FAM76A - FAM76A
| FAM76A | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||||||||||||||||
| Псевдонимы | FAM76A, семейство со сходством последовательностей 76 член A | ||||||||||||||||||||||||
| Внешние идентификаторы | MGI: 2385211 ГомолоГен: 27071 Генные карты: FAM76A | ||||||||||||||||||||||||
| |||||||||||||||||||||||||
| |||||||||||||||||||||||||
| |||||||||||||||||||||||||
| Ортологи | |||||||||||||||||||||||||
| Разновидность | Человек | Мышь | |||||||||||||||||||||||
| Entrez | |||||||||||||||||||||||||
| Ансамбль | |||||||||||||||||||||||||
| UniProt | |||||||||||||||||||||||||
| RefSeq (мРНК) | |||||||||||||||||||||||||
| RefSeq (белок) | |||||||||||||||||||||||||
| Расположение (UCSC) | Chr 1: 27.73 - 27.76 Мб | Chr 4: 132,9 - 132,92 Мб | |||||||||||||||||||||||
| PubMed поиск | [3] | [4] | |||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||
| |||||||||||||||||||||||||




FAM76A белок, который в Homo sapiens кодируется FAM76A ген.[5] Известные структурные характеристики FAM76A включают 83 аминокислоты. спиральная катушка домена, а также смещение состава полисерина из четырех аминокислот.[6] FAM76A сохраняется в большинстве хордовые но его нет в других дейтеростром фля, такая как иглокожие, гемихордовые, или же ксенакоеломорфа - предположение, что FAM76A возник где-то после хордовых в эволюционной линии. Кроме того, FAM76A не встречается в грибы, растения, археи, или же бактерии.[7] Ожидается, что FAM76A будет локализован на ядро и может сыграть роль в регулирование транскрипции.[8]
Ген
Место расположения

FAM76A расположен на (+) нити короткого плеча хромосома 1 (1p35.3) с геномной последовательностью, начинающейся с 27725979 и заканчивающейся 27762915. Кодирующая область состоит из 3462 пар оснований и транслируется в 341 аминокислоту.[5][10]
Джин соседство
Гены, фланкирующие FAM76A на теломерной стороне, включают IFI6, CHMP1AP1 и RPEP3, тогда как гены, фланкирующие FAM76A на центромерной стороне, включают STX12, PPP1R8 и L0C105376894.[5]
Общие псевдонимы
В Caenorhabditis elegans, FAM76A обозначается как K04F10.7.[11] Помимо этого, FAM76A не имеет каких-либо значимых альтернативных имен.
мРНК
В Homo sapiens, то FAM76A Ген продуцирует 9 различных мРНК, 7 из которых являются альтернативными сплайсерами, а 2 из которых не сплайсированы. Из альтернативно сплайсированных мРНК изоформа 1 является самым длинным вариантом гена и является предметом данной статьи.[5]
Протеин
Общие свойства
Молекулярная масса FAM76A составляет 38,4 кДа, позволяя этому белку диффундировать через ядерные поры.[12] Изоэлектрическая точка - 9,28. FAM76A не имеет каких-либо значительных кластеров положительного, отрицательного или смешанного заряда. Кроме того, FAM76A не имеет каких-либо предсказанных гидрофобных или трансмембранных сегментов, что позволяет предположить, что этот белок не обнаружен внутри клеточной мембраны.[13]
Сочинение
Аминокислотный состав белка FAM76A показал частотность аминокислот в пределах 1,5% от таковой для нормальных белков человека для всех, кроме цистеина, валина и лизина. Цистеин и лизин имеют более высокие частоты по сравнению с нормальным Homo sapiens белка, в то время как валин имеет более низкую частоту по сравнению с нормальным Homo sapiens белок. Такие же различия в частоте встречаемости аминокислот наблюдаются в ортологах FAM76A, таких как Gallus gallus (Х. сапиенс идентичность последовательностей 84%), Серин канария (Х. сапиенс идентичность последовательностей 77%), и Crassostrea gigas (Х. сапиенс идентичность последовательностей 57%).
Домены и мотивы

Поиск консервативных доменов NCBI выявил неохарактеризованный консервативный белок (YqiK), который содержит домен Band7 / PHB / SPFH, функция которого неизвестна и сохраняется у различных видов, от людей до бактерий.[10] В Homo sapiens, домен Band7 / PHB / SPFH состоит из аминокислот 252-326. Молекулярная масса этого домена составляет 8,9 кДа, а его изоэлектрическая точка составляет 9,23. Домен Band7 / PHB / SPFH не имеет частотного состава аминокислот, отличного от нормального. Homo sapiens белок.[13] Этот домен еще не был отнесен к какому-либо суперсемейству доменов.
Вторичная структура

Предполагается, что FAM76A будет иметь только альфа-спирали. Всего предсказано 17 альфа-спиралей, самая длинная из которых содержит домен Band7 / PHB / SPFH.[14] Из этого только 8 альфа-спиралей расположены в консервативных областях FAM76A (см. Концептуальный перевод).
Третичная / четвертичная структура
FAM76A содержит домен coiled-coil, который расположен в домене Band7 / PHB / SPFH. I-TASSER не предсказал никаких значимых сайтов связывания лиганда или активных сайтов.[15] Нет никаких доказательств того, что FAM76A взаимодействует с другими белками с образованием четвертичная структура.
Субклеточная локализация
Инструмент прогнозирования субклеточной локализации белка, PSORT II, предсказывает, что FAM76A находится внутри ядра. Это предсказание наблюдается у ортологов, таких как Gallus gallus и Callorhinchus milii.[16] Дальнейшее свидетельство локализации FAM76A в ядре обеспечивается наличием сигнала ядерной локализации.[8]
Выражение
Согласно NCBI Geo Profile, FAM76A выражается в Homo sapiens паращитовидная железа, лимфатический узел, пищевод, и Костный мозг ткань. Стадии развития, на которых обнаруживается экспрессия FAM76A, включают эмбриоидное тело, плод и взрослую особь.[17]
Атлас мозга
Атлас человеческого мозга аллена предсказания для экспрессии FAM76A изображены ниже. FAM76A, по-видимому, имеет более высокую экспрессию в кора головного мозга и более низкое выражение в частях рептильный мозг такой как pontine tegmentum (подробности см. в таблице выражений).[18]
- Различные виды мозга на экспрессию FAM76A у Homo sapiens: зеленый цвет означает высокую экспрессию, а красный - низкую.

Вид спереди

Каудальный вид
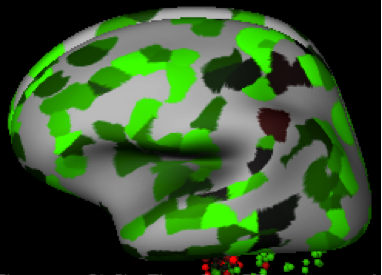
Сагиттальный вид


| Область мозга | Функция | Уровень экспрессии FAM76A |
|---|---|---|
| Лобная доля | Планирование, организация, решение проблем, избирательное внимание, личность и высшие когнитивные функции | Высоко |
| Затылочная доля | Визуальная обработка | Высоко |
| Височная доля | Слуховая обработка | Высоко |
| Теменная доля | Ощущение, восприятие и интеграция сенсорного ввода | Высоко |
| Мозжечок | Координация произвольных движений | Высоко |
| Формирование гиппокампа | Память / пространственное кодирование | Низкий |
| Pontine tegmentum | Сенсорная и двигательная функции, контроль стадии сна и возбуждение | Низкий |
| Клинчатое ядро продолговатого мозга | Получайте тонкие прикосновения и проприоцептивную информацию от верхней части тела | Низкий |
Экспериментальные данные
Ниже показаны избранные данные трех экспериментов с FAM76A. В одном эксперименте сверхэкспрессия CLDN1 в клетках аденокарциномы легких снижала экспрессию FAM76A.[19] В другом эксперименте было показано, что клетки рака простаты, нечувствительные к андрогенам, имеют пониженную экспрессию FAM76A по сравнению с клетками, чувствительными к андрогенам.[20] Другой эксперимент продемонстрировал, что клетки ооцитов в метафазе II обладают большей экспрессией FAM76A по сравнению с контрольными клетками.[21]
- Выбранные географические данные NCBI

Уровни FAM76A в Х. сапиенс Клеточные линии аденокарциномы легких сравнивали между культурами, которые имели клаудин-1 (CLDN1) сверхэкспрессия и контрольные клетки

Экспрессия FAM76A в Homo sapiens чувствительные к андрогенам и нечувствительные линии клеток рака простаты

FAM76A выражение между Х. сапиенс ооциты метафазы II и контрольные линии клеток (состоящие из смеси скелетных мышц, почек, легких, толстой кишки, печени, селезенки, груди, мозга, сердца и желудка)


Регулирование выражения
Посттрансляционные модификации
Предполагается, что FAM76A претерпит множество посттрансляционных модификаций. Посттрансляционные модификации, обнаруженные в консервативных областях, включают 7 сайтов фосфорилирования, 2 сайта сумоилирования и 1 сигнал ядерной локализации.[22] Эти модификации указывают на то, что FAM76A локализован в ядре. См. Концептуальный перевод для визуального представления вышеупомянутых модификаций.
- Концептуальный перевод для FAM76A



Ключ концептуального перевода


Промоутер
Программа Genomatrix ElDorado предсказывает промотор для FAM76A, который называется GXP_71042 и состоит из 679 пар оснований. Он расположен на хромосоме 1, начиная с 27725479 и заканчивая 27726157. GXP_71042 перекрывается с началом кодирующей последовательности FAM76A.[23] Есть несколько факторов транскрипции, которые связываются с этим промотором. Многие из факторов транскрипции, которые связываются с промоторной областью FAM76A, имеют функцию, связанную с кровяные клетки, то иммунная система, и лейкоциты - возможно, это указывает на то, что FAM76A участвует в иммунной функции. Также кажется, что наиболее распространенные семейства матриксов включают цинковые пальцы C2H2 и миелоидные цинковые пальцы, предполагая, что эти семейства матриксов могут быть сильно вовлечены в транскрипцию FAM76A.
РНК-связывающие белки
Общие связывающие РНК белки в 3 ’UTR FAM76A включают: PABPC1, ELAVL1, и PUM2 - каждый с предсказанными частотами связывания 32, 18 и 16 раз соответственно.[24]
Взаимодействующие белки
Было обнаружено, что FAM76A физически взаимодействует с ELAVL1. Взаимодействие было обнаружено методом иммунопреципитации Abdelmohsen et al., 2009.[25] ELAVL1 участвует в регуляции экспрессии генов.
Гомология / эволюция
Паралоги
FAM76B является паралогом FAM76A. По оценкам, FAM76A и FAM76B расходились друг от друга примерно на 17,5 млн лет.[5] Структурные сходства, которые сохраняются между FAM76A / B, включают домен спиральной спирали, а также смещение состава полисерина.[10] FAM76A и FAM76B оба проявляют высокую экспрессию в таких тканях, как лимфатический узел, цельная кровь, яички, яичники, мозг, почки, печень и легкие.[26] FAM76B имеет примерно 62% идентичности последовательности с FAM76A.[10]
Ортологи
| Род и вид | Распространенное имя | Дата расхождения (MYA) | Идентичность аминокислотной последовательности (%) |
| Homo sapiens | люди | 0 | 100 |
| Macaca fascicularis | крабоядная макака | 29.1 | 95 |
| Tarsius syrichta | Филиппинский долгопят | 67.6 | 85 |
| Dipodomys ordii | Кенгуровая крыса Орда | 90.9 | 85 |
| Nannospalax galili | слепой землекоп | 90.9 | 88 |
| Gallus gallus | красная джунглевая птица | 320.5 | 84 |
| Ниппония Ниппон | хохлатый ибис | 320.5 | 83 |
| Egretta garzetta | маленькая цапля | 320.5 | 75 |
| Анолис каролинский | Каролина анол | 320.5 | 73 |
| Oryzias latipes | Японская рисовая рыба | 429.6 | 59 |
| Callorhinchus milii | Австралийская акула-призрак | 482.9 | 64 |
| Crassostrea gigas | тихоокеанская устрица | 847 | 57 |
| Cryptosporidium parvum Iowa II | Нет данных | 1724.7 | 27 |
| Cryptosporidium hominis | Нет данных | 1724.7 | 26 |
Здесь представлена таблица избранного количества ортологов для Homo sapiens FAM76A. В таблицу включены близкие, средние и отдаленные ортологи. Показано, что млекопитающие имеют большее сходство, в то время как водные позвоночные, такие как актиноптеригии /хондрихи имеют меньшее сходство. Ортологи Homo sapiens белок FAM76A перечислены выше в порядке убывания даты расхождения, а затем по идентичности последовательностей.
Эволюция
FAM76A, по-видимому, имеет умеренную скорость мутации по сравнению с фибриноген (быстрая мутация) и цитохром с (медленные мутации).[7][27] Это говорит о том, что FAM76A был по крайней мере в некоторой степени устойчив к мутации в ходе эволюции.

Клиническое значение
Ассоциация болезней
Экспрессия FAM76A наиболее высока в опухоли надпочечников, опухоли пищевода и опухоли мягких / мышечных тканей.[5][28] Увеличение / уменьшение количества копий FAM76A - вместе с соседними генами - показало, что они вызывают вредные фенотипы. В одном отчете о случае у пациента с увеличением числа копий с 1p36.11-34.2 была обнаружена задержка в развитии.[29] Другой пациент, у которого было увеличение числа копий с 1p36.1-35, показал аналогичные задержки.[30] В другом отчете о случае у пациента с потерей числа копий 1p35.3, точное местоположение FAM76A, развилось макроцефалия.[30]
Множественное выравнивание последовательностей (MSA)
MSA, показанный ниже и созданный с помощью Biology Workbench CLUSTALW, упорядочивает ортологи по первой букве рода, а затем по первым двум буквам вида.[13] Существует 3 домена, которые являются высококонсервативными в ортологах. Два из этих доменов имеют неизвестную функцию, а третий домен представляет собой спиральный домен. Сохранение этих регионов восходит к Cryptosporidium parvum Iowa II, который расходился с Homo sapiens 1724,7 млн. Лет Консервативная область 1 содержит в основном полярные аминокислоты; консервативная область 2 содержит как полярные, так и неполярные аминокислоты; а домен спиральной спирали содержит в основном полярные аминокислоты.
- Выравнивание множественных последовательностей удаленного ортолога FAM76A

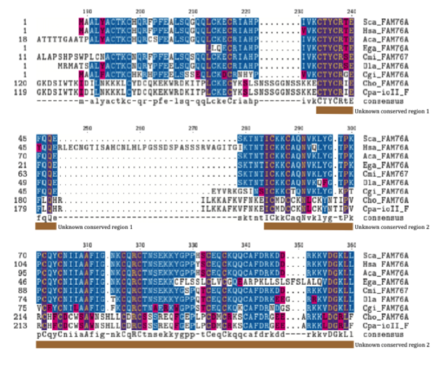



Фиолетовый = похожий химический состав аминокислот
Синий = та же аминокислота


Рекомендации
- ^ а б c ГРЧ38: Ансамбль выпуск 89: ENSG00000009780 - Ансамбль, Май 2017
- ^ а б c GRCm38: выпуск Ensembl 89: ENSMUSG00000028878 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ "Ссылка на Mouse PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ а б c d е ж «Семейство FAM76A со сходством последовательностей 76 член A [Homo sapiens (человек)] - Ген - NCBI». www.ncbi.nlm.nih.gov. Получено 2016-02-08.
- ^ "FAM76A Uniprot Entry".
- ^ а б "NCBI BLAST".
- ^ а б «NLS Mapper».
- ^ «Запись гена NCBI FAM76A».
- ^ а б c d "белок FAM76A изоформа 1 [Homo sapiens] - белок - NCBI". www.ncbi.nlm.nih.gov. Получено 2016-02-08.
- ^ «Профиль Wormbase K04F10.7».
- ^ Махаджан Р., Дельфин С., Гуан Т., Джерас Л., Мельхиор Ф. (январь 1997 г.). «Небольшой убиквитин-родственный полипептид, участвующий в нацеливании RanGAP1 на комплексный белок RanBP2 ядерных пор». Клетка. 88 (1): 97–107. Дои:10.1016 / S0092-8674 (00) 81862-0. PMID 9019411.
- ^ а б c "SDSC Biology Workbench". workbench.sdsc.edu. Получено 2016-02-28.
- ^ "Phyre2".[постоянная мертвая ссылка ]
- ^ «Прогнозы структуры и функций белка I-TASSER».
- ^ «Прогноз PSORT II».
- ^ "NCBI Geo Profile".
- ^ "Атлас мозга Аллена FAM76A".
- ^ Chao YC, Pan SH, Yang SC, Yu SL, Che TF, Lin CW, Tsai MS, Chang GC, Wu CH, Wu YY, Lee YC, Hong TM, Yang PC (январь 2009 г.). «Клаудин-1 является супрессором метастазов и коррелирует с клиническим исходом аденокарциномы легких». Американский журнал респираторной медицины и реанимации. 179 (2): 123–33. Дои:10.1164 / rccm.200803-456OC. PMID 18787218.
- ^ Чжао Х., Ким Й., Ван П., Лапойнт Дж., Тибширани Р., Поллак Дж. Р., Брукс Дж. Д. (май 2005 г.). «Полногеномная характеристика вариаций экспрессии генов и изменений числа копий ДНК в клеточных линиях рака простаты». Простаты. 63 (2): 187–97. Дои:10.1002 / pros.20158. PMID 15486987.
- ^ Коджабас А.М., Кросби Дж., Росс П.Дж., Оту Х.Х., Бейхан З., Кан Х., Там У.Л., Роза Г.Дж., Халгрен Р.Г., Лим Б., Фернандес Э., Сибелли Дж.Б. (сентябрь 2006 г.). «Транскриптом ооцитов человека». Труды Национальной академии наук Соединенных Штатов Америки. 103 (38): 14027–32. Дои:10.1073 / pnas.0603227103. ЧВК 1599906. PMID 16968779.
- ^ "Пост-транснациональные модификации ExPasy".
- ^ "Геноматрица Эльдорадо".[постоянная мертвая ссылка ]
- ^ «RBPDP: База данных специфичностей РНК-связывающих белков».
- ^ Абдельмохсен К., Срикантан С., Ян Х, Лал А., Ким Х. Х., Кувано И., Гальбан С., Беккер К. Г., Камара Д., де Кабо Р., Гороспе М. (май 2009 г.). «Убиквитин-опосредованный протеолиз HuR тепловым шоком». Журнал EMBO. 28 (9): 1271–82. Дои:10.1038 / emboj.2009.67. ЧВК 2683047. PMID 19322201.
- ^ "NCBI AceView".
- ^ «Дерево времени».
- ^ «Генные карты FAM76A».
- ^ Миллер Д.Т., Адам М.П., Арадья С., Бизекер Л.Г., Бротман А.Р., Картер Н.П., Чёрч Д.М., Кролла Дж. А., Эйхлер Э., Эпштейн С.Дж., Фосетт В.А., Феук Л., Фридман Дж. М., Хамош А., Джексон Л., Каминский Э. Б., Кок К. , Krantz ID, Kuhn RM, Lee C, Ostell JM, Rosenberg C, Scherer SW, Spinner NB, Stavropoulos DJ, Tepperberg JH, Thorland EC, Vermeesch JR, Wagoner DJ, Watson MS, Martin CL, Ledbetter DH (май 2010 г.). «Консенсус: хромосомный микрочип - это клинический диагностический тест первого уровня для людей с пороками развития или врожденными аномалиями». Американский журнал генетики человека. 86 (5): 749–64. Дои:10.1016 / j.ajhg.2010.04.006. ЧВК 2869000. PMID 20466091.
- ^ а б Каминский Е.Б., Каул В., Паскалл Дж., Черч Д.М., Бунке Б., Куниг Д., Морено-Де-Лука Д., Морено-Де-Лука А., Мюлле Дж. Г., Уоррен С. Т., Ричард Дж., Комптон Дж. Г., Фуллер А. Е., Глиэм Т. Дж., Хуанг С., Коллинсон Миннесота, Бил С.Дж., Экли Т., Пикеринг Д.Л., Голден ДМ, Астон Э, Уитби Х., Шетти С., Росси М.Р., Радд М.К., Южный СТ, Бротман А.Р., Сангер РГ, Айер Р.К., Кролла Дж.А., Торланд Э.С. , Арадхья С., Ледбеттер Д.Х., Мартин С.Л. (сентябрь 2011 г.). «Основанный на доказательствах подход к установлению функциональной и клинической значимости вариантов числа копий при интеллектуальных нарушениях и нарушениях развития». Генетика в медицине. 13 (9): 777–84. Дои:10.1097 / GIM.0b013e31822c79f9. ЧВК 3661946. PMID 21844811.
дальнейшее чтение
- Абдельмохсен К., Срикантан С., Ян Х, Лал А., Ким Х. Х., Кувано И., Гальбан С., Беккер К. Г., Камара Д., де Кабо Р., Гороспе М. (май 2009 г.). «Убиквитин-опосредованный протеолиз HuR тепловым шоком». Журнал EMBO. 28 (9): 1271–82. Дои:10.1038 / emboj.2009.67. ЧВК 2683047. PMID 19322201.
- Коджабас А.М., Кросби Дж., Росс П.Дж., Оту Х.Х., Бейхан З., Кан Х., Там У.Л., Роза Г.Дж., Халгрен Р.Г., Лим Б., Фернандес Э., Сибелли Дж.Б. (сентябрь 2006 г.). «Транскриптом ооцитов человека». Труды Национальной академии наук Соединенных Штатов Америки. 103 (38): 14027–32. Дои:10.1073 / pnas.0603227103. ЧВК 1599906. PMID 16968779.