Эмбриоидное тело - Embryoid body
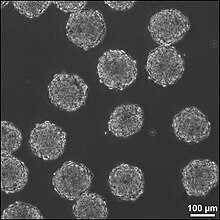

Эмбриоидные тела (EB) представляют собой трехмерные агрегаты плюрипотентных стволовые клетки.
EB представляют собой дифференцировку эмбриональных стволовых клеток человека в эмбриональные тельца, нарушающие три эмбриональных зародышевых листка.
Фон
Типы плюрипотентных клеток, которые составляют эмбриоидные тельца, включают: эмбриональные стволовые клетки (ESC) полученные из бластоциста стадия эмбрионов мыши (mESC),[1][2] примат[3] и человека (hESC)[4] источники. Кроме того, EB могут быть сформированы из эмбриональных стволовых клеток, полученных с помощью альтернативных методов, включая соматические клетки. передача ядер[5][6][7] или перепрограммирование соматических клеток для получения индуцированные плюрипотентные стволовые клетки (ИПС).[8][9][10][11] Подобно регуляторам скорости культивированный в монослой форматах ЭСК внутри эмбриоидных тел подвергаются дифференцировке и клеточной спецификации по трем зародышевые линии - энтодерма, эктодерма, и мезодерма - которые включают все соматический типы клеток.[12][13]
Однако, в отличие от однослойных культур, сфероидные структуры, которые образуются при агрегировании ESC, позволяют неадгезивную культуру EB в суспензии, что делает культуры EB по своей природе масштабируемыми, что полезно для подходов к биопроцессингу, в результате чего можно получать большие урожаи клеток для потенциальные клинические применения.[14] Кроме того, хотя EB в значительной степени демонстрируют гетерогенные паттерны дифференцированных типов клеток, ESC способны реагировать на аналогичные сигналы, которые направляют эмбриональное развитие.[15] Таким образом, трехмерная структура, в том числе создание сложных клеточные адгезии и паракрин сигнализация внутри микросреды БЭ,[16] позволяет дифференцировать и морфогенез что дает микроткани, похожие на структуры нативных тканей. Такие микроткани перспективны для непосредственного[15] или косвенно[17][18] восстанавливают поврежденные или больные ткани в применениях регенеративной медицины, а также для тестирования in vitro в фармацевтической промышленности и в качестве модели эмбрионального развития.
Формирование
EB формируются гомофильное связывание Са2 + зависимой молекулы адгезии E-кадгерин, который сильно выражен на недифференцированных ЭСК.[19][20][21] При культивировании как единичных клеток в отсутствие факторов, препятствующих дифференцировке, ESC спонтанно агрегируют с образованием EB.[19][22][23][24] Такое самопроизвольное образование часто достигается в объемных суспензионных культурах, когда чашка покрывается неадгезивными материалами, такими как агар или гидрофильные полимеры, чтобы способствовать предпочтительной адгезии между отдельными клетками, а не с субстратом культуры. Поскольку hESC подвергаются апоптозу при культивировании в виде отдельных клеток, образование EB часто требует применения ингибиторов rho-ассоциированная киназа (ROCK) путь, включая небольшие молекулы Y-27632[25] и 2,4 дизамещенный тиазол (Thiazovivin / Tzv).[26] Альтернативно, чтобы избежать диссоциации на отдельные клетки, EB могут быть сформированы из hESC путем ручного разделения прилипших колоний (или областей колоний) и впоследствии культивированы в суспензии. Формирование EB в суспензии поддается образованию больших количеств EB, но обеспечивает небольшой контроль над размером образующихся агрегатов, часто приводя к большим EB неправильной формы. В качестве альтернативы гидродинамический силы, передаваемые в платформы для смешанных культур, увеличивают однородность размеров EB, когда ESC инокулируют в объемных суспензиях.[27]
Образование EB также можно более точно контролировать путем инокуляции клеток известной плотности в отдельные капли (10–20 мкл), подвешенные на крышке чашки Петри, известные как висячие капли.[21] Хотя этот метод позволяет контролировать размер EB путем изменения количества клеток на каплю, образование висящих капель является трудоемким и нелегко поддается масштабированию культур. Кроме того, среда не может быть легко заменена в рамках традиционного формата висящих капель, что требует переноса висящих капель в объемные суспензионные культуры через 2–3 дня формирования, в результате чего отдельные EB имеют тенденцию к агломерации. Недавно были разработаны новые технологии, позволяющие обмениваться мультимедийными данными в модифицированном формате висячей капли.[28] Кроме того, были разработаны технологии для физического разделения клеток путем принудительной агрегации ESC в отдельных лунках или на адгезивных субстратах.[29][30][31][32] что позволяет повысить производительность и контролировать формирование ЭП. В конечном итоге методы, используемые для формирования EB, могут влиять на гетерогенность популяций EB с точки зрения кинетики агрегации, размера и выхода EB, а также траекторий дифференциации.[31][33][34]
Дифференциация внутри EB
В контексте ESC дифференциация протоколах, образование БЭ часто используется как метод инициирования спонтанной дифференциации в сторону трех зародышевые линии. Дифференцировка EB начинается со спецификации внешних клеток по направлению к фенотипу примитивной энтодермы.[35][36] Затем клетки снаружи откладываются внеклеточный матрикс (ECM), содержащий коллаген IV и ламинин,[37][38] похож на состав и структуру базальная мембрана. В ответ на отложение ECM EB часто образуют кистозную полость, при этом клетки, контактирующие с базальной мембраной, остаются жизнеспособными, а клетки внутри подвергаются апоптозу, в результате чего образуется заполненная жидкостью полость, окруженная клетками.[39][40][41] Последующая дифференциация приводит к образованию производных от трех зародышевых линий. В отсутствие добавок дифференцировка ЭСК по умолчанию в основном направлена на эктодерму и последующую нейронные линии.[42] Однако альтернативные медиа-композиции, включая использование фетальная бычья сыворотка а также определенные добавки факторов роста, были разработаны, чтобы способствовать дифференциации в сторону мезодерма и энтодерма родословные.[43][44][45]
В результате трехмерной структуры EB во время дифференцировки EB происходит сложный морфогенез, включающий появление популяций эпителиальных и мезенхимальных клеток, а также появление маркеров, связанных с эпителиально-мезенхимальный переход (ЕМТ).[46][47] Кроме того, индуктивные эффекты, возникающие в результате передачи сигналов между популяциями клеток в EB, приводят к пространственным и временным изменениям, которые способствуют сложным морфогенез.[48] Внутри БЭ часто проявляются тканеподобные структуры, включая появление островков крови, напоминающих ранние структуры кровеносных сосудов в развивающемся эмбрионе, а также формирование паттерна нейрит расширений (указывающих на организацию нейронов) и спонтанной сократительной активности (указывающих на кардиомиоцит дифференциация), когда EB наносятся на клейкие подложки, такие как желатин.[13] Совсем недавно сложные структуры, в том числе структуры, подобные оптическим чашечкам, были созданы in vitro в результате дифференцировки EB.[49]
Параллели с эмбриональным развитием
Большая часть исследований, имеющих важное значение для дифференцировки и морфогенеза эмбриональных стволовых клеток, происходит из исследований в области биологии развития и эмбриогенеза млекопитающих.[15] Например, сразу после бластоциста стадии развития (из которой происходят ЭСК) эмбрион претерпевает гаструляция, посредством чего клеточная спецификация внутренняя клеточная масса приводит к образованию висцеральной энтодермы и эпибласт.[50] Поскольку передне-задняя ось формируется, у эмбриона развивается временная структура, известная как примитивная полоса.[51] Большая часть пространственного паттерна, который происходит во время формирования и миграции примитивной полоски, является результатом секреции агонисты и антагонисты различными популяциями клеток, включая факторы роста из Wnt и трансформирующий фактор роста β (TGFβ) (Lefty 1, Nodal), а также репрессоры тех же молекул (Dkk-1, Sfrp1, Sfrp5).[52][53][54] Из-за сходства между эмбриогенезом и дифференцировкой ESC многие из тех же факторов роста являются центральными для подходов к направленной дифференцировке.
Кроме того, развитие культуры EB привело к развитию эмбриональные органоиды (гаструлоиды) которые показывают замечательные параллели с эмбриональным развитием[55][56][57][58][59] такие как нарушение симметрии, локализованные брахьюри выражение, формирование эмбриональные топоры (переднезадний, дорсовентральный и лево-правый) и движения, подобные гаструляции.[55][56][57]
Проблемы на пути к дифференциации
В отличие от дифференцировки ESCs в монослойных культурах, посредством чего добавление растворимых морфогенов и внеклеточное микроокружение может точно и однородно контролироваться, трехмерная структура EBs создает проблемы для направленной дифференцировки.[16][60] Например, популяция висцеральной энтодермы, которая образует внешнюю часть БЭ, создает внешнюю «оболочку», состоящую из плотно связанных эпителиальный -подобные клетки, а также плотный ECM.[61][62] Из-за таких физических ограничений в сочетании с размером EB, транспорт ограничения возникают внутри EB, создавая градиенты морфогенов, метаболитов и питательных веществ.[60] Было подсчитано, что транспорт кислорода ограничен в клеточных агрегатах диаметром более 300 мкм;[63] однако на развитие таких градиентов также влияют размер молекул и скорость поглощения клетками. Следовательно, доставка морфогенов к EB приводит к повышенной гетерогенности и снижению эффективности популяций дифференцированных клеток по сравнению с монослойными культурами. Одним из методов решения транспортных ограничений внутри EB является доставка морфогенов изнутри структуры EB.[61][64][65] Кроме того, EB можно культивировать как отдельные микроткани и впоследствии собирать в более крупные структуры для применения в тканевой инженерии.[66] Хотя сложность, возникающая из-за трехмерных адгезий и передачи сигналов, может повторять более естественные тканевые структуры,[67][68] это также создает проблемы для понимания относительного вклада механических, химических и физических сигналов в результирующие клеточные фенотипы и морфогенез.
Смотрите также
Рекомендации
- ^ Мартин, Г. Р. (1981). «Выделение плюрипотентной клеточной линии из ранних эмбрионов мыши, культивированных в среде, кондиционированной стволовыми клетками тератокарциномы». Труды Национальной академии наук Соединенных Штатов Америки. 78 (12): 7634–7638. Дои:10.1073 / pnas.78.12.7634. ЧВК 349323. PMID 6950406.
- ^ Evans, M. J .; Кауфман, М. Х. (1981). «Создание в культуре плюрипотенциальных клеток из эмбрионов мыши». Природа. 292 (5819): 154–156. Дои:10.1038 / 292154a0. PMID 7242681.
- ^ Thomson, J. A .; Калишман, Дж .; Голос, Т. Г .; Дурнинг, М .; Harris, C.P .; Becker, R.A .; Хирн, Дж. П. (1995). «Выделение линии эмбриональных стволовых клеток приматов». Труды Национальной академии наук Соединенных Штатов Америки. 92 (17): 7844–7848. Дои:10.1073 / пнас.92.17.7844. ЧВК 41242. PMID 7544005.
- ^ Thomson, J. A .; Itskovitz-Eldor, J .; Шапиро, С. С .; Вакниц, М. А .; Swiergiel, J. J .; Маршалл, В. С .; Джонс, Дж. М. (1998). «Линии эмбриональных стволовых клеток, полученные из бластоцист человека». Наука. 282 (5391): 1145–1147. Дои:10.1126 / science.282.5391.1145. PMID 9804556.
- ^ Briggs, R .; Кинг, Т. Дж. (1952). «Трансплантация живых ядер из клеток бластулы в энуклеированные яйца лягушек». Труды Национальной академии наук Соединенных Штатов Америки. 38 (5): 455–463. Дои:10.1073 / pnas.38.5.455. ЧВК 1063586. PMID 16589125.
- ^ Вильмут, И.; Schnieke, A.E .; McWhir, J .; Добрый, А. Дж .; Кэмпбелл, К. Х. С. (1997). «Жизнеспособное потомство, полученное из клеток плода и взрослых млекопитающих». Природа. 385 (6619): 810–813. Дои:10.1038 / 385810a0. PMID 9039911.
- ^ Munsie, M. J .; Michalska, A.E .; O'Brien, C.M .; Trounson, A. O .; Pera, M. F .; Маунтфорд, П. С. (2000). «Выделение плюрипотентных эмбриональных стволовых клеток из перепрограммированных ядер соматических клеток взрослых мышей». Текущая биология. 10 (16): 989–992. Дои:10.1016 / s0960-9822 (00) 00648-5. PMID 10985386.
- ^ Takahashi, K .; Яманака, С. (2006). «Индукция плюрипотентных стволовых клеток из эмбриональных и взрослых культур фибробластов мышей с помощью определенных факторов». Клетка. 126 (4): 663–76. Дои:10.1016 / j.cell.2006.07.024. HDL:2433/159777. PMID 16904174.
- ^ Takahashi, K .; Tanabe, K .; Охнуки, М .; Нарита, М .; Ichisaka, T .; Tomoda, K .; Яманака, С. (2007). «Индукция плюрипотентных стволовых клеток из фибробластов взрослого человека с помощью определенных факторов». Клетка. 131 (5): 861–872. Дои:10.1016 / j.cell.2007.11.019. HDL:2433/49782. PMID 18035408.
- ^ Yu, J .; Водяник, М. А .; Smuga-Otto, K .; Antosiewicz-Bourget, J .; Frane, J. L .; Tian, S .; Nie, J .; Jonsdottir, G.A .; Ruotti, V .; Стюарт, Р .; Слуквин, И. И .; Томсон, Дж. А. (2007). «Индуцированные линии плюрипотентных стволовых клеток, полученные из соматических клеток человека». Наука. 318 (5858): 1917–1920. Дои:10.1126 / science.1151526. PMID 18029452.
- ^ Парк, И. Х .; Arora, N .; Huo, H .; Maherali, N .; Ahfeldt, T .; Shimamura, A .; Lensch, M. W .; Cowan, C .; Hochedlinger, K .; Дейли, Г.К. (2008). "Плюрипотентные стволовые клетки, индуцированные конкретным заболеванием". Клетка. 134 (5): 877–886. Дои:10.1016 / j.cell.2008.07.041. ЧВК 2633781. PMID 18691744.
- ^ Itskovitz-Eldor, J .; Schuldiner, M .; Karsenti, D .; Eden, A .; Yanuka, O .; Amit, M .; Soreq, H .; Бенвенисти, Н. (2000). «Дифференциация эмбриональных стволовых клеток человека в эмбриональные тельца, нарушающие три эмбриональных зародышевых листка». Молекулярная медицина (Кембридж, Массачусетс). 6 (2): 88–95. ЧВК 1949933. PMID 10859025.
- ^ а б Doetschman, T. C .; Eistetter, H .; Кац, М .; Schmidt, W .; Кемлер Р. (1985). «Развитие in vitro линий эмбриональных стволовых клеток, полученных из бластоцист: формирование висцерального желточного мешка, островков крови и миокарда». Журнал эмбриологии и экспериментальной морфологии. 87: 27–45. PMID 3897439.
- ^ Dang, S.M .; Gerecht-Nir, S .; Chen, J .; Itskovitz-Eldor, J .; Зандстра, П. В. (2004). «Контролируемая масштабируемая культура дифференцировки эмбриональных стволовых клеток». Стволовые клетки. 22 (3): 275–282. Дои:10.1634 / стволовые клетки.22-3-275. PMID 15153605.
- ^ а б c Murry, C.E .; Келлер, Г. (2008). «Дифференциация эмбриональных стволовых клеток для клинически значимых популяций: уроки эмбрионального развития». Клетка. 132 (4): 661–680. Дои:10.1016 / j.cell.2008.02.008. PMID 18295582.
- ^ а б Bratt-Leal, A. S. M .; Карпенедо, Р. Л .; Макдевитт, Т. К. (2009). «Разработка микросреды эмбриоидного тела для прямой дифференцировки эмбриональных стволовых клеток». Прогресс биотехнологии. 25 (1): 43–51. Дои:10.1002 / btpr.139. ЧВК 2693014. PMID 19198003.
- ^ Nair, R .; Шукла, С .; Макдевитт, Т. К. (2008). «Бесклеточные матрицы, полученные из дифференцирующихся эмбриональных стволовых клеток». Журнал исследований биомедицинских материалов, часть A. 87A (4): 1075–1085. Дои:10.1002 / jbm.a.31851. PMID 18260134.
- ^ Baraniak, P.R .; Макдевитт, Т. К. (2010). «Паракринное действие стволовых клеток и регенерация тканей». Регенеративная медицина. 5 (1): 121–143. Дои:10.2217 / rme.09.74. ЧВК 2833273. PMID 20017699.
- ^ а б Куросава, Х. (2007). «Методы индукции образования эмбриоидных тел: система дифференцировки эмбриональных стволовых клеток in vitro». Журнал биологии и биоинженерии. 103 (5): 389–398. Дои:10.1263 / jbb.103.389. PMID 17609152.
- ^ Larue, L .; Antos, C .; Butz, S .; Huber, O .; Delmas, V .; Dominis, M .; Кемлер, Р. (1996). «Роль кадгеринов в формировании тканей». Разработка. 122 (10): 3185–3194. PMID 8898231.
- ^ а б Юн, Б. С .; Ю, С. Дж .; Lee, J. E .; Вы, С .; Lee, H.T .; Юн, Х.С. (2006). «Повышенная дифференцировка эмбриональных стволовых клеток человека в кардиомиоциты путем комбинирования культуры висячей капли и лечения 5-азацитидином». Дифференциация. 74 (4): 149–159. Дои:10.1111 / j.1432-0436.2006.00063.x. PMID 16683985.
- ^ Park, J. H .; Kim, S.J .; О, Э. Дж .; Moon, S. Y .; Roh, S. I .; Kim, C.G .; Юн, Х.С. (2003). «Создание и поддержание эмбриональных стволовых клеток человека на STO, постоянно растущей клеточной линии». Биология размножения. 69 (6): 2007–2014. Дои:10.1095 / биолрепрод.103.017467. PMID 12930726.
- ^ Williams, R.L .; Хилтон, Д. Дж .; Pease, S .; Willson, T. A .; Stewart, C.L .; Gearing, D. P .; Вагнер, Э. Ф .; Metcalf, D .; Nicola, N.A .; Гоф, Н. М. (1988). «Фактор ингибирования миелоидной лейкемии поддерживает потенциал развития эмбриональных стволовых клеток». Природа. 336 (6200): 684–687. Дои:10.1038 / 336684a0. PMID 3143916.
- ^ Ludwig, T. E .; Левенштейн, М. Э .; Jones, J.M .; Berggren, W. T .; Mitchen, E. R .; Frane, J. L .; Crandall, L.J .; Daigh, C.A .; Conard, K. R .; Piekarczyk, M. S .; Llanas, R.A .; Томсон, Дж. А. (2006). «Получение эмбриональных стволовых клеток человека в определенных условиях». Природа Биотехнологии. 24 (2): 185–187. Дои:10.1038 / nbt1177. PMID 16388305.
- ^ Watanabe, K .; Ueno, M .; Kamiya, D .; Нишияма, А .; Matsumura, M .; Wataya, T .; Takahashi, J. B .; Nishikawa, S .; Nishikawa, S.I .; Muguruma, K .; Сасай, Ю. (2007). «Ингибитор ROCK обеспечивает выживание диссоциированных эмбриональных стволовых клеток человека». Природа Биотехнологии. 25 (6): 681–686. Дои:10.1038 / nbt1310. PMID 17529971.
- ^ Xu, Y .; Чжу, X .; Hahm, H. S .; Wei, W .; Hao, E .; Хайек, А .; Дин, С. (2010). «Выявление основного механизма регуляции передачи сигналов для выживания плюрипотентных стволовых клеток и самообновления с помощью малых молекул». Труды Национальной академии наук. 107 (18): 8129–8134. Дои:10.1073 / pnas.1002024107. ЧВК 2889586. PMID 20406903.
- ^ Карпенедо, Р. Л .; Sargent, C. Y .; Макдевитт, Т. С. (2007). «Ротационная суспензионная культура повышает эффективность, урожайность и однородность дифференциации эмбриоидных тел». Стволовые клетки. 25 (9): 2224–2234. Дои:10.1634 / стволовые клетки.2006-0523. PMID 17585171.
- ^ Tung, Y.C .; Hsiao, A. Y .; Allen, S.G .; Torisawa, Y. S .; Хо, М .; Такаяма, С. (2011). «Высокопроизводительная 3D-культура сфероидов и тестирование лекарств с использованием массива 384 висящих капель». Аналитик. 136 (3): 473–478. Дои:10.1039 / c0an00609b. ЧВК 7454010. PMID 20967331. S2CID 35415772.
- ^ Park, J .; Cho, C.H .; Parashurama, N .; Li, Y .; Berthiaume, F. O .; Тонер, М .; Tilles, A.W .; Ярмуш, М. Л. (2007). «Модуляция дифференцировки эмбриональных стволовых клеток на основе микротехнологий». Лаборатория на чипе. 7 (8): 1018–1028. Дои:10.1039 / b704739h. PMID 17653344.
- ^ Mohr, J.C .; Де Пабло, Дж. Дж .; Палецек, С. П. (2006). «Трехмерная микропланшетная культура эмбриональных стволовых клеток человека». Биоматериалы. 27 (36): 6032–6042. Дои:10.1016 / j.biomaterials.2006.07.012. PMID 16884768.
- ^ а б Hwang, Y. -S .; Chung, B.G .; Ортманн, Д .; Hattori, N .; Moeller, H. -C .; Хадемхоссейни, А. (2009). «Микро-опосредованный контроль размера эмбрионального тела регулирует судьбу эмбриональных стволовых клеток посредством дифференциальной экспрессии WNT5a и WNT11». Труды Национальной академии наук. 106 (40): 16978–16983. Дои:10.1073 / pnas.0905550106. ЧВК 2761314. PMID 19805103.
- ^ Ungrin, M.D .; Joshi, C .; Nica, A .; Bauwens, C.L .; Зандстра, П. В. (2008). Каллетс, Патрик (ред.). «Воспроизводимое сверхвысокопроизводительное формирование многоклеточной организации из агрегатов человеческих эмбриональных стволовых клеток, полученных из суспензии единичных клеток». PLOS ONE. 3 (2): e1565. Дои:10.1371 / journal.pone.0001565. ЧВК 2215775. PMID 18270562.
- ^ Sargent, C. Y .; Berguig, G. Y .; Макдевитт, Т. К. (2009). «Кардиомиогенная дифференцировка эмбриоидных тел способствует культуре вращающейся орбитальной суспензии». Тканевая инженерия, часть А. 15 (2): 331–342. Дои:10.1089 / ten.tea.2008.0145. PMID 19193130.
- ^ Bauwens, C. L. L .; Peerani, R .; Niebruegge, S .; Вудхаус, К. А .; Кумачева, Е .; Husain, M .; Зандстра, П. В. (2008). "Контроль колоний человеческих эмбриональных стволовых клеток и неоднородность совокупного размера влияет на траектории дифференциации". Стволовые клетки. 26 (9): 2300–2310. Дои:10.1634 / стволовые клетки.2008-0183. PMID 18583540.
- ^ Chen, Y .; Li, X .; Eswarakumar, V.P .; Seger, R .; Лонаи, П. (2000). «Передача сигналов фактора роста фибробластов (FGF) через PI 3-киназу и Akt / PKB необходима для дифференцировки эмбриоидных тел». Онкоген. 19 (33): 3750–3756. Дои:10.1038 / sj.onc.1203726. PMID 10949929.
- ^ Эснер, М .; Pachernik, J .; Hampl, A .; Дворжак, П. (2002). «Целенаправленное нарушение рецептора фактора роста фибробластов-1 блокирует созревание висцеральной энтодермы и кавитацию в эмбриоидных телах мышей». Международный журнал биологии развития. 46 (6): 817–825. PMID 12382948.
- ^ Ван, Ю. Дж .; Wu, T. C .; Chung, A.E .; Дамжанов, И. (1984). «Моноклональные антитела к ламинину выявляют гетерогенность базальных мембран в тканях развивающихся и взрослых мышей». Журнал клеточной биологии. 98 (3): 971–979. Дои:10.1083 / jcb.98.3.971. ЧВК 2113154. PMID 6365932.
- ^ Li, X .; Chen, Y .; Schéele, S .; Arman, E .; Haffner-Krausz, R .; Экблом, П .; Лонай, П. (2001). «Передача сигналов фактора роста фибробластов и сборка базальной мембраны связаны во время эпителиального морфогенеза эмбриоидного тела». Журнал клеточной биологии. 153 (4): 811–822. Дои:10.1083 / jcb.153.4.811. ЧВК 2192393. PMID 11352941.
- ^ Coucouvanis, E .; Мартин, Г. Р. (1995). «Сигналы на смерть и выживание: двухступенчатый механизм кавитации в эмбрионе позвоночных». Клетка. 83 (2): 279–287. Дои:10.1016/0092-8674(95)90169-8. PMID 7585945.
- ^ Smyth, N .; Vatansever, H.S .; Murray, P .; Мейер, М .; Frie, C .; Paulsson, M .; Эдгар, Д. (1999). «Отсутствие базальных мембран после нацеливания на ген LAMC1 приводит к эмбриональной летальности из-за нарушения дифференцировки энтодермы». Журнал клеточной биологии. 144 (1): 151–160. Дои:10.1083 / jcb.144.1.151. ЧВК 2148127. PMID 9885251.
- ^ Murray, P .; Эдгар, Д. (2000). «Регулирование запрограммированной гибели клеток базальными мембранами в эмбриональном развитии». Журнал клеточной биологии. 150 (5): 1215–1221. Дои:10.1083 / jcb.150.5.1215. ЧВК 2175256. PMID 10974008.
- ^ Ying, Q. L .; Смит, А. Г. (2003). Определенные условия для нейронной приверженности и дифференциации. Методы в энзимологии. 365. С. 327–341. Дои:10.1016 / с0076-6879 (03) 65023-8. ISBN 9780121822682. PMID 14696356.
- ^ Wiles, M. V .; Келлер, Г. (1991). «Множественные гемопоэтические клоны развиваются из эмбриональных стволовых (ES) клеток в культуре». Разработка. 111 (2): 259–267. PMID 1893864.
- ^ Purpura, K. A .; Morin, J .; Зандстра, П. В. (2008). «Анализ временных и зависимых от концентрации эффектов BMP-4, VEGF и TPO на развитие мезодермы и клеток-предшественников эмбриональных стволовых клеток в определенной бессывороточной среде». Экспериментальная гематология. 36 (9): 1186–1198. Дои:10.1016 / j.exphem.2008.04.003. PMID 18550259.
- ^ Nostro, M.C .; Cheng, X .; Keller, G.M .; Гадью, П. (2008). «Передача сигналов Wnt, Activin и BMP регулирует отдельные этапы пути развития от эмбриональных стволовых клеток до крови». Стволовая клетка. 2 (1): 60–71. Дои:10.1016 / j.stem.2007.10.011. ЧВК 2533280. PMID 18371422.
- ^ Ten Berge, D .; Koole, W .; Fuerer, C .; Fish, M .; Eroglu, E .; Нуссе, Р. (2008). «Передача сигналов Wnt опосредует самоорганизацию и формирование оси в эмбриоидных телах». Стволовая клетка. 3 (5): 508–518. Дои:10.1016 / j.stem.2008.09.013. ЧВК 2683270. PMID 18983966.
- ^ Шукла, С .; Nair, R .; Rolle, M. W .; Браун, К. Р .; Chan, C.K .; Johnson, P. Y .; Wight, T. N .; Макдевитт, Т. К. (2009). «Синтез и организация гиалуронана и версикана эмбриональными стволовыми клетками, подвергающимися дифференцировке эмбриоидных тел». Журнал гистохимии и цитохимии. 58 (4): 345–358. Дои:10.1369 / jhc.2009.954826. ЧВК 2842597. PMID 20026669.
- ^ Bauwens, C.L .; Песня, H .; Thavandiran, N .; Унгрин, М .; Massé, S.P .; Nanthakumar, K .; Seguin, C .; Зандстра, П. В. (2011). «Геометрический контроль кардиомиогенной индукции в плюрипотентных стволовых клетках человека». Тканевая инженерия, часть А. 17 (15–16): 1901–1909. Дои:10.1089 / ten.TEA.2010.0563. PMID 21417693. S2CID 22010083.
- ^ Эйраку, М .; Takata, N .; Ishibashi, H .; Kawada, M .; Сакакура, Э .; Окуда, С .; Сэкигучи, К .; Адачи, Т .; Сасай, Ю. (2011). «Самоорганизующийся морфогенез глазного яблока в трехмерной культуре». Природа. 472 (7341): 51–56. Дои:10.1038 / природа09941. PMID 21475194.
- ^ Белинская, М .; Narita, N .; Уилсон, Д. Б. (1999). «Отличительные роли висцеральной энтодермы во время эмбрионального развития мыши». Международный журнал биологии развития. 43 (3): 183–205. PMID 10410899.
- ^ Burdsal, C.A .; Damsky, C.H .; Педерсен, Р. А. (1993). «Роль E-кадгерина и интегринов в дифференцировке мезодермы и миграции в примитивной полосе млекопитающих». Разработка. 118 (3): 829–844. PMID 7521282.
- ^ Finley, K. R .; Tennessen, J .; Шавлот, В. (2003). «Ген белка 5, секретируемого мышиной, экспрессируется в передней висцеральной энтодерме и энтодерме передней кишки во время раннего постимплантационного развития». Паттерны экспрессии генов. 3 (5): 681–684. Дои:10.1016 / s1567-133x (03) 00091-7. PMID 12972006.
- ^ Kemp, C .; Willems, E .; Abdo, S .; Ламбив, Л .; Лейнс, Л. (2005). «Экспрессия всех генов Wnt и их секретируемых антагонистов во время развития бластоцисты у мышей и после имплантации». Динамика развития. 233 (3): 1064–1075. Дои:10.1002 / dvdy.20408. PMID 15880404.
- ^ Rivera-Pérez, J.A .; Магнусон, Т. (2005). «Формированию примитивных полосок у мышей предшествует локальная активация Brachyury и Wnt3». Биология развития. 288 (2): 363–371. Дои:10.1016 / j.ydbio.2005.09.012. PMID 16289026.
- ^ а б Тернер, Дэвид; Алонсо-Крисостомо, Лус; Гиргин, Мехмет; Бэйли-Джонсон, Питер; Glodowski, Cherise R .; Хейворд, Пенелопа С .; Коллиньон, Жером; Густавсен, Карстен; Серуп, Палле (31.01.2017). «Гаструлоиды развивают три оси тела в отсутствие внеэмбриональных тканей и пространственно локализованной передачи сигналов». bioRxiv 10.1101/104539.
- ^ а б Тернер, Дэвид Эндрю; Glodowski, Cherise R .; Луз, Алонсо-Крисостомо; Бэйли-Джонсон, Питер; Hayward, Penny C .; Коллиньон, Жером; Густавсен, Карстен; Серуп, Палле; Шретер, Кристиан (13.05.2016). «Взаимодействия между Nodal и Wnt, сигнализирующими об устойчивом нарушении симметрии и осевой организации в гаструлоидах (эмбриональные органоиды)». bioRxiv 10.1101/051722.
- ^ а б Бэйли-Джонсон, Питер; Бринк, Сюзанна Карина ван ден; Балайо, Тина; Тернер, Дэвид Эндрю; Ариас, Альфонсо Мартинес (2015-11-24). "Создание агрегатов эмбриональных стволовых клеток мыши, которые демонстрируют нарушение симметрии, поляризацию и возникающее коллективное поведение. In vitro". Журнал визуализированных экспериментов (105). Дои:10.3791/53252. ISSN 1940-087X. ЧВК 4692741. PMID 26650833.
- ^ Бринк, Сюзанна К. ван ден; Бэйли-Джонсон, Питер; Балайо, Тина; Хаджантонакис, Анна-Катерина; Новочин, Соня; Тернер, Дэвид А .; Ариас, Альфонсо Мартинес (2014-11-15). «Нарушение симметрии, спецификация зародышевого листка и осевая организация в агрегатах эмбриональных стволовых клеток мыши». Разработка. 141 (22): 4231–4242. Дои:10.1242 / dev.113001. ISSN 0950-1991. ЧВК 4302915. PMID 25371360.
- ^ Тернер, Дэвид А .; Хейворд, Пенелопа С .; Бэйли-Джонсон, Питер; Руэ, По; Брум, Ребекка; Фавны, Фернандо; Ариас, Альфонсо Мартинес (2014-11-15). «Передача сигналов Wnt / β-катенина и FGF направляет спецификацию и поддержание нейромезодермального аксиального предшественника в ансамблях эмбриональных стволовых клеток мыши». Разработка. 141 (22): 4243–4253. Дои:10.1242 / dev.112979. ISSN 0950-1991. ЧВК 4302903. PMID 25371361.
- ^ а б Кинни, М. А .; Sargent, C. Y .; Макдевитт, Т. К. (2011). «Многопараметрические эффекты гидродинамической среды на культуру стволовых клеток». Тканевая инженерия, часть B: обзоры. 17 (4): 249–262. Дои:10.1089 / ten.TEB.2011.0040. ЧВК 3142632. PMID 21491967.
- ^ а б Карпенедо, Р. Л .; Bratt-Leal, A. S. M .; Marklein, R.A .; Seaman, S.A .; Bowen, N.J .; Макдональд, Дж. Ф .; Макдевитт, Т. К. (2009). «Гомогенная и организованная дифференциация внутри эмбриоидных тел, индуцированная микросферой доставки малых молекул». Биоматериалы. 30 (13): 2507–2515. Дои:10.1016 / j.biomaterials.2009.01.007. ЧВК 2921510. PMID 19162317.
- ^ Sachlos, E .; Огюст, Д. Т. (2008). «Морфология эмбрионального тела влияет на диффузионный транспорт индуктивных биохимических веществ: стратегия дифференцировки стволовых клеток». Биоматериалы. 29 (34): 4471–4480. Дои:10.1016 / j.biomaterials.2008.08.012. PMID 18793799.
- ^ Van Winkle, A. P .; Гейтс, И. Д .; Каллос, М. С. (2012). «Ограничения массопереноса в эмбриоидных телах во время дифференцировки человеческих эмбриональных стволовых клеток». Клетки Ткани Органы. 196 (1): 34–47. Дои:10.1159/000330691. PMID 22249133.
- ^ Bratt-Leal, A. S. M .; Карпенедо, Р. Л .; Ungrin, M.D .; Zandstra, P.W .; Макдевитт, Т. К. (2011). «Включение биоматериалов в многоклеточные агрегаты модулирует дифференцировку плюрипотентных стволовых клеток». Биоматериалы. 32 (1): 48–56. Дои:10.1016 / j.biomaterials.2010.08.113. ЧВК 2987521. PMID 20864164.
- ^ Purpura, K. A .; Bratt-Leal, A. S. M .; Хаммерсмит, К. А .; McDevitt, T. C .; Зандстра, П. В. (2012). «Систематическая инженерия трехмерных ниш плюрипотентных стволовых клеток для управления развитием крови». Биоматериалы. 33 (5): 1271–1280. Дои:10.1016 / j.biomaterials.2011.10.051. ЧВК 4280365. PMID 22079776.
- ^ Bratt-Leal, A. S. M .; Kepple, K. L .; Карпенедо, Р. Л .; Cooke, M.T .; Макдевитт, Т. К. (2011). «Магнитная манипуляция и формирование пространственного паттерна агрегатов многоклеточных стволовых клеток». Интегративная биология. 3 (12): 1224–1232. Дои:10.1039 / c1ib00064k. ЧВК 4633527. PMID 22076329.
- ^ Akins, R.E .; Rockwood, D .; Робинсон, К. Г .; Сандаски, Д .; Rabolt, J .; Писарро, К. (2010). «Трехмерная культура изменяет первичный фенотип сердечных клеток». Тканевая инженерия, часть А. 16 (2): 629–641. Дои:10.1089 / ten.tea.2009.0458. ЧВК 2813151. PMID 20001738.
- ^ Chang, T. T .; Хьюз-Фулфорд, М. (2009). «Монослойная и сфероидная культура клеток линии клеток гепатоцеллюлярной карциномы печени человека демонстрирует отчетливые глобальные паттерны экспрессии генов и функциональные фенотипы». Тканевая инженерия, часть А. 15 (3): 559–567. Дои:10.1089 / ten.tea.2007.0434. ЧВК 6468949. PMID 18724832.