Планы тела докембрия - Precambrian body plans
Эта статья включает в себя список общих Рекомендации, но он остается в основном непроверенным, потому что ему не хватает соответствующих встроенные цитаты. (Май 2015 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |
До конца 1950-х гг. Докембрийский эпоха, как полагали, не принимала многоклеточный организмы. Однако с радиометрическое датирование техники, было обнаружено, что окаменелости первоначально найденный в Ediacara Hills в Южная Австралия относятся к позднему докембрию. Эти окаменелости - отпечатки тел организмов в форме дисков, листья и некоторые с лентами, которые, скорее всего, щупальца.
Это самые ранние многоклеточные организмы в истории Земли, несмотря на то, что одноклеточные существовали задолго до этого. Требования к многоклеточности были заложены в гены некоторых из этих клеток, в частности хоанофлагелляты. Считается, что они являются предшественниками всех животных. Они очень связаны с губки (Porifera), которые являются простейшими многоклеточными животными.
Чтобы понять переход к многоклеточности во время докембрия, важно взглянуть на требования к многоклеточности - как биологические, так и экологические.
Докембрийская эпоха
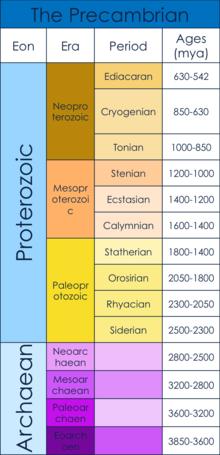
Докембрийская эра датируется от начала формирования Земли (4,6 миллиарда лет назад) до начала Кембрийский период, 542 миллиона лет назад.[1] Докембрий состоит из Hadean, Архей и Протерозойский эоны.[1] В частности, в этой статье исследуются Эдиакарский, когда, как полагают, возникли первые многоклеточные тела, а также то, что вызвало рост многоклеточности.[2] Этот временной период возник после Снежок Земля среднего неопротерозоя. «Земля-снежок» была периодом всемирного оледенение, который, как полагают, служил узкое место населения для последующего эволюция многоклеточных организмов.[3]
Докембрийские тела
Земля сформировалась около 4,6 миллиарда лет назад, а одноклеточная жизнь возникла несколько позже, после прекращения поздней тяжелой бомбардировки, периода интенсивных ударов астероидов, возможно, вызванных миграцией планет газовых гигантов на их нынешние орбиты. многоклеточность и тела - относительно недавнее событие в истории Земли.[4] Тела начали появляться ближе к концу Докембрийский Эра, во время Эдиакарский период. Окаменелости Эдиакарский период были впервые обнаружены в Южная Австралия в Ediacara Hills, отсюда и название. Однако первоначально считалось, что эти окаменелости являются частью Кембрийский и только в конце 1950-х годов Мартин Глесснер определили, что окаменелости действительно относятся к докембрийской эпохе. Окаменелости, которые были обнаружены, датируются примерно 600 миллионами лет назад и имеют различную морфологию.[4]
Окаменелости эдиакарских островов
Для получения дополнительной информации см. Эдиакарская биота.
Найденные окаменелости, относящиеся к докембрийской эпохе, не имеют четких структур, поскольку в этот период не было скелетных форм.[4] Скелеты не возникло до Кембрийский Период, когда кислород уровни увеличились. Это потому, что скелеты требуют коллаген, который использует Витамин С как кофактор, который требует кислород.[5] Для получения дополнительной информации о повышении содержания кислорода см. Раздел о кислороде. Большинство окаменелостей этой эпохи происходит от Ошибочная точка зрения на восточном побережье Канада или Ediacara Hills в Южная Австралия.[4]
Большинство окаменелостей найдено как отпечатки мягкотелых организмов в форме дисков, лент или листьев.[2][4] Существуют также следы окаменелостей, свидетельствующие о том, что некоторые из этих докембрийских организмов, скорее всего, были червеобразными существами, которые были локомотивами.[6] У большинства этих окаменелостей отсутствуют какие-либо узнаваемые головы, рты или органы пищеварения, и считается, что они питались посредством абсорбционных механизмов и симбиотических отношений с хемоавтотрофами (Хемотроф ), фотоавтотрофы (Фототроф ) или осмоавтотрофов.[1] Ленточные окаменелости напоминают организмы с щупальцами и, как полагают, питались, поимая добычу. В ветвь окаменелости напоминают морские ручки и другие книдарийцы. В следы окаменелостей предполагаю, что были кольчатые червя типа существ, а окаменелости диска напоминают губки. Несмотря на это сходство, большая часть идентификации является предположением, поскольку окаменелости не показывают очень четких структур. Другие окаменелости не похожи ни на какие известные родословные.[1]
Многие организмы, такие как Чарния, нашел в Ошибочная точка зрения, не были похожи на какие-либо организмы, наблюдаемые сегодня. У них были отдельные тела, но не хватало голова и пищеварительные области. Скорее их тело было организовано очень просто, фрактал -подобный узор ветвления.[7] Каждый элемент тела был мелко разветвлен и увеличивался за счет повторяющегося ветвления. Это позволило организму иметь большую площадь поверхности и максимизировать усвоение питательных веществ без необходимости во рту и пищеварительная система. Однако генетическая информация была минимальной, и поэтому не было требований, которые позволили бы им разработать более эффективные методы кормления. Это означает, что они, вероятно, были вытеснены другими организмами и, таким образом, вымерли.[7]
Организмы, обнаруженные на Эдиакарских холмах в Южной Австралии, имели либо радиально-симметричный план тела, либо один организм, Сприггина, отображена первая двусторонняя симметрия. Считается, что на Эдиакарских холмах когда-то было мелководье. риф где больше света могло проникнуть на дно океана. Это позволило больше разнообразие организмов. Найденные здесь организмы напоминают родственников книдарийцы, моллюски или же кольчатые червя.[7]
Чарния

Чарния окаменелости первоначально были найдены в Чарнвудский лес в Англия, отсюда и название Чарния.[7] Эти окаменелости принадлежат морским организмам, которые жили на дне океана. У окаменелостей есть фрактал В плане тела и имели форму вайя, то есть напоминали широколиственные растения, такие как папоротники. Однако они не могли быть растениями, поскольку обитали в темных глубинах океанского дна. В Чарнвудский лес, Чарния был обнаружен как изолированный вид, однако гораздо больше окаменелостей было найдено на восточном побережье Канады в районе Мистекен-Пойнт в Ньюфаундленде. Чарния был прикреплен ко дну океанского дна и сильно выровнен по течению. Это видно, потому что в нижней части экрана есть дискообразные формы. Чарния ископаемое, которое показывает где Чарния был привязан, и все ближайшие окаменелости обращены в одном направлении. Эти окаменелости на мысе Ошибок хорошо сохранились под вулканическим пеплом и слоями мягкой грязи.[7] Путем радиометрического датирования окаменелостей установлено, что Чарния должно быть, жили около 565 миллионов лет назад.[3][8]
Дикинсония

Дикинсония окаменелости - еще одна известная окаменелость эдиакарского периода, найденная в Южная Австралия и Россия.[9] Остается неизвестным, какой тип организма Дикинсония был; тем не менее, это считалось полихета, турбеллярий / кольчатые червь, медуза, полип, протест, лишайник или же гриб.[9] Они сохранились в кварцевых песчаниках и датируются примерно 550 миллионами лет назад. Дикинсония были мягкотелыми организмами, которые демонстрируют некоторые признаки очень медленного движения.[3] Есть слабые круглые отпечатки в скале, которые следуют по пути, а затем, следуя по тому же пути, появляется более определенный круглый отпечаток того же размера. Это указывает на то, что организм, вероятно, медленно перемещался из одной зоны кормления в другую и поглощал питательные вещества. Предполагается, что у этого организма, вероятно, были очень маленькие придатки, которые позволяли ему двигаться так же, как современные морские звезды.[10]
Spriggina

Spriggina окаменелости представляют собой первые известные организмы с двусторонне-симметричным планом тела. У них были голова, хвост и почти одинаковые половинки.[2] У них, вероятно, были органы чувств в голове и органы пищеварения в хвосте, что позволяло им более эффективно находить пищу. Они обладали способностью к передвижению, что давало им преимущество перед другими организмами той эпохи, которые были либо привязаны к дну океана, либо передвигались очень медленно. Spriggina был мягким телом, поэтому окаменелости оставались слабыми отпечатками. Скорее всего, это связано с кольчатыми червями, однако есть некоторые предположения, что это могло быть связано с членистоногими, поскольку оно несколько напоминает окаменелости трилобитов.[2][4]
Следы окаменелостей
В Эдиакарский окаменелости Южная Австралия содержат следы окаменелостей, которые указывают на наличие подвижных донных организмов. Все организмы, которые оставили следы в отложениях, были червеобразными питателями наносов или питателями детрита (Детритофаги ). Есть несколько следов окаменелостей, которые напоминают членистоногие тропы. Данные свидетельствуют о том, что организмы, похожие на членистоногих, существовали во время Докембрийский. Это свидетельство в виде оставленных позади следов; в частности, один экземпляр, который показывает шесть пар симметрично расположенных отпечатков, которые напоминают пешеходные тропы трилобитов.[6]
Переход от одноклеточности к многоклеточности
На протяжении большей части истории Земли жизнь была одноклеточный. Однако в одноклеточных организмах есть ингредиенты для возникновения многоклеточности. Несмотря на наличие ингредиентов для многоклеточности, организмы были ограничены из-за отсутствия благоприятных условий окружающей среды. Подъем кислорода ( Большое событие оксигенации ) привели организмы к развитию более сложных планов тела. Для возникновения многоклеточности организмы должны были быть способны к сотовая связь, агрегирование и специализированные функции. Переход к многоклеточности, положивший начало эволюции животных от простейшие - одно из самых малоизученных событий в истории. Понимание хоанофлагелляты и их отношение к губки важен при построении теорий происхождения многоклеточности[11]
Хоанофлагелляты
Хоанофлагелляты, также называемые «воротничковидными жгутиконосцами», представляют собой одноклеточные протисты, которые существуют как в пресных водах, так и в океанах.[12] Хоанофлагелляты имеют сферическое (или яйцевидное) тело клетки и жгутик, окруженный воротником, состоящим из микроворсинок актина.[12][13] В жгутик используется для облегчения движения и приема пищи. Поскольку жгутик удары, это принимает воды через микроворсинки, прикрепленные к воротнику, что помогает отфильтровывать нежелательные бактерии и другие крошечные еда частицы.[12] Хоанофлагелляты состоят примерно из 150 видов и размножаются простым делением.[14]
Хоанофлагеллята Salpingoeca rosetta
(также известный как Choanoflagellate Proterospongia)
Хоанофлагеллята Salpingoeca rosetta редкая пресноводная эукариот состоящий из ряда ячеек, заключенных в желеобразную матрицу. Этот организм демонстрирует очень примитивный уровень дифференциация клеток и специализация.[14] Это видно на флагеллированных клетках и их воротниковых структурах, которые перемещают клеточную колонию через воду, в то время как амебоидные клетки внутри служат для деления на новые клетки, чтобы способствовать росту колонии.
Аналогичный низкий уровень клеточная дифференциация и спецификации также можно увидеть на губках. У них также есть воротниковые клетки (также называемые хоаноциты из-за их сходства с хоанофлагеллятами) и амебоидные клетки, расположенные в студенистой матрице. В отличие от хоанофлагеллята Salpingoeca rosetta, у губок также есть другие типы клеток, которые могут выполнять различные функции (см. губки). Кроме того, воротничковые клетки губок бьются в каналах тела губки, тогда как Salpingoeca rosettaВоротничковые клетки расположены внутри, и в нем отсутствуют внутренние каналы. Несмотря на эти незначительные различия, есть веские доказательства того, что Proterospongia и Metazoa тесно связаны.[14]
Хоанофлагеллята Perplexa
Эти хоанофлагелляты могут прикрепляться друг к другу через соединение микроворсинок воротничка.[15]
Хоанофлагеллята Codosiga Botrytis и Desmerella
Эти хоанофлагелляты способны образовывать колонии через тонкие межклеточные мостики, которые позволяют отдельным клеткам прикрепляться. Эти мостики напоминают кольцевые каналы, которые связывают развивающиеся сперматогонии или оогонии у животных.[15]
Губки (Porifera)
Губки являются одними из самых старых и самых распространенных животных на Земле. Окаменелости спикул губок появились в докембрийскую эру около 580 миллионов лет назад.[16] Группа этих окаменелостей была обнаружена в формации Душанто в Южном Китае. Некоторые круглые отпечатки с Эдиакарских холмов в Южной Австралии, как сообщается, также являются губками. Это одна из немногих ветвей многоклеточных животных этой эпохи, которые продолжают существовать и остаются относительно неизменными.[16][17]Губки - такие успешные организмы из-за их простых, но эффективных морфология. У них нет рта, пищеварительной, нервной или кровеносной систем. Вместо этого они питатели-фильтры, что означает, что они получают пищу за счет питательных веществ, содержащихся в воде.[18] У них есть поры, называемые Остия, что вода проходит в камеру, называемую губчатый гель, и выходит через камеру, называемую оскулюм.[18] Благодаря этой системе фильтрации воды они получают питательные вещества, необходимые для их выживания. В частности, они внутриклеточно переваривают бактерии, микроводоросли или коллоиды.[19]
Скелеты губки состоят либо из губка или известковые и кремнистые спикулы с вкраплениями некоторых молекул коллагена.[20] Коллаген скрепляет клетки губки. Различные линии губок различаются в зависимости от состава их скелетов. Три основных класса губок: Demospongiae, Гексактинеллид, и Известковый.
Demonsponges - самый известный тип губок, поскольку они используются людьми. Они отличаются кремнистым скелетом из двух и четырехлучевых спикул и содержат белок спонгин.
Гексактинеллиды также называют стеклянными губками и отличаются шестилучевым стеклянным скелетом. Эти губки также способны выполнять потенциалы действия.
Известковые губки характеризуются каркасом из карбоната кальция и составляют менее 5% губок.[20]
Клетки
У губок есть около 6 различных типов клеток, которые могут выполнять разные функции.[20] Губки - хорошая модель для изучения происхождения многоклеточности, потому что клетки способны общаться друг с другом и повторно агрегироваться. В эксперименте, проведенном Генри Ван Питерс Уилсон в 1910 году было обнаружено, что клетки диссоциированных губок могут посылать сигналы и узнавать друг друга, чтобы сформировать новую особь.[21] Это говорит о том, что клетки, из которых состоят губки, способны к независимой жизни, однако, как только появилась возможность многоклеточности, объединение вместе в один организм было более эффективным способом жизни.
Наиболее заметными клеточными типами губок являются бокаловидные клетки, называемые хоаноциты, названные так из-за сходства с хоанофлагеллятами.[20] Сходство между этими двумя типами клеток заставляет ученых полагать, что хоанофлагелляты являются родственными таксонами метазоа. Жгутики этих клеток являются движущей силой движения воды через тело губки.[22] Клеточное тело хоаноцитов отвечает за усвоение питательных веществ. У некоторых видов эти клетки могут развиваться в гаметы.[20]
В Пинакоциты - это клетки на внешней стороне губки, выстилающие тело клетки. Они плотно упакованы и очень тонкие.[20]
В мезенхима выстилает область между пинакоцитами и хоаноцитами. Они содержат матрицу, состоящую из белков и спикул.[20]
Археоциты представляют собой особые типы клеток, поскольку они могут трансформироваться во все другие типы клеток. Они будут делать то, что необходимо организму губки, например принимать и переваривать пищу, транспортировать питательные вещества к другим клеткам тела губки. Эти клетки также способны развиваться в гаметы у некоторых видов губок.[20]
В склероциты отвечают за секрецию спикул. У видов губок, которые используют спонжин вместо известковых и кремнистых спикул, склероциты заменяются спонгоцитами, которые секретируют скелетные волокна спонгина.[20]
В миоциты и пороциты отвечают за сжатие губки. Эти сокращения аналогичны мышечным сокращениям у других организмов, поскольку у губок нет мышц. Они отвечают за регулирование потока воды через губку.[20]
Формирование многоклеточности
Формирование многоклеточности стало поворотным моментом в эволюции жизни на Земле. Вскоре после возникновения многоклеточности произошло огромное увеличение разнообразия живых организмов в начале XX века. Кембрийский Эра, названная Кембрийский взрыв. Считается, что многоклеточность развивалась на Земле несколько раз, потому что это была полезная жизненная стратегия для организмов.[23] Для возникновения многоклеточности клетки должны обладать способностью к саморепликации, межклеточной адгезии и межклеточной коммуникации. Также должен быть доступный кислород и давление отбора в окружающей среде.
Теория клеточного деления: С. Розетта.
Работа Fairclough, Dayel и King предполагает, что S. Rosetta может существовать либо в одноклеточной форме, либо в колониях из 4-50 клеток, которые образуют плотно связанные группы сфер.[15] Это было установлено путем выполнения эксперимент с интродукцией хищных бактерий Алгорифаг вида к образцу одноклеточной S. Rosetta организм и наблюдали за активностью в течение 12 часов. Результаты этого исследования продемонстрировали, что колонии клеток образовывались в результате деления исходной одиночной клетки S. Rosetta, а не в результате агрегации клеток. Дальнейшие исследования в поддержку теории разрастания клеток были проведены путем введения и удаления препарата. афидиколин который служит для блокировки деления клеток. Когда лекарство было введено, деление клеток прекращалось, и в результате агрегации клеток происходило образование колоний. Когда препарат, средство, медикамент был удален, снова доминировало деление клеток.[15]
Строительные блоки для клеточной адгезии
Глядя на геном из Хоанофлагеллята, "Monosiga brevicollis", ученые пришли к выводу, что хоанофлагелляты играют ключевую роль в развитии многоклеточности.[12] Николь Кинг проделал работу по изучению генома Monisiga brevicollis, и обнаружил ключевые белковые домены, общие для многоклеточных и хоанофлагеллят. Эти домены играют роль в клеточной передаче сигналов и процессах адгезии у многоклеточных животных. Открытие того, что у хоанофлагеллят также есть эти гены, является невероятным открытием, потому что ранее считалось, что только у многоклеточных животных есть гены, ответственные за межклеточную коммуникацию и агрегацию. Это говорит о том, что эти домены играют ключевую роль в происхождении многоклеточности, поскольку они связывают одноклеточный организм (хоанофлагелляты) с многоклеточными организмами (многоклеточные). Это показывает, что компоненты, необходимые для многоклеточности, присутствовали у общего предка между многоклеточными животными и хоанофлагеллятами.[12]
Сотовая сигнализация и сотовая связь
Ни губки, ни плакозоя Trichoplax adhaerens, похоже, не оснащены нейрон синапсы, однако оба они обладают несколькими факторами, связанными с одной и той же синаптической функцией.[24] Следовательно, вполне вероятно, что центральные особенности, участвующие в синаптической передаче, возникли на ранней стадии эволюции многоклеточных животных, скорее всего, примерно в то время, когда большая часть жизни на Земле переходила к многоклеточности. Было обнаружено, что Munc18 /синтаксин 1 комплекс мог бы быть важным компонентом для производства SNARE белок. Секреция SNARE белок из синаптические везикулы считается критически важным для нейронной коммуникации. Комплекс Munc18 / синтаксин 1, обнаруженный у M. brevicollis, структурно и функционально подобен комплексу многоклеточных животных. Это предполагает, что это важный шаг на пути реакции к SNARE сборка. Считается, что общий предок хоанофлагеллят и многоклеточных животных использовал этот первичный механизм секреции в качестве предшественника синаптической коммуникации. Этот механизм в конечном итоге будет использоваться для межклеточной коммуникации у животных.[24]
Причины развития многоклеточности
Несмотря на то, что прокариотические клетки содержат строительные блоки, необходимые для возникновения многоклеточности, этот переход не происходил в течение примерно 1500 миллионов лет после возникновения первой эукариотической клетки.[11] Ученые предложили две основные теории по той причине, что многоклеточность возникла так поздно после появления жизни на Земле.
Теория хищничества для многоклеточности
Эта теория постулирует, что многоклеточность возникла как средство спасения жертвы от хищников. Более крупная добыча с меньшей вероятностью станет жертвой, а более крупные хищники с большей вероятностью поймают добычу. Следовательно, вероятно, что многоклеточность возникла, когда появились первые хищники. Собираясь в более крупный многоклеточный организм, жертва могла избежать попыток хищника.[11] Следовательно, многоклеточность выборочно предпочиталась одноклеточности. Это можно увидеть в простом эксперименте, проведенном Boraas et al. (1998).[25] Когда хищный протист, Ochromonas valencia, был введен в популяцию жертв Хлорелла vulgarisбыло замечено, что менее чем за 100 поколений видов-жертв многоклеточная форма роста водорослей стала доминирующей. Это интересно, потому что до появления хищника численность населения из Хлорелла vulgaris сохранял одноклеточную форму роста на протяжении тысяч поколений. Вполне вероятно, что он оставался бы одноклеточным на неопределенное время, если бы не было введено давление отбора, которое было вызвано хищниками. После нескольких поколений с хищником вид водорослей сохранил форму роста из 8-10 клеток, которая была достаточно большой, чтобы избежать хищника, но достаточно маленькой, чтобы каждая клетка все еще имела доступ к питательным веществам.[25] Эти отношения хищник-жертва являются вероятной причиной того, почему для организмов было полезно быть многоклеточным.
Теория повышения уровня кислорода в многоклеточности
Несмотря на то, что у организмов был потенциал стать многоклеточным, вполне вероятно, что это было невозможно до позднего периода. Неопротерозойский. Это потому, что многоклеточность требует кислород, и до позднего Неопротерозойский было очень ограничено кислород доступность.[11] После плавления «Снежок Земля ”В середине Неопротерозойский, питательные вещества, которые были захвачены льдом, затопили океаны.[7] Выживание бактерии процветала из-за повышенного уровня питательных веществ. Среди этих микробов были цианобактерии и другие кислород производство бактерии, что привело к резкому повышению уровня кислорода. Повышенная доступность кислорода позволила клеткам использовать его для производства коллагена. Коллаген является ключевым компонентом агрегации клеток. Это веревочная молекула, которая «связывает» клетки вместе. Кислород требуется для коллаген синтез потому что аскорбиновая кислота (Витамин С ) необходим для этого процесса.[5] Ключевым компонентом молекулы аскорбиновой кислоты является кислород (химическая формула C6ЧАС8О6).[26] Таким образом, очевидно, что повышение содержания кислорода является решающим шагом к повышению многоклеточность поскольку это важно для синтез коллагена.[7]
Строительные блоки найдены как в губках, так и в людях
Коллаген
Коллаген является наиболее распространенным белком у млекопитающих и является важной молекулой в формировании костей, кожи и других соединительных тканей. Различные типы коллагена были обнаружены во всех многоклеточных организмах, включая губки.
Было обнаружено, что губки действительно имеют последовательность гена, кодирующую коллаген IV типа, что является диагностическим признаком базальная пластинка.[27]
Также было обнаружено, что 29 типов коллаген было обнаружено, что они существуют у людей. Эту обширную группу можно в дальнейшем разделить на несколько семейств в соответствии с их первичной структурой и надмолекулярной организацией. Среди многих типов коллагенов только фибриллярные коллагены и коллагены базальной мембраны (тип IV) были обнаружены у губок и книдарий, которые являются двумя самыми ранними ветвящимися клонами многоклеточных животных. Исследования были сосредоточены на происхождении фибриллярных молекул коллагена. У губок существует три клады фибриллярных молекул, A, B и C. Предполагается, что только фибриллярные коллагены B клады сохранили свою характерную модульную структуру от губки до человека.[28]
У млекопитающих фибриллярные коллагены, участвующие в образовании поперечно-полосатых фибрилл, относятся к типам I – III, V и XI. Коллагены типа II и типа XI составляют фибриллы присутствует в хрящ. Их можно отличить от коллагенов, находящихся в нехрящевых тканях, которые включают коллагены I, III и V.[28]
Протеин
Дополнительное исследование белков губок показало, что из 42 проанализированных белков губок все они имели гомологичные белки, обнаруженные у людей. Оценка идентичности 53% была дана сходству между белками губки и человека, по сравнению с показателем 42%, когда та же последовательность сравнивалась с последовательностью C. elegans.[29]
Рекомендации
- ^ а б c d Валентин, J.W (1994). «Позднедокембрийские билатерии: сорта и клады». Труды Национальной академии наук. 91 (15): 6751–6757. Bibcode:1994PNAS ... 91,6751V. Дои:10.1073 / пнас.91.15.6751. ЧВК 44279. PMID 8041693.
- ^ а б c d Эрвин, Дуглас; Валентин, Джеймс; Яблонски, Дэвид (1997). «Происхождение планов тела: недавние находки окаменелостей и новые взгляды на развитие животных дают свежий взгляд на загадку взрыва животных в раннем кембрии». Американский ученый. 85: 126–137.
- ^ а б c Нарбонна, Гай; Гелинг, Джеймс (2003). "Жизнь после снежного кома: древнейшие сложные эдиакарские окаменелости" (PDF). Геология. 31 (1): 27–30. Bibcode:2003Гео .... 31 ... 27N. Дои:10.1130 / 0091-7613 (2003) 031 <0027: lastoc> 2.0.co; 2. Архивировано из оригинал (PDF) на 2004-10-31.
- ^ а б c d е ж Глесснер, Мартин (1959). «Древнейшие ископаемые фауны Южной Австралии» (PDF). Geologische Rundschau. 47 (2): 522–531. Bibcode:1959ГеоРу..47..522Г. Дои:10.1007 / bf01800671. S2CID 140615593.
- ^ а б Тоу, Кеннет (1970). «Приоритет кислород-коллаген и ископаемые остатки ранних метазоа». Труды Национальной академии наук. 65 (4): 781–788. Bibcode:1970ПНАС ... 65..781Т. Дои:10.1073 / pnas.65.4.781. ЧВК 282983. PMID 5266150.
- ^ а б Глесснер, Мартин (1969). «Следы окаменелостей из докембрия и базального кембрия». Lethaia. 2 (4): 369–393. Дои:10.1111 / j.1502-3931.1969.tb01258.x.
- ^ а б c d е ж грамм Аттенборо, Дэвид. «Первая жизнь». YouTube.
- ^ Антклифф, Джонатан; Бразье, Мартин (2008). «Чарния в 50: модели развития эдиакарских земель». Палеонтология. 51 (1): 1475–4983. Дои:10.1111 / j.1475-4983.2007.00738.x.
- ^ а б Retallack, Грегори (2007). «Рост, распад и уплотнение захоронения Дикинсонии, культовой ископаемой эдиакарской останки». Алчеринга. 31 (3): 215–240. Дои:10.1080/03115510701484705. S2CID 17181699.
- ^ Сперлинг, Эрик; Винтер, Якоб (2010). «Сродство плакозоя к Dickinsonia и эволюция позднепротерозойских режимов питания многоклеточных». Эволюция и развитие. 12 (2): 201–209. Дои:10.1111 / j.1525-142X.2010.00404.x. PMID 20433459. S2CID 38559058.
- ^ а б c d Король, Николь (сентябрь 2004 г.). "Одноклеточные предки развития животных" (PDF). Клетка развития. 7 (3): 313–325. Дои:10.1016 / j.devcel.2004.08.010. PMID 15363407.
- ^ а б c d е Король, Николь; и другие. (2008). «Геном хоанофлагеллаты Monosiga brevicollis и происхождение многоклеточных животных». Природа. 451 (7180): 783–788. Bibcode:2008Натура.451..783K. Дои:10.1038 / природа06617. ЧВК 2562698. PMID 18273011.
- ^ Король, Николь; Кэррол, Шон Б. (10 сентября 2001 г.). «Рецепторная тирозинкиназа из хоанофлагеллят: молекулярное понимание ранней эволюции животных». Труды Национальной академии наук Соединенных Штатов Америки. 98 (26): 15032–15037. Bibcode:2001ПНАС ... 9815032К. Дои:10.1073 / pnas.261477698. ЧВК 64978. PMID 11752452.
- ^ а б c Ваггонер, Бен. «Знакомство с Choanoflagellata: где все началось для животных». UCMP Беркли. Беркли.
- ^ а б c d Дайел, Марк; Алегадо, Розанна; Fairclough, Стивен; Левин, Тера; Николс, Скотт; Макдональд, Кент; Король, Николь (сентябрь 2011 г.). «Дифференцировка и морфогенез клеток в колониеобразующей хоанофлагелляте Salpingoeca rosetta». Биология развития. 357 (1): 73–82. Дои:10.1016 / j.ydbio.2011.06.003. ЧВК 3156392. PMID 21699890.
- ^ а б Ли, Чиа-Вэй; Чен, Цзюнь-Юань; Хуа, Цзы-Эн (февраль 1998 г.). «Докембрийские губки с клеточными структурами». Наука. 279 (5352): 879–882. Bibcode:1998Sci ... 279..879L. Дои:10.1126 / science.279.5352.879. PMID 9452391.
- ^ Гелинг, Джеймс; Ригби, Кит (1996). «Долгожданные губки из неопротерозойской фауны Ediacara Южной Австралии». Журнал палеонтологии. 70 (2): 185–195. Дои:10.1017 / S0022336000023283. JSTOR 1306383.
- ^ а б Коллинз, Аллен Дж .; Ваггонер, Бен. "Porifera: Подробнее о морфологии". UCMP Беркли. Беркли.
- ^ Дюпон, Самуэль; Корре, Эрван; Ли, Яньян; Вэселет, Жан; Бурге-Кондраки, Мария-Лиз (декабрь 2013 г.). «Первые сведения о микробиоме хищной губки». FEMS Microbiology Ecology. 86 (3): 520–531. Дои:10.1111/1574-6941.12178. PMID 23845054.
- ^ а б c d е ж грамм час я j Коллинз, Аллен Г. "Porifera: клетки". UCMP Беркли. Беркли.
- ^ Ларру, Клэр; и другие. (2006). «Развитие экспрессии генов факторов транскрипции в демоспонге: понимание происхождения многоклеточности многоклеточных». Эволюция и развитие. 8 (2): 150–173. Дои:10.1111 / j.1525-142x.2006.00086.x. PMID 16509894. S2CID 36836333.
- ^ Майерс, Фил. «Губки Porifera». Сеть разнообразия животных.
- ^ Иваса, Джанет; Шостак, Джек. «Хронология эволюции жизни». Изучение истоков жизни.
- ^ а б Бакхардт, Павел; и другие. (2 августа 2011 г.). «Изначальный нейросекреторный аппарат, идентифицированный у хоанофлагеллаты Monosiga brevicollis». Труды Национальной академии наук Соединенных Штатов Америки. 108 (37): 15264–15269. Bibcode:2011PNAS..10815264B. Дои:10.1073 / pnas.1106189108. ЧВК 3174607. PMID 21876177.
- ^ а б Бораас, Мартин; Сил, Дайанн; Боксхорн, Джозеф (1998). «Фаготрофия жгутиконосцем выбирает колониальную добычу: возможное происхождение многоклеточности». Эволюционная экология. 12 (2): 153–164. Дои:10.1023 / А: 1006527528063. S2CID 12298325.
- ^ Найду, Ахилендер (2003). «Витамин С для здоровья и болезней человека все еще остается загадкой? Обзор». Журнал питания. 2 (7): 7. Дои:10.1186/1475-2891-2-7. ЧВК 201008. PMID 14498993.
- ^ Тайлер, Сет (2003). «Эпителий - основной строительный блок для многокомпонентной сложности». Интегративная и сравнительная биология. 43 (1): 55–63. Дои:10.1093 / icb / 43.1.55. PMID 21680409.
- ^ а б Экспозито, Жан-Ив; Ларру, Клэр; Клюзел, Кэролайн; Валькур, Ульрих; Летиас, Клэр; Дегнан, Бернар (2008). «Разнообразие фибриллярного коллагена демоспуба и морского анемона свидетельствует о раннем появлении кладок A / C и поддержании модульной структуры коллагенов типа V / XI от губки к человеку». Журнал биологической химии. 283 (42): 28226–28235. Дои:10.1074 / jbc.M804573200. ЧВК 2661393. PMID 18697744.
- ^ Гамулин, Вера; Мюллер, Изабель; Мюллер, Вернер (2008). «Белки губки больше похожи на белки Homo sapiens, чем на Caenorhabditis elegans». Биологический журнал Линнеевского общества. 71 (4): 821–828. Дои:10.1111 / j.1095-8312.2000.tb01293.x.