I-CreI - I-CreI
| ДНК-эндонуклеаза I-CreI | |||||||
|---|---|---|---|---|---|---|---|
 | |||||||
| Идентификаторы | |||||||
| Организм | |||||||
| Символ | ? | ||||||
| UniProt | P05725 | ||||||
| |||||||
Я-Creя это самонаводящаяся эндонуклеаза чей ген был впервые обнаружен в хлоропласт геном Хламидомонада Reinhardtii, вид одноклеточных зеленые водоросли.[1] Он назван в честь того факта, что: он находится в яntron; он был изолирован от Cламидомонада повторноInhardtii; это был первый (я) такой ген выделен из C. reinhardtii. Его ген находится в группе I интрон в 23-е годы рибосомная РНК ген C. reinhardtii хлоропласт и I-CreЯ выражается только тогда, когда его мРНК сращен из первичная стенограмма гена 23S. Я-Creя фермент, который функционирует как гомодимер, распознает 22-нуклеотидную последовательность дуплексной ДНК и расщепляет одну фосфодиэфирную связь на каждой цепи в определенных положениях. Я-CreЯ являюсь членом Семья LAGLIDADG самонаводящихся эндонуклеаз, каждая из которых имеет консервативный аминокислотный мотив LAGLIDADG, который способствует их ассоциативным доменам и активным сайтам. Когда я-CreI-содержащий интрон встречает ген 23S, лишенный интрона, I-CreФермент I «обитает» на аллеле «минус интрон» 23S и влияет на вставку его родительского интрона в аллель без интрона. Интроны с таким поведением называются мобильные интроны. Потому что я-CreI обеспечивает собственное распространение, не принося никакой пользы хосту, это пример эгоистичная ДНК.
Открытие
Я-CreВпервые меня заметили как промежуточная последовательность в 23-е годы рРНК ген C. reinhardtii хлоропластный геном.[1] Ген 23S - это Ген РНК, что означает, что его транскрипт не транслируется в белок. Как РНК, он является частью большой субъединицы рибосома. Открытая рамка считывания, кодирующая белок из 163 аминокислот, была обнаружена в этом интроне 23S, что позволяет предположить, что белок может способствовать поведению хоминга мобильного интрона. Кроме того, предсказанный белок имел мотив LAGLIDADG, консервативную аминокислотную последовательность, которая присутствует в других белках, кодируемых в мобильных интронах группы I. Исследование 1991 года установило, что ORF кодирует эндонуклеазу ДНК, I-CreI, который выборочно разрезает сайт, соответствующий тому месту, где интрон сплайсирован из первичного транскрипта 23S.[2] Исследование также показало, что интрон способен проникать в аллели 23S, в которых его еще не было.[2]
Механизм распространения
Я-CreI эволюционировал, чтобы разрезать 22-нуклеотидную последовательность ДНК, которая встречается в аллели гена 23S рибосомальной РНК, лишенного I-CreI-содержащий интрон. Когда такой аллель "минус интрон" отсекается, пути восстановление двухниточного разрыва активируются в ячейке. Клетка использует в качестве шаблона для восстановления аллеля 23S, который дал ответственный I-CreI фермент, таким образом реплицируя I-CreI-содержащий интрон.[3] Получающийся в результате аллель "интрон-плюс" больше не содержит интактного сайта хоминга для I-CreI фермент и поэтому не расщепляется. Поскольку этот интрон обеспечивает собственную репликацию, не принося никакой пользы своему хозяину, I-CreЯ форма эгоистичная ДНК.
Структурные исследования и возможные применения

Потому что я-CreЯ эволюционировал, чтобы разрезать такую длинную последовательность ДНК, в отличие от эндонуклеазы рестрикции который обычно разрезает последовательности из четырех или шести нуклеотидов, он способен разрезать один сайт в очень большом геном. Ожидается, что последовательность из четырех или шести нуклеотидов будет встречаться много-много раз в геноме из миллионов или миллиардов нуклеотидов просто случайно, тогда как последовательность из 22 нуклеотидов может встречаться только один раз (109/46 против 109/422). Эта специфика I-CreЯ расщепление делает Я-CreЯ многообещающий инструмент для нацеливание на гены. Если человек заболел из-за дефектного аллель некоторых ген, было бы полезно иметь возможность заменить этот аллель на функциональный. Если бы можно было заставить я-CreЯ разрезал ДНК только на дефектном аллеле, одновременно предоставляя клетке нормальный аллель для использования в качестве матрицы восстановления, собственный гомологичная рекомбинация машины могут вставить желаемый аллель вместо дисфункционального. Специфика I-CreI также позволяет уменьшить вредные воздействия из-за двухниточные разрывы вне интересующего гена.
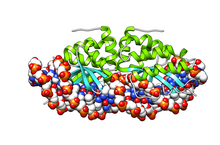
Чтобы использовать I-CreВ качестве инструмента в этой области необходимо заставить его распознавать и расщеплять последовательности ДНК, отличные от исходного сайта. An кишечная палочка генетическая система для изучения взаимосвязи между I-CreI структура и специфика ее сайта была создана в 1997 году.[5] В 1997 году была определена структура белка I-CreI,[6] а в 1998 году была решена его кристаллическая структура, связанная с его нативным сайтом наведения ДНК, что в значительной степени помогло исследованиям по изменению распознавания этого белка сайтом самонаведения.[4] С тех пор были созданы мутантные формы белка, которые проявляют измененную специфичность сайта хинга.[7][8][9] Генетическая система в Saccharomyces cerevisiae также был создан, давая дополнительные I-CreI-мутанты с измененной специфичностью сайта самонаведения.[10][11]
Я-CreI уже успешно использовался для индукции гомологичной рекомбинации в Drosophila melanogaster, чрезвычайно популярный эукариотический модельный организм.[12] Весьма вероятно, что успехи в молекулярно-биологических методах и создание библиотеки I-CreНовые эндонуклеазы, производные от I, в конечном итоге позволят воздействовать на многие гены этиологического значения.
Рекомендации
- ^ а б Rochaix, JD; Мальное, П (1978). «Анатомия рибосомной ДНК хлоропласта Chlamydomonas reinhardtii". Клетка. 15 (2): 661–670. Дои:10.1016 / 0092-8674 (78) 90034-х. PMID 719757.
- ^ а б Dürrenberger F, Rochaix JD (ноябрь 1991 г.). «Рибосомный интрон хлоропласта Chlamydomonas reinhardtii: самосплайсинг in vitro, активность эндонуклеаз ДНК и подвижность in vivo». Журнал EMBO. 10 (11): 3495–501. Дои:10.1002 / j.1460-2075.1991.tb04913.x. ЧВК 453078. PMID 1915304.
- ^ Дюрренбергер Ф., Томпсон А.Дж., Херрин Д.Л., Роша Д.Д. (сентябрь 1996 г.). «Двухцепочечная рекомбинация, индуцированная разрывом в хлоропластах Chlamydomonas reinhardtii». Исследования нуклеиновых кислот. 24 (17): 3323–31. Дои:10.1093 / nar / 24.17.3323. ЧВК 146090. PMID 8811085.
- ^ а б Юрица М.С., Моннат Р.Дж., Стоддард Б.Л. (октябрь 1998 г.). «Распознавание ДНК и расщепление эндонуклеазой LAGLIDADG самонаведения I-CreI». Молекулярная клетка. 2 (4): 469–76. Дои:10.1016 / с1097-2765 (00) 80146-х. PMID 9809068.
- ^ Селигман, Л. М.; Стивенс, КМ; Savage, JH; Моннат, Р.Дж. (1997). «Генетический анализ Chlamydomonas reinhardtii Я-CreI Мобильная система самонаведения интрон в кишечная палочка". Генетика. 147 (4): 1653–1664. ЧВК 1208338. PMID 9409828.
- ^ Хит П.Дж., Стивенс К.М., Моннат Р.Дж., Стоддард Б.Л. (июнь 1997 г.). «Структура I-Crel, эндонуклеазы, кодирующей интрон группы I». Структурная биология природы. 4 (6): 468–76. Дои:10.1038 / nsb0697-468. PMID 9187655.
- ^ Селигман Л.М., Чисхолм К.М., Шевалье Б.С., Чадси М.С., Эдвардс С.Т., Сэвидж Дж. Х., Вейле А.Л. (сентябрь 2002 г.). «Мутации, изменяющие специфичность расщепления самонаводящейся эндонуклеазы». Исследования нуклеиновых кислот. 30 (17): 3870–9. Дои:10.1093 / nar / gkf495. ЧВК 137417. PMID 12202772.
- ^ Суссман Д., Чадси М., Фаус С., Энгель А., Брутт А., Моннат Р., Стоддард Б. Л., Селигман Л. М. (сентябрь 2004 г.). «Выделение и характеристика специфичности новых эндонуклеаз самонаведения в положениях отдельных сайтов-мишеней». Журнал молекулярной биологии. 342 (1): 31–41. Дои:10.1016 / j.jmb.2004.07.031. PMID 15313605.
- ^ Розен Л.Е., Моррисон Х.А., Масри С., Браун М.Дж., Спрингстубб Б., Суссман Д., Стоддард Б.Л., Селигман Л.М. (2006). «Самонаводящиеся производные эндонуклеазы I-CreI с новой специфичностью ДНК-мишени». Исследования нуклеиновых кислот. 34 (17): 4791–800. Дои:10.1093 / нар / gkl645. ЧВК 1635285. PMID 16971456.
- ^ Арно С, Чамес П, Перес С., Лакруа Э, Дюклер А, Эпинат Дж. С., Стрише Ф., Пети А.С., Патен А, Гийе С., Роллан С, Прието Дж., Бланко Ф.Дж., Браво Дж, Монтойя Дж., Серрано Л., Дюшато П. , Pâques F (январь 2006 г.). «Разработка большого количества высокоспецифичных эндонуклеаз самонаведения, которые индуцируют рекомбинацию на новых мишенях ДНК». Журнал молекулярной биологии. 355 (3): 443–58. Дои:10.1016 / j.jmb.2005.10.065. PMID 16310802.
- ^ Смит Дж., Гризо С., Арно С., Дюклер А., Эпинат Дж. К., Чамес П., Прието Дж., Редондо П., Бланко Ф. Дж., Браво Дж., Монтойя Дж., Пак Ф., Дюшато П. (2006). «Комбинаторный подход к созданию эндонуклеаз искусственного самонаведения, расщепляющих выбранные последовательности». Исследования нуклеиновых кислот. 34 (22): e149. Дои:10.1093 / нар / gkl720. ЧВК 1702487. PMID 17130168.
- ^ Маггерт К.А., Голич К.Г. (ноябрь 2005 г.). «Высокоэффективные обмены половых хромосом, вызванные экспрессией I-CreI у дрозофилы». Генетика. 171 (3): 1103–14. Дои:10.1534 / генетика.104.040071. ЧВК 1456814. PMID 16020774.