Фотопериодизм - Photoperiodism
Фотопериодизм это физиологическая реакция организмов на продолжительность ночи или темный период. Это происходит в растения и животные. Фотопериодизм также можно определить как реакцию развития растений на относительную продолжительность светового и темного периодов. В соответствии с фотопериодами они подразделяются на три группы: растения короткого дня, растения длинного дня и растения нейтрального дня.
Растения
Много цветущие растения (покрытосеменные) используют фоторецепторный белок, Такие как фитохром или же криптохром,[1] чтобы почувствовать сезонные изменения продолжительности ночи или фотопериода, которые они воспринимают как сигнал к цветению. В следующем подразделе обязать фотопериодическим растениям абсолютно необходима длинная или короткая ночь перед цветением, тогда как факультативный фотопериодические растения с большей вероятностью зацветут при одном условии.
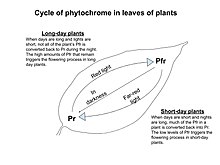
Фитохром бывает двух видов: Pр и Pfr. Красный свет (который присутствует в течение дня) превращает фитохром в его активную форму (pfr). Затем это заставляет растение расти. В свою очередь, дальний красный свет присутствует в тени или в темноте, и это превращает фитохром из pfr в pr. пр является неактивной формой фитохрома и не позволяет растениям расти. Эта система Pfr верхр преобразование позволяет растению чувствовать, когда сегодня ночь, а когда день.[2] пfr также можно преобразовать обратно в Pр с помощью процесса, известного как темная реверсия, когда длительные периоды темноты запускают преобразование Pfr.[3] Это важно для цветения растений. Эксперименты Халлидея и др. показали, что манипуляции с соотношением красного к дальнему красному у Arabidopsis могут изменить цветение. Они обнаружили, что растения, как правило, зацветают позже, когда подвергаются большему количеству красного света, доказывая, что красный свет тормозит цветение.[4] Другие эксперименты доказали это, подвергая растения дополнительному воздействию красного света посреди ночи. Растение короткого дня не зацветет, если свет включен на несколько минут посреди ночи, а растение длинного дня может зацвести, если подвергнуться большему количеству красного света посреди ночи.[5]
Криптохромы - это еще один тип фоторецепторов, который важен при фотопериодизме. Криптохромы поглощают синий свет и УФ-А. Криптохромы заставляют светиться циркадные часы.[6] Было обнаружено, что изобилие как криптохрома, так и фитохрома зависит от света, а количество криптохрома может меняться в зависимости от продолжительности дня. Это показывает, насколько важны оба фоторецептора для определения длины дня.[7]
В 1920 году У. Гарнер и Х. А. Аллард опубликовали свои открытия по фотопериодизму и сочли, что решающее значение имеет продолжительность светового дня.[1][8] но позже было обнаружено, что решающим фактором была продолжительность ночи.[9][10] Фотопериодические цветущие растения классифицируются как растения длинного дня или же растения короткого дня хотя ночь является решающим фактором из-за первоначального неправильного понимания того, что дневной свет является решающим фактором. Наряду с растениями длинного дня и растениями короткого дня, есть растения, которые попадают в «категорию длины двойного дня». Это растения либо растения длинного короткого дня (LSDP), либо растения короткого длинного дня (SLDP). LSDP цветут после серии длинных дней, за которыми следуют короткие дни, тогда как SLDP цветут после серии коротких дней, за которыми следуют длинные.[11] У каждого растения разная длина критического фотопериода или критическая продолжительность ночи.[1]
Современные биологи считают[12] что это совпадение активных форм фитохрома или криптохрома, создаваемых светом в дневное время, с ритмами циркадные часы что позволяет растениям измерять продолжительность ночи. Помимо цветения, фотопериодизм у растений включает рост стеблей или корней в определенные сезоны и потерю листьев. Искусственное освещение можно использовать для увеличения продолжительности рабочего дня.[1]
Многолетние растения
Растения с длинным днем цветут, когда длина ночи опускается ниже критического светового периода.[13] Эти растения обычно цветут в конце весны или в начале лета, когда дни становятся длиннее. В северном полушарии самый длинный день в году (летнее солнцестояние) приходится на 21 июня или около того.[14] После этой даты дни становятся короче (то есть ночи становятся длиннее) до 21 декабря (зимнего солнцестояния). В южном полушарии ситуация обратная (т.е. самый длинный день - 21 декабря, а самый короткий - 21 июня).[1][8]
Некоторые облигатные растения длинного дня:
Некоторые факультативные растения длинного дня:
Растения короткого дня
Короткодневные растения цветут, когда продолжительность ночи превышает их критический световой период.[15] Они не могут цвести в короткие ночи или если на растение в течение нескольких минут попадает импульс искусственного света в течение ночи; им требуется постоянный период темноты, прежде чем начнется развитие цветков. Естественный ночной свет, такой как лунный свет или молния, не обладает достаточной яркостью или продолжительностью, чтобы прервать цветение.[1][8]
Как правило, растения с коротким днем (т. Е. С длинными ночами) цветут, когда дни становятся короче (а ночи становятся длиннее) после 21 июня в северном полушарии, то есть летом или осенью. Продолжительность темного периода, необходимого для цветения, различается для разных видов и разновидностей вида.
Фотопериодизм влияет на цветение, заставляя побеги производить цветочные почки вместо листьев и боковых бутонов.
Некоторые факультативные растения короткого дня:[16]
- Кенаф ( Гибискус каннабинус)
- Марихуана (Каннабис)
- Хлопок (Госсипиум)
- Рис (Орыза)
- Джовар (Сорго двухцветное)
- Зеленый грамм (Фасоль мунг, Vigna radiata)
- Соевые бобы[17] (Глицин макс)
Дневно-нейтральные растения
Дневно-нейтральные растения, такие как огурцы, розы, помидоры, и Рудералис (автоцветущий каннабис ) не инициируют цветение на основании фотопериодизма.[18] Вместо этого они могут начать цветение после достижения определенной общей стадии развития или возраста или в ответ на альтернативные раздражители окружающей среды, такие как яровизация (период низкой температуры).[1][8]
Животное
Продолжительность светового дня и, следовательно, знание времени года жизненно важны для многих животных. От этого знания зависит ряд биологических и поведенческих изменений. Фотопериод вместе с перепадами температуры провоцирует изменение цвета меха и перьев, миграцию, попадание в спячка, сексуальное поведение и даже изменение размеров половых органов.
Частота пения птиц, таких как канарейка зависит от фотопериода. Весной, когда увеличивается световой период (больше дневного света), семенники у самца канарейки растут. По мере роста яичек секретируется больше андрогенов, и частота пения увеличивается. Осенью, когда световой период уменьшается (меньше дневного света), семенники самцов канарейки регрессируют, и уровень андрогенов резко падает, что приводит к снижению частоты пения. От фотопериода зависит не только частота пения, но и репертуар песни. Продолжительный световой период весны приводит к расширению песенного репертуара. Более короткий световой период осени приводит к сокращению песенного репертуара. Эти поведенческие изменения фотопериода у самцов канареек вызваны изменениями песенного центра мозга. По мере увеличения фотопериода высокий вокальный центр (HVC) и надежное ядро архистриатум (RA) увеличиваются в размерах. Когда фотопериод уменьшается, эти области мозга регрессируют.[19]
У млекопитающих продолжительность светового дня регистрируется в супрахиазматическое ядро (SCN), о чем сообщает светочувствительные ганглиозные клетки сетчатки, которые не участвуют в зрении. Информация проходит через ретиногипоталамический тракт (RHT). Некоторые млекопитающие очень сезонны, в то время как сезонность людей считается эволюционный багаж.[20][соответствующий? ]
Смотрите также
Рекомендации
- ^ а б c d е ж грамм Маузет, Джеймс Д. (2003). Ботаника: введение в биологию растений (3-е изд.). Садбери, Массачусетс: Джонс и Бартлетт Обучение. С. 422–27. ISBN 978-0-7637-2134-3.
- ^ Фанкхаузер, Кристиан (2001). «Фитохромы, семейство красных / дальних красных фоторецепторов». Журнал биологической химии. 276 (15): 11453–11456. Дои:10.1074 / jbc.R100006200. ISSN 0021-9258. PMID 11279228.
- ^ Casal, J.J. (2014). «Восприятие света и сигнализация фитохромом А». Журнал экспериментальной ботаники. 65 (11). (11): 2835–2845. Дои:10.1093 / jxb / ert379. PMID 24220656.
- ^ Линь, Чентао (2000). «Фоторецепторы и регуляция времени цветения». Физиология растений. 123 (1): 39–50. Дои:10.1104 / стр.123.1.39. ЧВК 1539253. PMID 10806223.
- ^ Чамовиц, Даниэль (2013). Что знает растение. Scientific American. С. 17–18. ISBN 978-0-374-28873-0.
- ^ Линь, Чентао (2005). «Криптохромы». Геномная биология. 6 (5): 220. Дои:10.1186 / gb-2005-6-5-220. ЧВК 1175950. PMID 15892880.
- ^ Моклер, Тодд (2003). «Регулирование фотопериодического цветения фоторецепторами Arabidopsis». Труды Национальной академии наук Соединенных Штатов Америки. 100 (4): 2140–2145. Bibcode:2003ПНАС..100.2140М. Дои:10.1073 / pnas.0437826100. ЧВК 149972. PMID 12578985.
- ^ а б c d Каплон, Брайан (2005). Ботаника для садоводов (2-е изд.). Портленд, штат Орегон: Timber Publishing. С. 148–51. ISBN 978-0-88192-655-2.
- ^ Hamner, K.C .; Боннер, Дж. (1938). «Фотопериодизм в отношении гормонов как факторов зарождения и развития цветков» (PDF). Ботанический вестник. 100 (2): 388–431. Дои:10.1086/334793. JSTOR 2471641.
- ^ Хамнер, К. (1940). «Взаимосвязь света и тьмы в фотопериодической индукции». Ботанический вестник. 101 (3): 658–87. Дои:10.1086/334903. JSTOR 2472399.
- ^ Тайз, Линкольн; Зейгер, Эдуардо; Мёллер, Ян; Мерфи, Ангус (2015). Физиология и развитие растений (Шестое изд.). Сандерленд, Массачусетс: Sinauer Associates, Inc. ISBN 978-1-60535-353-1.
- ^ Андрес, Фернандо; Гэлбрейт, Дэвид В .; Талон, Мануэль; Доминго, Конча (12 августа 2009 г.). «Анализ ЧУВСТВИТЕЛЬНОСТИ ФОТОПЕРИОДА5 проливает свет на роль фитохромов в фотопериодическом цветении риса». Физиология растений. 151 (2): 681–690. Дои:10.1104 / pp.109.139097. ЧВК 2754645. PMID 19675157.
- ^ Старр, Сеси; Таггарт, Ральф; Эверс, Кристина; Старр, Лиза (2013). Структура и функции завода. 4 (13-е изд.). Брукс / Коул. п. 517. ISBN 978-1-111-58068-1.
- ^ Гули, Тристан (30 марта 2010 г.). Естественный навигатор. Случайный дом. ISBN 978-0-7535-2311-7.
- ^ Биология BSCS (9-е изд.). BSCS. 2002. с. 519. ISBN 978-0-7872-9008-5.
- ^ Джонс, Хэмлин Г. (1992). Растения и микроклимат: количественный подход к экологической физиологии растений. Издательство Кембриджского университета. п.225. ISBN 978-0-521-42524-7.
- ^ Перселл, Ларри С.; Салмерон, Монтсеррат; Эшлок, Лэнни (2014). "Глава 2" (PDF). Справочник по производству сои в Арканзасе - MP197. Литл-Рок, Арканзас: Кооперативная служба распространения знаний Университета Арканзаса стр. 5–7. Получено 21 февраля 2016.
- ^ Менели, Филипп (2014). Генетический анализ: гены, геномы и сети у эукариот (2-е изд.). Издательство Оксфордского университета. п. 373. ISBN 978-0-19-968126-6.
- ^ Нельсон Рэнди Дж. (2005) Введение в поведенческую эндокринологию (стр.189). Сандерленд, Массачусетс: Sinauer Associates.
- ^ Фостер, Рассел; Уильямс, Робин (5 декабря 2009 г.). «Внесетчатые фоторецепторы» (Опрос). Научное шоу. ABC Radio National. Получено 2010-05-28.
... у нас есть эволюционный багаж демонстрации сезонности, но мы не совсем уверены, что это за механизм.
дальнейшее чтение
- D.E. Фоскет, Рост и развитие растений, молекулярный подход. Academic Press, Сан-Диего, 1994, стр. 495.
- Б. Томас и Д. Винс-Прю, Фотопериодизм у растений (2-е изд.). Academic Press, 1997 г.
| Внутренние ритмы |  | |
|---|---|---|
| Внешние циклы | ||
| Поля | ||
| Смотрите также | ||