Хвостатое ядро - Caudate nucleus
| Хвостатое ядро | |
|---|---|
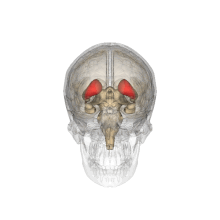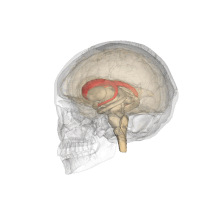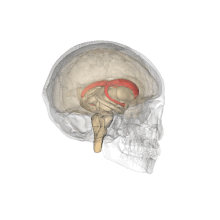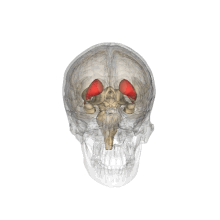
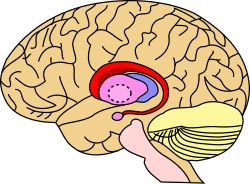 Хвостатое ядро (красным) показано в головном мозге | |
 Поперечный разрез мозга (горизонтальный разрез), базальный ганглий синий | |
| Подробности | |
| Часть | спинное полосатое тело |
| Идентификаторы | |
| латинский | хвостовое ядро |
| MeSH | D002421 |
| NeuroNames | 226 |
| НейроЛекс Я БЫ | birnlex_1373 |
| TA98 | A14.1.09.502 |
| TA2 | 5561 |
| FMA | 61833 |
| Анатомические термины нейроанатомии | |
В хвостатое ядро одна из структур, составляющих полосатое тело, который является составной частью базальный ганглий.[1] Хвостатое ядро долгое время было связано с моторными процессами из-за его роли в болезнь Паркинсона,[2][требуется разъяснение ][3] он также играет важную роль в различных других немоторных функциях, включая процедурное обучение,[4] ассоциативное обучение[5] и тормозящий контроль действия,[6] среди других функций. Хвостатое тело также является одной из структур головного мозга, составляющих система вознаграждений и функционирует как часть кортик –базальный ганглий –таламический петля.[1]
Структура
Вместе с скорлупа, хвостатое спинное полосатое тело, который считается единой функциональной структурой; анатомически он разделен большим трактом белого вещества, внутренняя капсула, поэтому его иногда также называют двумя структурами: медиальным дорсальным полосатым телом (хвостатым) и латеральным дорсальным полосатым телом (скорлупа). В этом ключе, эти два функционально отличаются не в результате структурных различий, а просто из-за топографического распределения функций.
Хвостатые ядра расположены недалеко от центра мозга, сидят верхом на таламус. Внутри каждого из них есть хвостатое ядро. полушарие мозга. По отдельности они напоминают С-образную конструкцию с более широкой «головой» (caput на латыни) спереди, сужаясь к «телу» (корпус) и «хвост» (кауда). Иногда часть хвостатого ядра называют «коленом» (Genu).[7] Хвостатая головка получает кровоснабжение от лентикулостриарной артерии, а хвост хвостатого тела получает кровоснабжение от передней хориоидальной артерии.[8]

Голова и тело хвостатого ядра образуют часть дна переднего рога боковой желудочек. После того, как тело ненадолго перемещается к затылку, хвост изгибается назад к передней части, образуя крышу нижнего рога бокового желудочка. Это означает, что коронал (в плоскости, параллельной лицо ) часть, которая прорезает хвост, также пересекает тело и голову хвостатого ядра.
Нейрохимия
Хвостатое тело сильно иннервируется дофаминергические нейроны которые происходят из черная субстанция pars compacta (SNc). SNc находится в средний мозг и содержит выступы клеток на хвостатое и скорлупа, используя нейротрансмиттер дофамин.[9] Также есть дополнительные входы от различных ассоциация коры.
Двигательные функции
Пространственная мнемоническая обработка
Хвостатое ядро объединяет пространственную информацию с формулировкой моторного поведения. Избирательное нарушение пространственная рабочая память по предметам с болезнь Паркинсона и знание влияния болезни на количество дофамин поставляется в полосатое тело связали хвостатое ядро с пространственной и непространственной мнемонической обработкой. Пространственно-зависимая двигательная подготовка была связана с хвостатым ядром через событие, связанное с фМРТ методы анализа. Было продемонстрировано, что активность хвостатого ядра выше при выполнении задач, требующих пространственной и моторной памяти, чем при выполнении непространственных задач.[10]Конкретно, пространственная рабочая память активность наблюдалась через фМРТ исследования отсроченного распознавания, чтобы быть больше в хвостатом ядре, когда активность непосредственно предшествовала двигательной реакции. Эти результаты указывают на то, что хвостатое ядро может участвовать в кодировании двигательной реакции. Имея это в виду, хвостатое ядро могло бы участвовать в рекрутировании двигательной системы для поддержки производительности рабочей памяти путем посредничества сенсомоторных преобразований.[11]
Направленные движения
Хвостатое ядро играет важную роль в осанке тела и конечностей, а также в скорости и точности направленных движений. После удаления хвостатых ядер у животных из семейства кошачьих наблюдались нарушения осанки и точности при выполнении задач на использование лап. После частичного удаления ядер у кошек наблюдалась задержка в начале работоспособности и необходимость постоянно менять положение тела.[12]
После применения кокаин по отношению к хвостатому ядру и образовавшимся в результате повреждениям у обезьян наблюдалось "прыжок или движение вперед". Из-за связи с повреждением хвостатого ядра это движение демонстрирует тормозящую природу хвостатого ядра. «Моторное освобождение», наблюдаемое в результате этой процедуры, указывает на то, что хвостатое ядро подавляет тенденцию животного двигаться вперед без сопротивления.[13]
Когнитивные функции
Целенаправленное действие
Обзор нейровизуализационных исследований, анатомических исследований связности хвостатого ядра и поведенческих исследований показывает роль хвостатого тела в исполнительном функционировании. Исследование пациентов с болезнью Паркинсона (см. Ниже) также может способствовать увеличению количества доказательств.
Двусторонний подход к нейровизуализации (включая ДОМАШНИЙ ПИТОМЕЦ и фМРТ ) и анатомические исследования выявляют сильную взаимосвязь между хвостатым и кортикальным слоями, связанную с исполнительным функционированием: «неинвазивные измерения анатомической и функциональной связи у людей демонстрируют четкую связь между хвостатым и исполнительным лобными областями».[14]
Между тем, поведенческие исследования предоставляют еще один уровень аргументации: недавние исследования показывают, что хвостатое тело имеет фундаментальное значение для целенаправленных действий, то есть «выбора поведения, основанного на меняющихся ценностях целей и знания о том, какие действия приводят к каким результатам. . "[14] В одном из таких исследований у крыс были рычаги, которые запускали высвобождение раствора со вкусом корицы. После того, как крысы научились нажимать на рычаг, исследователи изменили ценность результата (крыс учили не любить аромат либо из-за того, что им давали слишком много аромата, либо из-за того, что крысам становилось плохо после питья раствора) и эффекты. наблюдались. Нормальные крысы реже нажимали на рычаг, в то время как крысы с поражениями хвостатого тела не подавляли поведение так эффективно. Таким образом, исследование демонстрирует связь между хвостатым и целенаправленным поведением; крысы с поврежденными хвостатыми ядрами испытывали трудности с оценкой меняющейся ценности результата.[14] В исследовании поведения людей в 2003 году аналогичный процесс был повторен, но на этот раз решение заключалось в том, доверять ли другому человеку, когда на карту поставлены деньги.[15] Хотя здесь выбор был гораздо более сложным - испытуемых не просто просили нажать на рычаг, а нужно было взвесить множество различных факторов, - сутью исследования по-прежнему оставался поведенческий отбор, основанный на изменении значений результатов.
Короче говоря, нейровизуализация и анатомические исследования подтверждают утверждение о том, что хвостатое тело играет роль в исполнительном функционировании, в то время как поведенческие исследования углубляют наше понимание способов, которыми хвостатое тело направляет некоторые из наших процессов принятия решений.
объем памяти
Подкорковая петля дорсально-префронтальной коры, вовлекающая хвостатое ядро, была связана с дефицитом рабочей памяти, особенно в шизофреник пациенты. Функциональная визуализация показала активацию этой подкорковой петли во время задач рабочей памяти у приматов и здоровых людей. Хвостатое тело может быть связано с дефицитом рабочей памяти еще до начала болезни. Было обнаружено, что объем хвостатого ядра обратно пропорционален персеверативный ошибки в задачах пространственной рабочей памяти.[16][17]
В миндалина посылает прямые проекции в хвостатое ядро. Оба миндалина и хвостатое ядро имеют прямые и косвенные проекции на гиппокамп. Влияние миндалина обработки памяти в хвостатом ядре было продемонстрировано с обнаружением, что повреждения, вовлекающие связи между этими двумя структурами, "блокируют эффекты улучшения памяти оксотреморин вливается в хвостатое ядро ". В исследовании с участием крыс, прошедших обучение в водном лабиринте, было обнаружено, что хвостатое ядро улучшает память о тренировках с визуальными подсказками после амфетамин была введена после тренировки в хвостатое.[18]
Учусь
В исследовании 2005 года испытуемых просили научиться классифицировать визуальные стимулы, классифицируя изображения и получая отзывы о своих ответах. Активность, связанная с успешным классификационным обучением (правильная категоризация), была сосредоточена на теле и хвосте хвостатого тела, в то время как деятельность, связанная с обработкой обратной связи (результат неправильной категоризации), была сосредоточена на голове хвостатого тела.[19]
Спать
Двусторонние поражения в головке хвостатого ядра у кошек коррелировали с уменьшением продолжительности глубокого медленный сон во время цикла сна-бодрствования. При уменьшении общего объема глубокого медленноволнового сна переход краткосрочная память к Долгосрочная память также могут быть затронуты отрицательно.[20]Однако влияние удаления хвостатого ядра на характер бодрствования кошек не было постоянным. Нормализация была обнаружена через три месяца после появления хвостатых ядер. абляция. Это открытие могло быть связано с взаимосвязанным характером ролей хвостатого ядра и лобная кора в контроле уровней Центральная нервная система активация. У кошек с удаленным хвостатым концом, хотя и постоянно гиперактивно, наблюдалось значительное снижение быстрое движение глаз во сне (REMS) всего около двух месяцев. Однако у афронтальных кошек наблюдалось постоянное уменьшение времени REMS и только временный период гиперактивности.[21]
В отличие от ассоциаций между «глубоким», быстрым сном и хвостатым ядром, исследование, включающее ЭЭГ и фМРТ Измерения во время циклов сна человека показали, что хвостатое ядро демонстрирует пониженную активность во время не-REM-сна на всех стадиях сна.[22] Кроме того, исследования объема хвостатых ядер человека в врожденный синдром центральной гиповентиляции (CCHS) субъекты установили корреляцию между CCHS и значительным уменьшением объема левого и правого хвостатого ядра. CCHS - это генетическое заболевание, которое влияет на цикл сна из-за пониженного желания дышать. Таким образом, предполагается, что хвостатое ядро играет роль в циклах сна человека.[23]
Эмоции
Хвостатое ядро участвует в реакции на визуальную красоту и считается одним из «нейронных коррелятов романтической любви».[24][25]
Поведение подхода-привязанности и аффект также контролируются хвостатым ядром. Кошки с двусторонним удалением хвостатых ядер настойчиво приближались и следовали за объектами, пытаясь связаться с целью, демонстрируя при этом дружелюбный настрой, вызывая топот передних конечностей и мурлыканье. Величина поведенческих реакций коррелировала со степенью удаления ядер. Сообщения о людях с избирательным повреждением хвостатого ядра показывают одностороннее повреждение хвостатого ядра, приводящее к потере инстинкта, обсессивно-компульсивное расстройство, привязанный к стимулу персеверативный поведение и гиперактивность. Большинство этих недостатков можно классифицировать как связанные с поведением приближения и привязанности, от приближения к цели до романтической любви.[12]
Язык
Исследования с помощью нейровизуализации показывают, что люди, которые могут общаться на нескольких языках, активируют одни и те же области мозга независимо от языка. Публикация 2006 года исследует это явление и определяет хвостатое тело как центр языкового контроля. Пожалуй, наиболее показательный случай наблюдался у трехъязычного субъекта с поражением хвостатого ядра. Пациентка сохранила понимание речи на трех ее языках, но когда ее попросили воспроизвести язык, она невольно переключилась между тремя языками. Вкратце, «эти и другие данные, полученные у двуязычных пациентов, предполагают, что левое хвостатое тело требуется для мониторинга и контроля лексических и языковых альтернатив в производственных задачах».[26][27]
Локальные деформации формы медиальной поверхности хвостатого тела коррелировали с речевой способностью к обучению у женщин и количеством ошибок настойчивости при выполнении задач рабочей памяти на пространственную и вербальную беглость у мужчин. В частности, больший объем хвостатого ядра был связан с лучшей вербальной беглостью.[16]
Неврологическое исследование глоссолалия показали значительное снижение активности в левом хвостатом ядре во время глоссолалии по сравнению с пением на английском языке.[28]
Контроль порога
Мозг содержит большие коллекции нейронов, взаимно связанных между собой возбуждающие синапсы, образуя большую сеть элементов с положительный отзыв. Трудно понять, как такая система может работать без какого-либо механизма предотвращения взрывной активации. Есть некоторые косвенные доказательства[29] что хвостатое может выполнять эту регулирующую роль, измеряя общую активность кора головного мозга и контроль пороговый потенциал.
Клиническое значение
Болезнь Альцгеймера
Исследование 2013 г. показало связь между Болезнь Альцгеймера пациенты и хвостатое ядро. Изображения МРТ использовались для оценки объема хвостатых ядер у пациентов с болезнью Альцгеймера и здоровых добровольцев. Исследование показало «значительное уменьшение объема хвостатого ядра» у пациентов с болезнью Альцгеймера по сравнению с нормальными добровольцами. Хотя корреляция не указывает на причинно-следственную связь, это открытие может иметь значение для ранней диагностики.[30]
болезнь Паркинсона
болезнь Паркинсона вероятно, наиболее изученное заболевание базальных ганглиев. Пациенты с этим прогрессирующим нейродегенеративным расстройством часто сначала испытывают симптомы, связанные с движением (три наиболее распространенных - тремор в покое, мышечная ригидность и акатизия ), которые позже сочетаются с различными когнитивными нарушениями, включая деменцию.[31] Болезнь Паркинсона истощает дофаминергические нейроны в нигростриатном тракте, пути дофамина, который связан с головкой хвостатого тела. Таким образом, многие исследования коррелировали потерю дофаминергических нейронов, которые посылают аксоны в хвостатое ядро, и степенью деменции у пациентов с болезнью Паркинсона.[14] И хотя связь между хвостатым отростком и двигательной недостаточностью Паркинсона была установлена, хвостатое тело также было связано с сопутствующими когнитивными нарушениями Паркинсона. В одном обзоре сравниваются показатели пациентов с болезнью Паркинсона и пациентов, строго страдающих поражением лобных долей Тест лондонского Тауэра. Различия в производительности между двумя типами пациентов (в тесте, который, вкратце, требует от испытуемых выбора подходящих промежуточных целей, имея в виду более крупную цель), устанавливает связь между хвостатым и целенаправленным действием. Однако исследования не являются окончательными. Хотя хвостатое тело связано с исполнительной функцией (см. «Действие, направленное на достижение цели»), остается «совершенно неясным, отражают ли исполнительные дефициты [пациентов с болезнью Паркинсона] преимущественно их корковое или подкорковое повреждение».[14]
болезнь Хантингтона
В болезнь Хантингтона генетическая мутация происходит в HTT ген, кодирующий белок Htt. Белок Htt взаимодействует с более чем 100 другими белками и, по-видимому, выполняет множество биологических функций.[32] Поведение этого мутированного белка до конца не изучено, но он токсичен для определенных типов клеток, особенно в головном мозге. Раннее повреждение наиболее очевидно в полосатое тело, но по мере прогрессирования заболевания другие области мозга также более заметно поражаются. Ранние симптомы связаны с функциями полосатого тела и его корковых связей, а именно с контролем над движением, настроением и высшими когнитивными функциями.[33]
Синдром дефицита внимания и гиперактивности
Исследование 2002 года показывает взаимосвязь между асимметрией хвостатого ядра и симптомами, связанными с СДВГ. Авторы использовали МРТ-изображения для сравнения относительных объемов хвостатых ядер (поскольку хвостатое ядро является двусторонней структурой) и установили связь между любыми асимметричными проявлениями и симптомами СДВГ: «Степень асимметрии хвостатого ядра в значительной степени предсказывала совокупные оценки тяжести невнимательного поведения. . " Эта корреляция согласуется с предыдущими ассоциациями хвостатого тела с функцией внимания.[34] Более недавнее исследование 2018 года воспроизвело эти результаты и продемонстрировало, что асимметрия хвостатого тела, связанная с СДВГ, была более выражена в дорсальных медиальных областях хвостатого тела.[35]
Шизофрения
Объем белого вещества в хвостатом ядре был связан с пациентами с диагнозом: Шизофрения. В исследовании 2004 года используется магнитно-резонансная томография для сравнения относительного объема белого вещества в хвостатом теле у пациентов с шизофренией. У пациентов, страдающих этим заболеванием, «абсолютный и относительный объем белого вещества в хвостатом ядре меньше, чем у здоровых людей».[36]
Биполярный тип I
Исследование 2014 года показало, что пациенты с биполярным расстройством I типа имеют относительно более высокий объем серого и белого вещества в хвостатом ядре и других областях, связанных с обработкой вознаграждения и принятием решений, по сравнению с контрольной группой и субъектами с биполярным расстройством II типа. В целом количество серого и белого вещества у пациентов с биполярным расстройством было ниже, чем у контрольной группы.[37][38]
Обсессивно-компульсивное расстройство
Было высказано предположение, что хвостатое ядро может быть дисфункциональным у людей с обсессивно-компульсивное расстройство (ОКР), в том смысле, что он может быть не в состоянии должным образом регулировать передачу информации о тревожных событиях или идеях между таламус и орбитофронтальная кора.
А нейровизуализация учиться с позитронно-эмиссионная томография обнаружили, что правое хвостатое ядро имело наибольшее изменение метаболизма глюкозы после лечения пациентов пароксетин.[39] Недавний Мета-анализ SDM из морфометрия на основе вокселей исследования, сравнивающие людей с ОКР и здоровых людей, показали, что у людей с ОКР увеличивается серое вещество объемы в двустороннем линзовидные ядра, распространяясь на хвостатые ядра, в то время как объем серого вещества уменьшался в двустороннем дорсальном медиальный лобный /передняя поясная извилина извилины.[40][41] Эти данные контрастируют с результатами у людей с другими тревожными расстройствами, у которых наблюдается снижение (а не увеличение) серое вещество объемы в двустороннем линзовидный / хвостатых ядер, а также уменьшил объем серого вещества в двусторонних дорсальных медиальный лобный /передняя поясная извилина извилины.[41]
Дополнительные изображения

Два вида модели полосатое тело (в правом полушарии): А, боковой аспект; B, медиальный аспект.
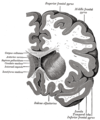
Венечный разрез по переднему рогу боковых желудочков.

Венечный отдел головного мозга через переднюю комиссуру.

Поверхностное рассечение ствола мозга. Боковой вид. Над зрительным нервом можно увидеть хвостатое ядро.

Рассечение ствола мозга. Боковой вид.

Глубокое рассечение ствола мозга. Боковой вид.

Глубокое рассечение ствола мозга. Боковой вид.
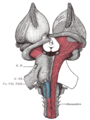
Поверхностное рассечение ствола мозга. Вид снизу.

Рассечение ствола мозга. Вид сверху.

Центральная часть, а также передний и задний рог боковых желудочков обнажены сверху.

Хвостатое ядро

Хвостатое ядро

Желудочки головного мозга и базальные ганглии. Улучшенный вид, горизонтальный разрез, глубокое рассечение

Желудочки головного мозга и базальные ганглии. Крупный план предыдущего изображения

Хвостатые ядра вместе с другими подкорковыми структурами в стеклянном мозге














Рекомендации
- ^ а б Ягер Л.М., Гарсия А.Ф., Вунш А.М., Фергюсон С.М. (август 2015 г.). «Все аспекты полосатого тела: роль в наркомании». Неврология. 301: 529–541. Дои:10.1016 / j.neuroscience.2015.06.033. ЧВК 4523218. PMID 26116518.
- ^ Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). Sydor A, Brown RY (ред.). Молекулярная нейрофармакология: основа клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. С. 147–148. ISBN 9780071481274.
- ^ Медведь, Марк Ф. (2016). Неврология: исследование мозга. Коннорс, Барри В. ,, Парадизо, Майкл А. (Четвертое изд.). Филадельфия. п. 502. ISBN 9780781778176. OCLC 897825779.
- ^ Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). Sydor A, Brown RY (ред.). Молекулярная нейрофармакология: основа клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. п. 326. ISBN 9780071481274.
Доказательства того, что хвостатое ядро и скорлупа влияют на обучение ответной реакции, получены из исследований повреждений на грызунах и приматах, а также из исследований нейровизуализации на людях и из исследований болезней человека. При болезни Паркинсона дофаминергическая иннервация хвостатого тела и скорлупы серьезно нарушается из-за гибели дофаминовых нейронов в компактной части черной субстанции (Глава 17). Пациенты с болезнью Паркинсона имеют нормальную декларативную память (если у них нет сопутствующей деменции, которая может возникать при болезни с тельцами Леви). Однако у них наблюдаются заметные нарушения обучения ответной реакции. Пациенты с болезнью Паркинсона или другими нарушениями базальных ганглиев, такими как болезнь Хантингтона (при которой сами хвостатые нейроны повреждаются), испытывают дефицит в других процедурных задачах обучения, таких как освоение новых двигательных программ.
- ^ Андерсон Б.А., Кувабара Х., Вонг Д.Ф., Робертс Дж., Рахмим А., Брашич Дж. Р., Кортни С.М. (август 2017 г.). «Связь сигналов дофаминергического вознаграждения с развитием предвзятости внимания: исследование позитронно-эмиссионной томографии». NeuroImage. 157: 27–33. Дои:10.1016 / j.neuroimage.2017.05.062. ЧВК 5600829. PMID 28572059.
- ^ Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). Sydor A, Brown RY (ред.). Молекулярная нейрофармакология: основа клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. п. 321. ISBN 9780071481274.
Функциональная нейровизуализация у людей демонстрирует активацию префронтальной коры и хвостатого ядра (части полосатого тела) при выполнении задач, требующих тормозящего контроля над поведением.
- ^ Йетериан Э. Х., Пандья Д. Н. (февраль 1995 г.). «Кортикостриатальные связи экстрастриарных зон зрения у макак-резусов». Журнал сравнительной неврологии. 352 (3): 436–57. Дои:10.1002 / cne.903520309. PMID 7706560.
- ^ Д'Суза, Донна. "Территории сосудов головного мозга - Справочная статья по радиологии - Radiopaedia.org". radiopaedia.org.
- ^ Макдугал, Дэвид. "Черная субстанция". Архивировано из оригинал 30 сентября 2013 г.. Получено 5 декабря 2013.
- ^ Постл Б.Р., Д'Эспозито М. (июль 1999 г.). «Диссоциация активности хвостатого ядра человека в пространственной и непространственной рабочей памяти: исследование фМРТ, связанное с событием». Исследование мозга. Когнитивные исследования мозга. 8 (2): 107–15. Дои:10.1016 / s0926-6410 (99) 00010-5. PMID 10407200.
- ^ Postle BR, D'Esposito M (июнь 2003 г.). «Пространственная активность рабочей памяти хвостатого ядра чувствительна к системе отсчета». Когнитивная, аффективная и поведенческая неврология. 3 (2): 133–44. Дои:10.3758 / cabn.3.2.133. PMID 12943328.
- ^ а б Вильябланка-младший (2010). «Почему у нас хвостатое ядро?». Acta Neurobiologiae Experimentalis. 70 (1): 95–105. PMID 20407491.
- ^ Белый НМ (апрель 2009 г.). «Некоторые основные моменты исследований эффектов поражений хвостатого ядра за последние 200 лет». Поведенческие исследования мозга. 199 (1): 3–23. Дои:10.1016 / j.bbr.2008.12.003. PMID 19111791.
- ^ а б c d е Grahn JA, Parkinson JA, Owen AM (апрель 2009 г.). «Роль базальных ганглиев в обучении и памяти: нейропсихологические исследования». Поведенческие исследования мозга. 199 (1): 53–60. Дои:10.1016 / j.bbr.2008.11.020. PMID 19059285.
- ^ Эллиотт Р., Ньюман Дж. Л., Лондж О. А., Дикин Дж. Ф. (январь 2003 г.). «Паттерны дифференциальной реакции в полосатом теле и орбитофронтальной коре головного мозга на финансовое вознаграждение у людей: исследование параметрической функциональной магнитно-резонансной томографии». Журнал неврологии. 23 (1): 303–7. Дои:10.1523 / JNEUROSCI.23-01-00303.2003. ЧВК 6742125. PMID 12514228.
- ^ а б Ханнан К.Л., Вуд С.Дж., Юнг А.Р., Велакулис Д., Филлипс Л.Дж., Соулсби Б., Бергер Дж., МакГорри П.Д., Пантелис С. (июнь 2010 г.). «Объем хвостатого ядра у лиц с очень высоким риском психоза: исследование поперечной магнитно-резонансной томографии». Психиатрические исследования. 182 (3): 223–30. Дои:10.1016 / j.pscychresns.2010.02.006. PMID 20488675.
- ^ Левитт Дж. Дж., Маккарли Р. У., Дики К. С., Фоглмайер М. М., Низникевич М. А., Зейдман Л. Дж., Хираясу Ю., Чишевский А. А., Кикинис Р., Джолес Ф. А., Шентон М.Э. (июль 2002 г.). «МРТ-исследование объема хвостатого ядра и его когнитивных коррелятов у пациентов с шизотипическим расстройством личности, ранее не получавших нейролептики». Американский журнал психиатрии. 159 (7): 1190–7. Дои:10.1176 / appi.ajp.159.7.1190. ЧВК 2826363. PMID 12091198.
- ^ Макгоу Дж. Л. (2004). «Миндалевидное тело модулирует консолидацию воспоминаний об эмоционально возбуждающих переживаниях». Ежегодный обзор нейробиологии. 27: 1–28. Дои:10.1146 / annurev.neuro.27.070203.144157. PMID 15217324.
- ^ Seger CA, Cincotta CM (март 2005 г.). «Роль хвостатого ядра в обучении классификации человека». Журнал неврологии. 11. 25 (11): 2941–51. Дои:10.1523 / jneurosci.3401-04.2005. ЧВК 6725143. PMID 15772354.
- ^ Гогичадзе М, Ониани МТ, Немсадзе М, Ониани Н (2009). «Нарушения сна и нарушения обработки памяти, связанные с поражением хвостатого ядра». Паркинсонизм и связанные с ним расстройства. 15: S167 – S168. Дои:10.1016 / S1353-8020 (09) 70639-X.
- ^ Виллабланка-младший (сентябрь 2004 г.). «Противопоставление функциональной роли переднего мозга и ствола мозга в управлении системой сна-бодрствования». Журнал исследований сна. 13 (3): 179–208. Дои:10.1111 / j.1365-2869.2004.00412.x. PMID 15339255.
- ^ Кауфманн С., Верле Р., Веттер Т.С., Хольсбоер Ф., Ауэр Д.П., Поллмахер Т., Циш М. (март 2006 г.). «Активация мозга и функциональная связь гипоталамуса во время сна человека с небыстрым движением глаз: исследование ЭЭГ / фМРТ». Мозг. 129 (Pt 3): 655–67. Дои:10.1093 / мозг / awh686. PMID 16339798.
- ^ Кумар Р., Ахдут Р., Мейси П.М., Ву М.А., Аведиссиан К., Томпсон П.М., Харпер Р.М. (ноябрь 2009 г.). «Снижение объемов хвостатого ядра у пациентов с врожденным синдромом центральной гиповентиляции». Неврология. 163 (4): 1373–9. Дои:10.1016 / j.neuroscience.2009.07.038. ЧВК 2761724. PMID 19632307.
- ^ Ишизу Т., Зеки С. (май 2011 г.). Ордер EJ (ред.). «К теории красоты, основанной на мозге». PLOS ONE. 6 (7): e21852. Дои:10.1371 / journal.pone.0021852. ЧВК 3130765. PMID 21755004.
- ^ Арон А., Фишер Х, Машек Д. Д., Стронг Дж., Ли Х, Браун Л. Л. (июль 2005 г.). «Системы вознаграждения, мотивации и эмоций, связанные с интенсивной романтической любовью на ранней стадии». Журнал нейрофизиологии. 94 (1): 327–37. Дои:10.1152 / ян.00838.2004. PMID 15928068.
- ^ Кринион Дж., Тернер Р., Гроган А., Ханакава Т., Ноппени У., Девлин Дж. Т., Асо Т., Ураяма С., Фукуяма Х, Стоктон К., Усуи К., Грин Д. У., Прайс Си Джей (июнь 2006 г.). «Языковой контроль в двуязычном мозгу». Наука. 312 (5779): 1537–40. Дои:10.1126 / science.1127761. PMID 16763154. S2CID 10445511.
- ^ «Как двуязычный мозг переключается между языками». newscientist.com.
- ^ Newberg AB, Wintering NA, Morgan D, Waldman MR (ноябрь 2006 г.). «Измерение регионального церебрального кровотока во время глоссолалии: предварительное ОФЭКТ-исследование». Психиатрические исследования. 148 (1): 67–71. Дои:10.1016 / j.pscychresns.2006.07.001. PMID 17046214.
- ^ Брайтенберг В. (1984) Транспортные средства. Эксперименты по синтетической психологии.
- ^ Джиджи С., Смита К.А., Гупта А.К., Пиллаи В.П., Джаясри Р.С. (сентябрь 2013 г.). «Сегментация и объемный анализ хвостатого ядра при болезни Альцгеймера». Европейский журнал радиологии. 82 (9): 1525–30. Дои:10.1016 / j.ejrad.2013.03.012. PMID 23664648.
- ^ Колб, Брайан; Иэн К. Уишоу (2001). Введение в мозг и поведение (4-е изд.). Нью-Йорк: Worth Publishers. п.590. ISBN 978-1429242288.
- ^ Goehler H, Lalowski M, Stelzl U, Waelter S, Stroedicke M, Worm U, Droege A, Lindenberg KS, Knoblich M, Haenig C, Herbst M, Suopanki J, Scherzinger E, Abraham C, Bauer B, Hasenbank R, Fritzsche , Ludewig AH, Büssow K, Buessow K, Coleman SH, Gutekunst CA, Landwehrmeyer BG, Lehrach H, Wanker EE (сентябрь 2004 г.). «Сеть взаимодействия белков связывает GIT1, усилитель агрегации хантингтина, с болезнью Хантингтона». Молекулярная клетка. 15 (6): 853–65. Дои:10.1016 / j.molcel.2004.09.016. PMID 15383276.
- ^ Уокер Ф.О. (январь 2007 г.). "Болезнь Хантингтона". Ланцет. 369 (9557): 218–28. Дои:10.1016 / S0140-6736 (07) 60111-1. PMID 17240289.
- ^ Шримшер Г.В., Биллингсли Р.Л., Джексон Е.Ф., Мур Б.Д. (декабрь 2002 г.). «Асимметрия объема хвостатого ядра предсказывает симптоматику синдрома дефицита внимания и гиперактивности (СДВГ) у детей». Журнал детской неврологии. 17 (12): 877–84. Дои:10.1177/08830738020170122001. PMID 12593459.
- ^ Дуглас П.К., Гутман Б., Андерсон А., Лариос С., Лоуренс К. Э., Нарр К., Сенгупта Б., Курей Г., Дуглас Д. Б., Томпсон П. М., Макгоф Дж. Дж., Букхаймер С. Ю. (февраль 2018 г.). «Различия в асимметрии полушарного мозга у молодых людей с синдромом дефицита внимания / гиперактивности». NeuroImage: Клинический. 18: 744–52. Дои:10.1016 / j.nicl.2018.02.020. ЧВК 5988460. PMID 29876263.
- ^ Такасе К., Тамагаки С., Окугава Г., Нобухара К., Минами Т., Сугимото Т., Савада С., Киношита Т. (2004). «Уменьшение объема белого вещества хвостатого ядра у больных шизофренией». Нейропсихобиология. 50 (4): 296–300. Дои:10.1159/000080956. PMID 15539860. ProQuest 293981781.
- ^ Маллер, Джером Дж .; Тавентиран, Прасантан; Томсон, Ричард Х .; Маккуин, Сьюзен; Фитцджеральд, Пол Б. (2014). «Объемные, корковые изменения толщины и целостности белого вещества при биполярном расстройстве I и II типа». Журнал аффективных расстройств. 169: 118–127. Дои:10.1016 / j.jad.2014.08.016. PMID 25189991.
- ^ «Путешествие в биполярный мозг». psmag.com.
- ^ Hansen ES, Hasselbalch S, Law I, Bolwig TG (март 2002 г.). «Хвостатое ядро при обсессивно-компульсивном расстройстве. Снижение метаболизма после лечения пароксетином: исследование ПЭТ». Международный журнал нейропсихофармакологии. 5 (1): 1–10. Дои:10.1017 / S1461145701002681. PMID 12057027.
- ^ Radua J, Mataix-Cols D (ноябрь 2009 г.). «Воксельный метаанализ изменений серого вещества при обсессивно-компульсивном расстройстве». Британский журнал психиатрии. 195 (5): 393–402. Дои:10.1192 / bjp.bp.108.055046. PMID 19880927.
- ^ а б Радуа Дж., Ван ден Хеувель О.А., Сургуладзе С., Матэ-Колс Д. (июль 2010 г.). «Метааналитическое сравнение исследований морфометрии на основе вокселей при обсессивно-компульсивном расстройстве и других тревожных расстройствах». Архив общей психиатрии. 67 (7): 701–11. Дои:10.1001 / archgenpsychiatry.2010.70. PMID 20603451.
внешняя ссылка
- Изображения окрашенных срезов головного мозга, включающие «хвостатое ядро» на Проект BrainMaps
- Схема на uni-tuebingen.de
- Поиск NIF - Хвостатое ядро через Информационная структура по неврологии
{kind=link}
| Авторитетный контроль |
|---|