Гибридогенез у водных лягушек - Hybridogenesis in water frogs
В плодородный гибриды из Европейские водяные лягушки (род Пелофилакс) воспроизводить гибридогенез (гемиклонально ). Это означает, что во время гаметогенез, они отбрасывают геном одного из родительских видов и производят гаметы другого родительского вида (содержащего геном не рекомбинированный с геномом первого родительского вида).[1][2][3][4] Первый родительский геном восстанавливается оплодотворение этих гамет с гаметами первого вида (сексуальный хозяин ).[5][1][4] В полностью гибридных популяциях съедобная лягушка Пелофилакс кл. Esculentus, тем не мение, триплоид гибриды обеспечивают этот недостающий геном.[3][6][2]
Потому что половина генома передается следующему поколению клонально (не исключен нерекомбинированный интактный геном), и только другая половина половым путем (рекомбинированный геном полового хозяина) гибридогенез представляет собой гемиклональный способ размножения.[7][8][4]
Например, съедобная лягушка Пелофилакс кл. Esculentus (в основном геном RL), который является гибридогенетическим гибридом болотная лягушка P. ridibundus (RR) и лягушка P. lessonae (LL), обычно исключает Lessonae геном (L) и генерирует гаметы P. ridibundus (Р). Другими словами, съедобные лягушки производят гаметы болотных лягушек.[2][3][4]
Однако гибридные популяции размножаются не указанными выше первичными гибридизациями, а преимущественно обратные кресты с одним из родительских видов они сосуществуют (живут в сочувствие[9][10]) с помощью (см. ниже посередине). [11][2][3][12][9][4]
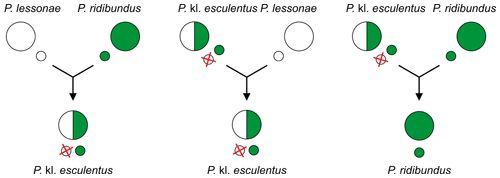
Поскольку гибридогенетическим гибридам требуется другой таксон в качестве полового хозяина для воспроизводства, обычно один из родительских видов, их называют клептоны[13][14][5] (с «кл.» в научных названиях[15]).

Известны три гибридогенетических гибрида Европейский водяные лягушки:
- съедобная лягушка Пелофилакс кл. Esculentus (обычно генотип RL):
лягушка P. lessonae (LL) × P. ridibundus (RR)[11][2] - Гибридная лягушка Графа Пелофилакс кл. графи (PR):
Лягушка Переса П. Перези (ПП) × P. ridibundus (RR) или
Лягушка Переса П. Перези (ПП) × съедобная лягушка П. кл. Esculentus (RE)
(неясно, какой из скрещиваний был первичной гибридизацией)[2] - Итальянская съедобная лягушка Пелофилакс кл. испанец (РБ):
Итальянская лягушка в бассейне П. бергери (BB) × P. ridibundus (RR)[2]
Исключение родительского генома
Гибридогенез подразумевает, что гаметы гибриды не содержат смешанных родителей геномы, как это обычно происходит в результате независимой сегрегации хромосом и кроссовер в мейоз (смотрите также второй закон Менделя, рекомбинация ). Вместо этого каждая гамета несет полную (гаплоидный ) геном только одного родительского вида. Обычно один полный геном родительского вида исключается до мейоза во время гаметогенез, так что среди гамет представлен только один (оставшийся) родительский геном, а гены от другого родителя не передаются гибридогеном.[16][3][2] Этот сброс происходит постепенно во время последующих митотический деления, а не за один шаг.[2]

 Типичный гаметогенез в Пелофилакс кл. Esculentus (в системе L-E). 1 - исключение P. lessonae геном, 2 - дупликация (эндоредупликация ) из P. ridibundus геном - восстановление диплоидия, 3 - мейоз, L и R - P. lessonae и ридибундус геномы. [16] | 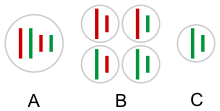 Гаметы гибридогенетической гибридный содержать несоединенных геном одного родительского вида (C) вместо всех возможных комбинаций обоих родительских (красный и зеленый) хромосомы (В). А - соматический клетка. |
Гемиклоны
Гибридогенез - это гемиклональный способ размножения - половина гибридный геном передается нетронутым клонально из поколения в поколение (р геном в системе L-E) - не рекомбинирован с геномом родительского вида (здесь L), тогда как другая половина (L) передается сексуально - получено (заменено) каждое поколение путем полового размножения родительским видом (сексуальный хозяин[5][1][4], P. lessonae в системе L-E).[7][8][4]
Обзор гибридогенетических систем
Существует как минимум три гибридогенетических вида (гибриды ) водяных лягушек в Европа – съедобная лягушка Пелофилакс кл. Esculentus, Гибридная лягушка Графа Пелофилакс кл. графи и Итальянская съедобная лягушка Пелофилакс кл. испанец. Их модели спаривания подразделяются на несколько гибридогенетических систем:[2]
| Гибридный | Возник из | Поддерживается скрещивается с | не входит геном | Система | ||
|---|---|---|---|---|---|---|
| Пелофилакс кл. Esculentus RL | P. ridibundus RR | × | P. lessonae LL | P. lessonae LL | L | L – E |
| P. ridibundus RR | R или L 3: 1 | R – E | ||||
| П. кл. Esculentus LLR | L из RL R из LLR | E | ||||
| П. кл. Esculentus RRL | R или L 3: 1 от RL L из РРЛ | E | ||||
| Пелофилакс кл. графи RP | P. ridibundus ? RR или П. кл. Esculentus ? RL | × | П. Перези PP | П. Перези PP | п | P – G |
| Пелофилакс кл. испанец РБ | P. ridibundus RR | × | П. бергери BB | П. бергери BB | B | B – H |
(заглавные аббревиатуры под научными названиями генотипы )
Все эти гибриды содержат геном из болотная лягушка P. ridibundus (R) и геном второго родительского вида (L, P или B).[2]
Большинство вышеперечисленных гибридогенных систем представляют собой сосуществующие гибридные (живущие в сочувствие[9][10]) с одним из родительских видов, необходимых для его воспроизводства.[2] П. кл. Esculentus например, в наиболее часто встречающейся системе L-E должен спариваться с P. lessonae для производства новых гибридов в системе R-E с P. ridibundus.[3][4] Поскольку эти гибриды зависят от других таксонов как сексуальный хозяева для размножения ("паразитировать" на них половым путем) они клептоны[13][14][5] ("кл." в научных названиях[15]).
Съедобная лягушка Пелофилакс кл. Esculentus
В Pelophylax esculentus комплекс состоит из гибридный таксон - съедобная лягушка П. кл. Esculentus (генотип RL) и родительские виды - болотная лягушка P. ridibundus (RR) и лягушка P. lessonae (LL). Гибриды бывают самки и самцы, что необычно, потому что гибриды других гибридогенных видов - только самки.[2]
Происхождение первичной гибридизации П. кл. Esculentus (генотип RL) это:
- P. lessonae (LL) × P. ridibundus (RR)
Это происходит между P. lessonae (LL) мужчины и P. ridibundus (RR) самки[11][2][3][9][4], потому что меньше P. lessonae самцы предпочитают более крупных самок.[11][2][3][4] Родословные гибридов поддерживаются позже посредством других вязок, описанных ниже.[2][3][6]
P. lessonae и P. ridibundus имеют различные требования к среде обитания и обычно не живут вместе.[17][18]
P. lessonae – П. кл. Esculentus (L – E) система
В P. lessonae – П. кл. Esculentus[2] (L – E[2][4][9][12][8], LE[3][6], lessonae – esculentus[3]) система является наиболее распространенной гибридогенетической системой.[2][4] Встречается в западных Европа.[2]
Гибриды П. кл. Esculentus (генотип RL) исключить здесь P. lessonae геном (L) и делать исключительно клональный P. ridibundus гаметы (R).[2][4] Другими словами, съедобные лягушки производят гаметы болотных лягушек![4] Их родословная обычно сохраняется через обратные кресты женщины П. кл. Esculentus (RL) с мужчиной P. lessonae (LL). Потомство состоит только из П. кл. Esculentus.[2][3]
П. кл. Esculentus гибриды (RL) могут также спариваться друг с другом, но только 3% полученных головастиков (RR) доживают до половой зрелости (97% - нет). Геномы межгибридных скрещиваний женские, из-за несения Х-хромосомы самок от первичной гибридизации.[2]

P. ridibundus – П. кл. Esculentus (R – E) система
В P. ridibundus – П. кл. Esculentus[2] (R – E[2][4], RE[3][6], ridibundus – esculentus[3]) система обитает на востоке Европа.[2]
По сути, это обратная форма системы L – E.[2][3]
Гибриды П. кл. Esculentus (генотип RL) исключить здесь P. ridibundus (R) или P. lessonae (L) геном в соотношении 3: 1 и в основном клональный P. lessonae (L), меньше P. ridibundus гаметы (R).[2] Одна лягушка производит либо L, либо R гаметы, либо их смесь.[4] Их родословная сохраняется через обратные кресты мужчины[3] П. кл. Esculentus (RL) с самкой[3] P. ridibundus (RR).[2][3] Потомство состоит из П. кл. Esculentus мужчины (75%) или P. ridibundus самки (25%). Это называется гибридной амфиспермией.[2]
Полностью гибридные популяции (система E)
Все-гибридный население[3][2] (Система E[2], EE – system[6]) состоят исключительно из П. кл. Esculentus – диплоид RE и триплоид Гибриды LLR или RRL.[3][2] Есть даже известные тетраплоид LLRR гибриды.[3] Полногибридные популяции населяют весь ареал комплекса водных лягушек.[3]
RL диплоиды отбросить L геном и производят гаметы P. ridibundus (R), или отказаться от генома R или L и произвести гаметы P. lessonae (L) или P. ridibundus (R) соответственно. В обоих случаях диплоидные гибриды генерируют также не восстановленные диплоидные гаметы (RL), необходимые для воссоздания триплоидов.[2]
Триплоиды LLR и RRL являются поставщиками P. lessonae (L)[2] и P. ridibundus гаметы (R) соответственно в этой системе, лишенные обоих родительских видов.[2] Так триплоидные гибриды позволяют П. кл. Esculentus популяции остаться без родительского вида.[3]
Поскольку триплоиды отбрасывают этот геном, который доступен в одной копии, и оставляют две копии второго генома, они не выполняют эндоредупликацию.[2] Более того, этот неэлиминированный геном передается гаплоидным гаметам. сексуально, нет клонально (рекомбинированный между двумя L или между двумя R), в отличие от генома диплоидных гибридов.[3][6]
Такой модифицированный гибридогенез[19] (или же гаметогенетический система[20]) происходящие в аллотриплоид гибриды, где во время мейоз хромосомы (геномы) из удвоенного набора (здесь LL из LLR или RR из RRL) используются для производства гаплоидных гамет (L или R соответственно), тогда как остальные могут быть исключены (R из LLR или L из RRL), известный как мейотический гибридогенез.[19][20][6]
В одной Словацкий Однако в популяции триплоидные самцы (LLR) и диплоидные самки LR генерируют клональные LL и клональные R гаметы, соответственно, вместо рекомбинированных L и клональных LR.[6]
P. lessonae (LL) и P. ridibundus (RR) потомство не доживает до половой зрелости в системе E.[2][3]
| ||||||||||||||||||||||||||||||||||||
| Поддержание чистого (полностью гибридного) П. кл. Esculentus населения, без P. lessonae и ридибундус.[3] L, р – P. lessonae и P. ridibundus гаплоидный геномы; | ||||||||||||||||||||||||||||||||||||

Гибридная лягушка Графа Пелофилакс кл. графи и система P – G

Неясно, возникла ли первичная гибридизация Гибридная лягушка Графа Пелофилакс кл. графи (генотип PR) был:[2]
- Лягушка Переса П. Перези (ПП) × болотная лягушка P. ridibundus (RR) или
- Лягушка Переса П. Перези (ПП) × съедобная лягушка П. кл. Esculentus (RE)
В отличие от П. Перези и Пелофилакс кл. графи, P. ridibundus и П. кл. Esculentus не принадлежат к местной фауне Пиренейский полуостров.[2]
Гибриды П. кл. графи (PR) выбросить П. Перези геном (P) и производить исключительно клональный P. ridibundus гаметы (R). Их родословная поддерживается в так называемой системе P – G через обратные кресты из П. кл. графи (PR) с П. Перези (ПП).[2]
Итальянская съедобная лягушка Пелофилакс кл. испанец и система B – H

Первичная гибридизация, которая возникла Итальянская съедобная лягушка Пелофилакс кл. испанец (генотип RB) был:[2]
- Итальянская лягушка в бассейне П. бергери (BB) × болотная лягушка P. ridibundus (RR)
Гибриды Пелофилакс кл. испанец (RB) отбросить П. бергери геном (B) и производить исключительно клональный P. ridibundus гаметы (R). Их родословная поддерживается в так называемой системе B – H через обратные кресты из П. кл. испанец (PR) с П. бергери (BB).[2]
Водяные лягушки и определение гибридогенеза
Способы матирования гибридогенетических водных лягушек не соответствуют точно известным определениям гибридогенеза:[21][1][7]
- гибридогенетический гибриды не только женщины[21][1][7], но и самцы[2][3][4]
- в полностью гибридных популяциях съедобная лягушка Пелофилакс кл. не другой вид,[21][1] но Esculentus триплоид гибриды одного вида обеспечивают исключенный геном,[3][6][2] служение (принятие роли[22]) в качестве полового хозяина (виды полового хозяина из определений[1]),[22][23] что на самом деле не является исключением из правил.
Митохондриальная ДНК
В Пелофилакс кл. Esculentus сложные лягушки имеют один из четырех фенотипов мтДНК:[9]
| Таксон | тип мтДНК | |||
|---|---|---|---|---|
| А | B | C | D | |
| болотная лягушка P. ridibundus | + | + | ||
| лягушка P. lessonae | + | + | ||
| съедобная лягушка П. кл. Esculentus | + | + | + | + |
Тип А - это P. ridibundus специфичен, а тип B P. lessonae-подобно[2] (отличается только на 0,3% от типа С[9]). Большинство П. кл. Esculentus имеют фенотип C или D P. lessonae, нет P. ridibundus мтДНК.[9][2]
Распределение этих фенотипов не совсем точно отражает типичные модели матирования. Митохондрии вместе с мтДНК наследуются исключительно от самки. Поскольку первичные гибридизации производят П. кл. Esculentus происходить между P. ridibundus самки (большие) и P. lessonae самцы (маленькие) и позже сохраняются через обратные кресты П. кл. Esculentus женщины с P. lessonae мужчины (система L – E[2]), ожидаемый фенотип мтДНК П. кл. Esculentus будет фенотипом P. ridibundus. Это неожиданное распределение фенотипов можно объяснить таким образом, что большая часть П. кл. Esculentus линии могут пройти по крайней мере одно обратное скрещивание между П. кл. Esculentus мужчина с P. lessonae женский.[9][2] И такая картина фенотипа говорит о том, что первичные гибридизации редки.[9]
Интрогрессия P. lessonae мтДНК в P. ridibundus (тип B[9]) может быть вызвано матированием между P. ridibundus и П. кл. Esculentus имея P. lessonae мтДНК.[2]
Эволюционное происхождение гибридогенеза у съедобной лягушки
Вовремя ледниковые периоды,[требуется разъяснение ] популяция общего предка обоих родительских видов съедобная лягушка был разделен на две части. Эти популяции расходились, но оставались генетически достаточно близкими, чтобы иметь возможность создавать плодородные гибриды. Однако, когда диплоидные съедобные лягушки спариваются друг с другом, их потомство часто искажается, поэтому нет чистых популяций съедобных лягушек, если не присутствуют некоторые триплоидные особи (система E, описанная выше).
Воздействие чужеродных видов
Вступление чужеродных видов, относящихся к комплексу водяных лягушек (Pelophylax esculentus сложный), например, экзотический болотная лягушка P. ridibundus, может быть вредным для местных популяций лягушек из-за создания новых возможностей гибридизации и последующего исключения некоторых из геномы от населения. В некоторых случаях это было доказано.[2][8][24]
Смотрите также
Рекомендации
- ^ а б c d е ж грамм Вриенхук, Роберт К. (1998). «Партеногенез и природные клоны» (PDF). В Кнобиле, Эрнст; Нил, Джимми Д. (ред.). Энциклопедия репродукции. 3. Академическая пресса. стр.695–702. ISBN 978-0-12-227020-8.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление ае аф аг ах ай эй ак аль являюсь ан ао ap водный ар в качестве в au средний ау топор ай az ба bb до н.э Holsbeek, G .; Джорис, Р. (2010). «Возможное влияние исключения генома чужеродными видами на гибридогенетических водных лягушек (Pelophylax esculentus сложный)" (PDF). Биологические вторжения. 12: 1–13. Дои:10.1007 / s10530-009-9427-2. ISSN 1387-3547. Получено 2015-06-21.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab ac объявление Кристиансен, Д. Г. (2009). "Типы гамет, определение пола и стабильное равновесие общегибридных популяций диплоидных и триплоидных съедобных лягушек (Pelophylax esculentus) Рана Эскулента как установлено на основе анализа мтДНК ". BMC Эволюционная биология. 9 (135): 135. Дои:10.1186/1471-2148-9-135. ЧВК 2709657. PMID 19527499.
- ^ а б c d е ж грамм час я j k л м п о п q р s Раггианти, М; Bucci, S; Марраччи, S; Casola, C; Mancino, G; Hotz, H; Guex, GD; Plötner, J; Уззелл, Т. (февраль 2007 г.). «Гаметогенез межгрупповых гибридов гемиклональных лягушек». Генетические исследования. 89 (1): 39–45. Дои:10.1017 / S0016672307008610. PMID 17517158. Получено 2015-06-21.
- ^ а б c d Опросы Пелаз, Мануэль (октябрь 1990 г.). «Концепция биологического клептона (BKC)». Алитес. 8 (3): 75–89. Получено 2015-06-22.
- ^ а б c d е ж грамм час я Pruvost, Nicolas B.M; Хоффманн, Александра; Рейер, Хайнц-Ульрих (сентябрь 2013 г.). "Образцы производства гамет, плоидность и популяционная генетика выявляют эволюционно значимые единицы у гибридных водных лягушек (Pelophylax esculentus)". Экология и эволюция. 3 (9): 2933–2946. Дои:10.1002 / ece3.687. ЧВК 3790541. PMID 24101984.
- ^ а б c d Simon, J.-C .; Delmotte, F .; Rispe, C .; Crease, T. (2003). «Филогенетические отношения между партеногенами и их половыми родственниками: возможные пути к партеногенезу у животных» (PDF). Биологический журнал Линнеевского общества. 79: 151–163. Дои:10.1046 / j.1095-8312.2003.00175.x. Получено 2015-06-21.
- ^ а б c d е ж Форбургер, Кристоф; Рейер, Хайнц-Ульрих (2003). "Генетический механизм смены видов у европейских лягушек?" (PDF). Сохранение генетики. 4 (2): 141–155. Дои:10.1023 / А: 1023346824722. ISSN 1566-0621. Получено 2015-06-21.
- ^ а б c d е ж грамм час я j k л Спольски, К; Уззелл, Т. (1986). «Эволюционная история гибридогенетической гибридной лягушки. Рана Эскулента как установлено на основе анализа мтДНК ". Молекулярная биология и эволюция. 3 (1): 44–56. Дои:10.1093 / oxfordjournals.molbev.a040376. PMID 2832687. Получено 2015-06-21.
- ^ а б Christiansen, Ditte G .; Рейер, Хайнц-Ульрих (июль 2009 г.). «От клональных гибридов к половым: генетическая рекомбинация с помощью триплоидов в полностью гибридных популяциях водных лягушек» (PDF). Эволюция. 63 (7): 1754–1768. Дои:10.1111 / j.1558-5646.2009.00673.x. PMID 19245393. Получено 2015-06-21.
- ^ а б c d е Бергер, Л. (1970). «Некоторые характеристики крестов внутри Рана Эскулента комплекс в постличиночном развитии ». Annales Zoologici, Варшава. 27: 374–416.
- ^ а б c Абт Титже, Габи; Рейер, Хайнц-Ульрих (2004). «Развитие личинок и пополнение молоди в естественной популяции Rana Lessonae и Рана Эскулента" (PDF). Копея. 2004 (3): 638–646. Дои:10.1643 / ce-03-273r1. Получено 2015-06-21.
- ^ а б Дюбуа, Ален (2009). «Бесполые и метасексуальные позвоночные. Рецензия на книгу». Алитес. 27 (2): 62–66. Получено 2015-06-22.
Джон С. Авис, 2008. – Клональность. Генетика, экология и эволюция полового воздержания у позвоночных животных. Нью-Йорк, Oxford University Press: i – xi + 1–237. ISBN 978-0-19-536967-0.
- ^ а б Дюбуа, А .; Гюнтер, Р. (1982). «Клептон и синклептон: две новые категории эволюционной систематики в зоологии». Zoologische Jahrbücher. Abteilung für Systematik, Geographie und Biologie der Tiere. 109: 290–305. ISSN 0044-5193.
- ^ а б Дюбуа, Ален (октябрь 1990). «Номенклатура партеногенетических, гиногенетических и гибридогенетических таксонов позвоночных: новые предложения». Алитес. 8 (3): 61–74. Получено 2015-06-22.
- ^ а б Tunner, H.G .; Хеппич-Таннер, С. (1991). «Исключение генома и две стратегии дупликации хромосом в оогенезе гибридной лягушки». Naturwissenschaften. 78 (1): 32–34. Дои:10.1007 / BF01134041.
- ^ Бергер, Л. (1982). «Зимняя спячка европейских водяных лягушек (Рана Эскулента сложный)". Zoologica Poloniae. 29: 57–72.
- ^ Холенвег Петер, Анна-Катерина (декабрь 2001 г.). "Скорости рассеяния и расстояния у взрослых водных лягушек, Rana Lessonae, Р. ридибунда и их гибридогенетический партнер R. esculenta". Herpetologica. 57 (4): 449–460. JSTOR 3893052. url2
- ^ а б Алвес, М. Юдит; Коэльо, М. Мануэла; Колларес-Перейра, М. Жоао (1998). "Разнообразие репродуктивных форм самок Rutilus alburnoides комплекс (Teleostei, Cyprinidae): способ избежать генетических ограничений униродительства " (PDF). Молекулярная биология и эволюция. 15 (10): 1233–1242. Дои:10.1093 / oxfordjournals.molbev.a025852. ISSN 0737-4038. Получено 2015-06-27.
- ^ а б Моришима, К .; Yoshikawa, H .; Араи, К. (2008). «Мейотический гибридогенез в триплоидных Мисгурнус вьюн происходит от клональной линии ". Наследственность. 100 (6): 581–586. Дои:10.1038 / hdy.2008.17. PMID 18382473.
- ^ а б c Шульц, Р. Джек (ноябрь – декабрь 1969 г.). "Гибридизация, однополость и полиплоидия костистых зубов Poeciliopsis (Poeciliidae) и других позвоночных ". Американский натуралист. 103 (934): 605–619. Дои:10.1086/282629. JSTOR 2459036.
- ^ а б Хоффманн, Александра; Рейер, Хайнц-Ульрих (4 декабря 2013 г.). «Влияние генома на структуру рекламного вызова у диплоидных и триплоидных гибридных водяных лягушек (Anura, Pelophylax esculentus)". BMC Ecology. 13 (47): 47. Дои:10.1186/1472-6785-13-47. ЧВК 4235041. PMID 24304922.
- ^ Arioli, M .; Jakob, C .; Рейер, Х.У. (Май 2010 г.). «Генетическое разнообразие гибридов водяной лягушки (Pelophylax esculentus) зависит от структуры населения и географического положения ". Молекулярная экология. 19 (9): 1814–28. Дои:10.1111 / j.1365-294X.2010.04603.x. PMID 20374490.
- ^ Quilodrán, Claudio S .; Монтойя-Бургос, Хуан I .; Куррат, Матиас (2015). "Моделирование межвидовой гибридизации с исключением генома для определения действий по сохранению: случай нативной и инвазивной Пелофилакс водяные лягушки " (PDF). Эволюционные приложения. 8 (2): 199–210. Дои:10.1111 / eva.12245. ЧВК 4319866. PMID 25685194. Архивировано из оригинал (PDF) на 2015-06-22. Получено 2015-06-21.