецируется на латеральный дорсомедиальный GPi / ростролатеральный SNr, который проецируется на ventralis anterior pars parvocellularis и medialis dorsalis pars parvocellularis.
- Латеральный орбитофронтальный контур проходит к вентромедиальной хвостатой головке, которая проходит через медиальный дорсомедиальный GPi / ростромедиальный SNr к медиальной вентральной медиальной части pars magnocellularis / medialis dorsalis pars magnocellularis.
- Передняя поясная извилина, которая включает проекции от ACC к вентральному полосатому телу через ростолатеральный GPi / VP / ростродорсальный SNr, который возвращается через заднемедиальную медиалис дорсалис.
Проблема, связанная с текущей анатомией схемы, заключается в том, что временная задержка между прямым и косвенным путями должна приводить к тому, что эта схема не работает. Чтобы преодолеть это, гипотеза центрального окружения постулирует, что гиперпрямой путь из коры будет препятствовать другим входам, кроме одного сфокусированного кортикального входа. Однако время активности базальных ганглиев и момента конечностей, а также исследования повреждений не подтверждают эту гипотезу.[12]
Функция
Были предложены две модели, чтобы объяснить, как выбираются действия в базальных ганглиях. Модель «актер-критик» предполагает, что действия генерируются и оцениваются «критиком» в брюшном полосатом теле, в то время как действия выполняются «актером» в дорсальном полосатом теле. Другая модель предполагает, что базальные ганглии действуют как механизм отбора, при котором действия генерируются в коре и выбираются базальными ганглиями в зависимости от контекста.[13] Цикл CBGTC также участвует в дисконтировании вознаграждения, когда увольнение увеличивается с неожиданным или большим, чем ожидалось, вознаграждением.[2] Один обзор подтвердил идею о том, что кора головного мозга участвует в обучающих действиях независимо от их результата, в то время как базальные ганглии участвуют в выборе подходящих действий на основе ассоциативного обучения методом проб и ошибок на основе вознаграждения.[14]
Роль в болезни
Петля CBGTC участвует во многих заболеваниях. Например, в болезнь Паркинсона, считается, что дегенерация дофаминергических нейронов, ведущая к снижению активности возбудительного пути, приводит к гипокинезии,[15] И в болезнь Хантингтона считается, что дегенерация ГАМКергических нейронов, управляющих тормозным путем, приводит к резким движениям тела.[2] Совместная дегенерация лимбических проекций вместе с моторными проекциями может приводить ко многим психиатрическим симптомам этих преимущественно моторных заболеваний.[9] При ОКР петля может быть дисфункциональной, с дисбалансом между непрямыми и прямыми путями, что приводит к нежелательному мысли, застрять".[3] Считается, что при СДВГ снижение тонической дофаминергической сигнализации, приводящее к чрезмерному дисконтированию отложенных вознаграждений, приводит к снижению внимания.[2]
Исследование
Петля CBGTC была изучена в отношении сознание выбор действия по отношению к другим схемам и в контексте памяти и познания.[16][17] Модель петли CBGTC подверглась критике как чрезмерно упрощенная и слишком жестко применяемая, учитывая свидетельства анатомического и функционального перекрытия и взаимодействия между прямыми и непрямыми путями.[18] Петля также исследовалась в контексте глубокая стимуляция мозга.[16] По состоянию на 2013 год велись интенсивные дебаты по поводу деления цепи, взаимодействия путей, количества путей и общей анатомии.[17]
Рекомендации
- ^ а б Силкис, И. (1 января 2001 г.). «Кортико-базальные ганглии-таламокортикальный контур с синаптической пластичностью. II. Механизм синергетической модуляции таламической активности через прямые и непрямые пути через базальные ганглии». Биосистемы. 59 (1): 7–14. Дои:10.1016 / S0303-2647 (00) 00135-0. ISSN 0303-2647. PMID 11226622.
- ^ а б c d Maia, Tiago V .; Франк, Майкл Дж. (15 января 2017 г.). «От моделей обучения с подкреплением базальных ганглиев до патофизиологии психиатрических и неврологических расстройств». Природа Неврология. 14 (2): 154–162. Дои:10.1038 / № 2723. ISSN 1097-6256. ЧВК 4408000. PMID 21270784.
- ^ а б Maia, Tiago V .; Куни, Ребекка Э .; Петерсон, Брэдли С. (1 января 2008 г.). «Нейронные основы обсессивно-компульсивного расстройства у детей и взрослых». Развитие и психопатология. 20 (4): 1251–1283. Дои:10.1017 / S0954579408000606. ISSN 0954-5794. ЧВК 3079445. PMID 18838041.
- ^ а б c Делонг, Махлон; Вичманн, Томас (15 января 2017 г.). «Изменение взглядов на контуры базальных ганглиев и расстройства контуров». Клиническая ЭЭГ и нейробиология. 41 (2): 61–67. Дои:10.1177/155005941004100204. ISSN 1550-0594. ЧВК 4305332. PMID 20521487.
- ^ Уттер, Эми А .; Бассо, Микеле А. (1 января 2008 г.). «Базальные ганглии: обзор цепей и функций». Неврология и биоповеденческие обзоры. 32 (3): 333–342. Дои:10.1016 / j.neubiorev.2006.11.003. ISSN 0149-7634. PMID 17202023.
- ^ Ким, HF; Хикосака, О. (июль 2015 г.). «Параллельные контуры базальных ганглиев для произвольного и автоматического поведения для достижения награды». Мозг: журнал неврологии. 138 (Pt 7): 1776–800. Дои:10.1093 / мозг / awv134. ЧВК 4492412. PMID 25981958.
- ^ Fettes, P .; Schulze, L .; Даунар, Дж. (2017). "Кортико-стриатально-таламические петли орбитофронтальной коры: многообещающие терапевтические цели при психических заболеваниях". Границы системной нейробиологии. 11: 25. Дои:10.3389 / fnsys.2017.00025. ЧВК 5406748. PMID 28496402.
- ^ Родитель, А .; Хазрати, Л. Н. (1 января 1995 г.). «Функциональная анатомия базальных ганглиев. I. Кортико-базальные ганглии-таламо-кортикальная петля». Исследование мозга. Обзоры исследований мозга. 20 (1): 91–127. Дои:10.1016 / 0165-0173 (94) 00007-C. PMID 7711769.
- ^ а б Н.Ф., Мехраби; Мальвиндар, Сингх-Бейнс; Генри, Вальдфогель; Ричард, Фаулл (21 июля 2016 г.). «Кортико-базальные ганглии при болезни Хантингтона». Цитировать журнал требует
| журнал =(помощь) - ^ Икемото, Сатоши; Ян, Чен; Тан, Аарон (1 сентября 2015 г.). «Петли цепи базальных ганглиев, дофамин и мотивация: обзор и исследование». Поведенческие исследования мозга. 290: 17–31. Дои:10.1016 / j.bbr.2015.04.018. ЧВК 4447603. PMID 25907747.
- ^ Сквайр, Ларри (2013). Фундаментальная нейробиология (4-е изд.). Амстердам: Elsevier / Academic Press. п. 728. ISBN 9780123858702.
- ^ Делонг, Махлон; Вичманн, Томас (15 января 2017 г.). «Обновленная информация о моделях функции и дисфункции базальных ганглиев». Паркинсонизм и связанные с ним расстройства. 15 (Приложение 3): S237 – S240. Дои:10.1016 / S1353-8020 (09) 70822-3. ISSN 1353-8020. ЧВК 4275124. PMID 20082999.
- ^ Redgrave, P .; Прескотт, T.J .; Герни, К. (апрель 1999 г.). «Базальные ганглии: позвоночное решение проблемы отбора?». Неврология. 89 (4): 1009–1023. CiteSeerX 10.1.1.32.4792. Дои:10.1016 / S0306-4522 (98) 00319-4. PMID 10362291.
- ^ Хели, Себастьян; Ell, Shawn W .; Эшби, Ф. Грегори (1 марта 2015 г.). «Изучение надежных кортико-корковых ассоциаций с базальными ганглиями: интегративный обзор». Кора. 64: 123–135. Дои:10.1016 / j.cortex.2014.10.011. ISSN 1973-8102. PMID 25461713.
- ^ Lanciego, José L .; Люкин, Наташа; Обесо, Хосе А. (15 января 2017 г.). «Функциональная нейроанатомия базальных ганглиев». Перспективы Колд-Спринг-Харбор в медицине. 2 (12): a009621. Дои:10.1101 / cshperspect.a009621. ISSN 2157-1422. ЧВК 3543080. PMID 23071379.
- ^ а б Бриттен, JS; Шаротт, А; Браун, П. (июнь 2014 г.). «Максимумы и минимумы бета-активности в петлях кортико-базальных ганглиев». Европейский журнал нейробиологии. 39 (11): 1951–9. Дои:10.1111 / ejn.12574. ЧВК 4285950. PMID 24890470.
- ^ а б Шролл, Хеннинг; Хамкер, Фред Х. (30 декабря 2013 г.). «Вычислительные модели функций проводящих путей базальных ганглиев: основное внимание уделяется функциональной нейроанатомии». Границы системной нейробиологии. 7: 122. Дои:10.3389 / fnsys.2013.00122. ЧВК 3874581. PMID 24416002.
- ^ Калабрези, Паоло; Пиккони, Барбара; Тоцци, Алессандро; Гильери, Вероника; Филиппо, Массимилиано Ди (1 августа 2014 г.). «Прямые и непрямые пути базальных ганглиев: критическая переоценка». Природа Неврология. 17 (8): 1022–1030. Дои:10.1038 / № 3743. PMID 25065439.