Dyrosauridae - Dyrosauridae
| Dyrosauridae | |
|---|---|
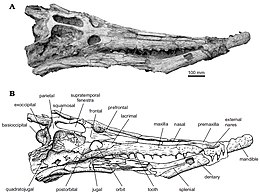 | |
| Череп дирозаврида Arambourgisuchus khouribgaensis | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Рептилии |
| Подотряд: | †Тетисухия |
| Семья: | †Dyrosauridae де Стефано, 1903 г. |
| Роды | |
Dyrosauridae это семья вымерших неозухий крокодиловидные который жил из Поздний мел (Маастрихтский ) к эоцен. Окаменелости дирозаврид распространены по всему миру, они были найдены в Африке, Азии, Европе, Северной Америке и Южной Америке. В настоящее время известно более десятка видов, сильно различающихся по размеру и форме черепа. Все они были предположительно водными, причем виды обитали как в пресноводной, так и в морской среде. Обитающие в океане дирозавриды были одними из немногих морских рептилий, переживших Меловое – палеогеновое вымирание.
Дирозавриды представляли собой группу в основном морских, длинночелюстных, крокодилоподобных четвероногих до 6 метров (20 футов) в длину.[1] Основываясь на данных о костной ткани, было высказано предположение, что они медленно растут.[2] прибрежные морские животные с сомкнутыми челюстями,[2] умеет плавать, а также ходить по суше. Наружные ноздри на заднем конце рыла и внутренний нос в крыловидном коже указывали на привычку к охоте во время плавания с макушкой над водой, что позволяло ему дышать во время преследования добычи.[1]
Обзор

Известно, что дирозавриды имеют очень характерную форму черепа с длинной и тонкой мордой, которая составляет примерно 68% от общей длины черепа. Самая передняя часть черепа и морды - это внешний нарис за которыми следуют два предчелюстные кости кости, пока они не достигнут двух максиллы кости, разделенные одной носовая кость.[1]
Типичным у дирозавридов является единственный носовой элемент с характерным набором небольших ямок и постоянной шириной, пока он не расширится, чтобы контактировать с слезный кости, затем сужаются на короткое расстояние, пока не встретятся с границей лобных и префронтальных костей.[1]

У дирозавров есть предчелюстная кость с неглубокими ямками, которые простираются кзади от третьих альвеол верхней челюсти. Есть две предчелюстные кости, которые более узкие по сравнению с верхнечелюстными костями и проходят в двух длинных. верхняя челюсть кости, которые разделены единственной носовой костью. Последняя предчелюстная кость и первая верхняя челюсть широко разделены четвертым зубным зубом. Альвеолы широко расставлены спереди и расстояние между ними уменьшается. сзади из пятнадцатой альвеолы при постоянном диаметре.[1]
Верхняя челюсть длинная (примерно в два с половиной раза длиннее скуловой ) и составляет большую часть бокового края черепа.[3] Согласно Жуву и Барбозе и, возможно, в зависимости от возраста животного, каждая верхняя челюсть имеет 13-19 зубов.[3]
Важная особенность дирозавридов зубной ряд глубокие окклюзионный ямки, особенно присутствующие в задней части верхних челюстей, становятся менее выраженными спереди. Ямки указывают на взаимосвязанную закрытую челюсть для дирозавридов, потому что ямки дают возможность чередоваться верхней и нижней челюстям. Хотя они все еще присутствуют у dyrosaurus phosphaticus, эта особенность глубоких окклюзионных ямок становится менее развитой и широкой.[3] Зубы Dyrosaurus гомодонты, конические, длинные и тонкие, с асимметричными губными и язычными поверхностями. Задние зубы короче и более сжатые, чем передний зуб, что указывает на уменьшение размера зуба от переднего к заднему.

У дирозавров длина от морды до черепа составляет около 68%, и у рода Рабдогнат и Атлантозух, Дирозавр и Арамбургизухус имеют самые большие пропорции морды среди всех дирозаврид. Длина рыла ранее использовалась для установления дирозавридных отношений, при этом считалось, что удлинение морды является «более развитым» признаком. Это не соответствовало заключению Жува, который заключался в том, что самая длинная морда на самом деле является примитивным состоянием, поэтому более короткая или длинная морда появляется независимо по крайней мере четыре раза в эволюции дирозаврид.[4]
Палеобиогеография
Когда-то дирозавриды считались африканской группой, но открытия, сделанные начиная с 2000-х годов, указывают на то, что они населяли большинство континентов.[5] Фактически, базальные формы предполагают, что их колыбелью могла быть Северная Америка.
Общий
| Род | Положение дел | Возраст | Место расположения | Описание | Изображений |
|---|---|---|---|---|---|
| Ахеронтисух | Действительный | Палеоцен | Пресноводный дирозаврид с большим телом и длинной мордой из Формация Серрехон | ||
| Эгиалозух | Действительный | Кампанский | Морской дирозаврид из Кристианстад Бассейн | ||
| Антракозух | Действительный | Палеоцен | Пресноводный дирозаврид с короткой мордой из Формация Серрехон | ||
| Арамбургизухус | Действительный | Палеоцен | Морской дирозаврид с длинной мордой | ||
| Атлантозух | Действительный | Палеоцен | Морской дирозаврид с длинной мордой и самой длинной мордой по сравнению с размером тела среди всех дирозаврид. | ||
| Cerrejonisuchus | Действительный | Палеоцен | Пресноводный дирозаврид с мелким телом и короткой мордой из Формация Серрехон | ||
| Chenanisuchus | Действительный | Маастрихтский -Палеоцен | Род охватывает K-Pg граница |  | |
| Конгозавр | Действительный | Палеоцен | |||
| Дирозавр | Действительный | эоцен | Крупный, длинноносый морской дирозаврид |  | |
| Fortignathus | Действительный | Альбианский –Сеноманский | Дирозаврид из Эчкарская формация. | ||
| Guarinisuchus | Действительный | Палеоцен | Морской дирозаврид |  | |
| Гипозавр | Действительный | Маастрихтский -Палеоцен | Было названо пять видов, большинство из всех родов дирозаврид; род охватывает K-Pg граница | ||
| Фосфатозавр | Действительный | эоцен | Крупный, длинноносый морской дирозаврид с тупыми зубами и кончиком морды в форме ложки. | ||
| Рабдогнат | Действительный | Маастрихтский -Палеоген | Крупный, длинноносый морской дирозаврид; род охватывает K-Pg граница | ||
| Сабинозух | Действительный | Маастрихтский | Дирозаврид из Формация Эскондидо. | ||
| Сокотозавр | Младший синоним | Младший синоним Гипозавр | |||
| Sokotosuchus | Действительный | Маастрихтский | Морской дирозаврид с длинной мордой | ||
| Тилемсисухус | Действительный | эоцен |
Филогения
Jouve и другие. (2005) диагностировали Dyrosauridae как клады на основе следующих семи синапоморфии или общие персонажи:
- Заднемедиальное крыло ретроартикулярного отростка, расположенное дорсально снизу на ретроартикулярном отростке
- Затылочные бугорки небольшие
- Exoccipital в значительной степени участвует в затылочный мыщелок
- Надвисочное отверстие переднезаднее сильно удлиненное
- Симфиз примерно такой же ширины, как высота
- Квадратично-скуловая кость в значительной степени участвует в черепном мыщелке для сочленения с челюстью.
- 4 предчелюстных зуба
Ниже приводится кладограмма после Жува и другие. (2005), показывающие филогенетические отношения Dyrosauridae и других близкородственных неозухийцы:
| Neosuchia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Составная кладограмма Dyrosauridae (от Jouve и другие. 2008 и Барбоза и другие. 2008):
| Dyrosauridae |
Dyrosauridae incertae sedis:Тилемсисухус | |||||||||||||||||||||||||||||||||||||||||||||||||||
Кладограмма по Гастингсу и другие. (2011) с указанием географического расположения таксонов:[6]
| Neosuchia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Анализ показывает, что ближайшие родственники дирозаврид Саркозух и Терминонарис.
Палеобиология
Рост

Доказательства полуводной жизни дирозаврид получены в результате тщательного анализа костной структуры. У водных четвероногих может быть два типа структурной организации кости: остеопоротический или же пахиостотический. Остеопоротическая кость является губчатой и пористой, тогда как при пахиостозе увеличивается скелетная масса. Губчатая / пористая кость, такая как остеопоротическая, связана с более быстрым плаванием и лучшей маневренностью в воде из-за сокращения костной ткани, у многих современных китообразных и морских черепах есть остеопоротическая кость, что позволяет им хорошо плавать. Пахиостотическая кость - это общее / местное увеличение массы скелета, которое может быть вызвано остеосклерозом (внутреннее уплотнение кости), пахиостозом (гиперплазией компактной коры) или пахиосклерозом (комбинация этих двух). Исследование кости дирозавра, проведенное Рафаэлем Сезаром Лимой Педросо де Андраде и Джулианой Мансо Саяо, показало, что у этой семьи была остеопоротическая костная ткань, указывающая на быстрое плавание. экология а также некоторые остеосклероз который входит в состав костной ткани пахиостоза. Остеопороз связан с полностью водным образом жизни, в то время как пахиостоз не является полностью водным, но связан с экологией быстрого плавания. Следовательно, дирозавры - полуводные быстро плавающие, на что указывает их костная структура. Другим свидетельством прибрежного полуводного образа жизни являются окаменелости, часто в переходных морских отложениях.[2] -использование осевого частотного плавания (которое используется в основном современными крокодилами) с большей волнообразный движение и частота хвоста благодаря высокоразвитому мускулатура позволяя более мощный толчок вперед.[2]
У дирозавридов есть структура ткани, которая указывает на медленнорастущее животное, что было определено путем тщательного анализа правильного бедренная кость и влево большеберцовая кость. В левой большеберцовой кости кора головного мозга имела пластинчатую зональную кость с пятью линиями задержанного роста (LAG), которые были разнесены на 300 мм друг от друга, а также четкую сосудистую сеть первичных остеонов, плотность которых уменьшалась по направлению к мембране (периостально). На правом бедре были двойные LAG и позже EFS, а также вторичные остеоны, возникающие в глубокой коре и спонгиозе. Такой паттерн роста тканей является общей характеристикой медленно растущих животных.[2]
Среда обитания
Дирозавриды встречаются в переходных морских отложениях от позднего мела до нижнего эоцена.[2] Это семейство известно в основном из маастрихтских отложений в Нью-Джерси и от позднего мела до раннего палеогена в породах моря Тетис в Северной и Западной Африке.[2] Окаменелости также были обнаружены в палеоценовых и эоценовых слоях Пакистана, а также в Южной Америке, Бразилии, Индии, Южной Азии и прибрежных районах. Обычно дирозавриды извлекаются из прибрежных и устьевых отложений в Северной Африке и на Ближнем Востоке, что подтверждает их существование как полуводных животных.[7]
Наибольшее таксономическое разнообразие Dyrosauridae было в период раннего Палеоген но похоже, что клады смог получить большее и более широкое географическое распространение в течение позднего мелового периода. Самые ранние записи дирозаврид находятся либо в Африке, либо рядом с ней, с фрагментарными находками из Сеноманский Судана и Португалии и некоторых другихМаастрихтский, Позднемеловые открытия в Египте. Позже, к маастрихту Северной Америки, летопись дирозаврид стала более полной, установив широкое распространение, которое, по-видимому, сохранялось в палеоцене и эоцене.[8]
Дирозавриды также были обнаружены в неморских отложениях. В северной Судан, дирозавриды известны из речной отложения, что указывает на то, что они жили в условиях реки.[9] Кости неопределенных дирозавридов были найдены во внутренних отложениях в Пакистан также. Некоторые дирозавриды, например, из Формация Умм Химар в Саудовская Аравия, жилой эстуарий окружающая среда недалеко от побережья. Недавно названные дирозавриды Cerrejonisuchus и Ахеронтисух были восстановлены из Формация Серрехон на северо-западе Колумбия, который, как считается, представляет собой переходную морско-пресноводную среду, окруженную тропическими лесами, более удаленными от суши, чем устьевой среды формации Умм-Химар.[10] Cerrejonisuchus и Ахеронтисух жили в неотропической обстановке в то время, когда глобальные температуры были намного теплее чем они есть сегодня.[11][12]
Размножение
В 1978 году было высказано предположение, что взрослые особи дирозавриды жили в океане, но размножались во внутренних пресноводных средах. Останки мелкотелых дирозаврид из Пакистана интерпретировались как молодые. Их присутствие во внутренних отложениях рассматривалось как свидетельство того, что дирозавриды вылупились далеко от океана.[13] Из нижнего эоцена бассейна Улад-Абдун очень мало молодых дирозаврид, но много взрослых особей аналогичного размера. Это подтвердило предположение, что молодые особи жили в пресноводных средах, а взрослые особи - в морских.[14] Однако недавно крупнотелые и полностью зрелые дирозавриды формации Серрехон показали, что некоторые дирозавриды прожили всю свою жизнь во внутренних районах, никогда не возвращаясь на побережье.[6]
Рекомендации
- ^ а б c d е Жув, Стефани. Новое описание черепа Dyrosaurus phosphaticus (Thomas, 1893) (Mesoeucrocodylia: Dyrosauridae) из нижнего эоцена Северной Африки. 2005. Получено 2 марта 2015 г. из http://cjes.nrc.ca В архиве 2002-06-04 на Wayback Machine.
- ^ а б c d е ж грамм Андраде RCLPd, Saya ̃o JM (2014) Палеогистология и выводы об образе жизни дирозавридов (Archosauria: Crocodylomorpha) из бассейна Параиба (северо-восток Бразилии). PLoS ONE 9 (7): e102189. DOI: 10.1371 / journal.pone.0102189
- ^ а б c Антонио Барбоза, Хосе; Вильгельм Армин Келлнер, Александр. Сомалиа Сейлз Виана, Мария. Новые крокодиломорфы дирозаврид и свидетельства круговорота фауны на переходе К – П в Бразилии. 2008. Труды Королевского общества. Загружено с http://rspb.royalsocietypublishing.org/
- ^ Жув, Стефани; Буйя, Баади; Амагзаз, Мбарек. ДИРОЗАВРИД С ДЛИННЫМ РЫКОМ (CROCODYLIFORMES, MESOEUCROCODYLIA) ИЗ ПАЛЕОЦЕНА МАРОККО: ФИЛОГЕНЕТИЧЕСКИЕ И ПАЛАЕОБИОГЕОГРАФИЧЕСКИЕ ПОСЛЕДСТВИЯ. 2008. Палеонтология, т. 51, часть 2. С. 281-294.
- ^ Jouve и другие. (2008)
- ^ а б Гастингс А.К., Блох Дж. И Харамилло К.А. (2011). «Новый лонгирострин дирозаврид (Crocodylomorpha, Mesoeucrocodylia) из палеоцена на северо-востоке Колумбии: биогеографические и поведенческие последствия для дирозаврид нового мира» (PDF). Палеонтология. 54 (5): 1095–116. Дои:10.1111 / j.1475-4983.2011.01092.x. Архивировано из оригинал (PDF) 31 марта 2012 г.. Получено 14 сен 2011.CS1 maint: несколько имен: список авторов (связь)
- ^ Мэтью С. Ламанна, Джошуа Б. Смит, Юсри С. Аттиа и Питер Додсон Источник: Journal of Vertebrate Paleontology, Vol. 24, No. 3 (10 сентября 2004 г.), стр. 764-768. Издано: Taylor & Francis, Ltd. от имени Общества палеонтологии позвоночных. Стабильный URL: https://www.jstor.org/stable/4524765 Дата обращения: 03.04.2015 17:11 UTC.
- ^ Ашу Хосла, Джозеф Дж. В. Сертич, Гунтупалли В. Р. Прасад и Омкар Верма Источник: Journal of Vertebrate Paleontology, Vol. 29, No. 4 (12 декабря 2009 г.), pp. 1321-1326 Издатель: Taylor & Francis, Ltd. от имени Общества палеонтологии позвоночных. Стабильный URL: https://www.jstor.org/stable/20627142. Дата обращения: 03-03-2015 07:01 UTC
- ^ Buffetaut, E .; Буссерт, Р .; Бринкманн, В. (1990). «Новая неморская фауна позвоночных в верхнемеловом периоде северного Судана». Berliner Geowissenschaftlische Abhandlungen. 120: 183–202.
- ^ Hastings, A.K; Bloch, J. I .; Cadena, E. A .; Харамилло, К. А. (2010). «Новый маленький короткомордый дирозаврид (Crocodylomorpha, Mesoeucrocodylia) из палеоцена на северо-востоке Колумбии». Журнал палеонтологии позвоночных. 30 (1): 139–162. Дои:10.1080/02724630903409204. S2CID 84705605.
- ^ Head, J. J .; Bloch, J. I .; Hastings, A.K .; Borque, J. R .; Cadena, E. A .; Herrera, F.A .; Polly, P.D .; Харамилло, К. А. (2009). «Гигантская змея из неотропов палеоцена показывает более горячие прошлые экваториальные температуры». Природа. 457 (7230): 715–717. Bibcode:2009Натура.457..715H. Дои:10.1038 / природа07671. PMID 19194448. S2CID 4381423.
- ^ Канапо, Б. (2 февраля 2010 г.). "Исследователи UF: Древний крокодил, относительный вероятный источник пищи для Титанобоа". Новости Университета Флориды. Архивировано из оригинал 8 июня 2010 г.. Получено 3 февраля, 2010.
- ^ Баффето, Э. (1978). «Останки крокодилов из эоцена Пакистана». Neues Jahrbuch für Geologie und Paläontologie, Абхандлунген. 156: 262–283.
- ^ Жув, Стефани; Бардет, Натали; Джалил, Нур-Эддин; Супербиола, Хабье Переда; Буйя, Баади и Амагзаз, Мбарек. 2008. Самый старый африканский крокодил: филогения, палеобиогеография и дифференциальная выживаемость морских рептилий через границу мелового и третичного периода. Журнал палеонтологии позвоночных 28 (2): 409–421, июнь 2008 г.
- Барбоза, Д.А., Келлнер, А.В.А. и Виана, M.S.S. (2008). Новый крокодиломорф дирозаврид и свидетельства круговорота фауны на переходе К – П в Бразилии. Труды Королевского общества B: биологические науки: Firstcite
- Баффето, Э. (1985). L'evolution des crocodiliens. Les animaux disparus-Pour la science, Париж 109.
- Jouve, S .; Bouya, B .; Амагзаз, М. (2008). «Длиннорылый дирозаврид (Crocodyliformes, Mesoeucrocodylia) из палеоцена Марокко: филогенетические и палеогеографические последствия». Палеонтология. 51 (2): 281–294. Дои:10.1111 / j.1475-4983.2007.00747.x.
- Jouve, S .; Iarochène, M .; Bouya, B .; Амагзаз, М. (2005). "Новый крокодилиформ дирозаврид из палеоцена Марокко и филогенетический анализ Dyrosauridae". Acta Palaeontologica Polonica. 50 (3): 581–594.
внешняя ссылка
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||

