Фиона пинната - Fiona pinnata
| Фиона пинната | |
|---|---|
 | |
| Научная классификация | |
| Королевство: | |
| Тип: | |
| Учебный класс: | |
| (без рейтинга): | клады Гетеробранхий клады Euthyneura клады Нудиплевра клады Нудибранхии клады Дексиархия клады Кладобранхии клады Aeolidida |
| Надсемейство: | |
| Семья: | |
| Род: | |
| Разновидность: | F. pinnata |
| Биномиальное имя | |
| Фиона пинната | |
| Синонимы[11] | |
| |
Фиона пинната, распространенное имя Фиона, это разновидность малых пелагический голожаберный (морской слизень), а морской брюхоногие моллюски моллюск в надсемействе Fionoidea. Этот вид голожаберных обитает во всем мире на плавучих объектах в море и питается в основном ракушки, конкретно гусиные ракушки в роду Лепас.
Анатомия этого вида очень необычна. В настоящее время это единственный названный член род Фиона но исследование 2016 года показало, что этот вид видовой комплекс.[12] В семья Fionidae был расширен в 2016 году, чтобы включить Tergipedidae, Eubranchidae и Calmidae в результате исследования молекулярной филогенетики.[13] Особенности, характерные для рода Фиона включают[14] похожие оральные щупальца и ринофоры; то Cerata с мембраной и без cnidosac; дорсальное анальное отверстие; а репродуктивная система с двумя половыми отверстиями; две челюсти с режущей кромкой, и радула только с одним центральным зубцом в каждом ряду зубов. Этот зубчик имеет центральный бугорок и несколько окружающих бугорков.

Распределение
Фиона пинната встречается во всех морях мира, на многих различных плавучих объектах.[15]
В тип местности это островное расположение Ситка, Аляска (Остров Бараноф ), на крайнем северо-западном побережье Северной Америки.[1]
Таксономия
Исследование 2016 года показало, что этот вид является комплексом видов, но не назвало отдельные виды.[12]
Для этого вида созданы разные названия. Лимакс к Питер Форскол с 1775 г. озабоченный к Йохан Эрнст Гуннерус в 1770 г.[14] Имя Олдера и Хэнкока 1851 года Oithona был озабочен Циклопоид род от W. Baird в 1843 г.,[4][16] поэтому в 1855 году они выбрали Фиону из персонажа из Оссиан.[4] Гарольд Джон Финлей предложил новый род Доличеолис для одного из этих синонимов Eolidia longicauda в 1927 г.[17]
Описание
Туловище удлиненно-продолговато-эллиптическое. Длина тела обычно около 20 мм,[10] но самый большой зарегистрированный размер тела - 50 мм.[11] Размеры экземпляра общей длиной 31,7 мм следующие: 17,7 мм - тело до кончика цераты, длина стопы 14,4 мм, хвост на конце стопы 14 мм.[18] Цвет головы и тела варьируется от белого до коричневого или пурпурного в зависимости от пищи.[19] Стопа длинная и ланцетная, округлая спереди и переходит в тонкое острие сзади.[3] Край стопы тонкий, бахромчатый и смятый, за исключением области головы, где он простой. Он разделен спереди, но не образует передних щупалец.[10]
В Cerata многочисленные, удлиненные, с перепончатой бахромой с внутренней стороны. Cerata может показаться без видимого порядка, но они расположены косыми рядами, содержащими от четырех до шести cerata.[18] Есть также небольшие чешуйки у краев тела.[14] Cerata по бокам спины темно-коричневые, каждая с белым краем. У цераты нет cnidosacs.[10] Они особенно прижаты к основанию.[3]
Фиона пинната не имеет глаз.[8]
В ринофоры просты и напоминают оральные щупальца. Они далекие, шиловидные, сужающиеся и выступают наружу.[8] Они не втягиваются и без карманов.[10]
Ротовые щупальца короче, утолщены у основания, сужаются, выступают в стороны и горизонтально и загнуты назад.[8][18] Рот расположен на нижней поверхности головы. Рот небольшой, внешняя губа разделена сзади по средней линии.[3]
Анус находится между мозговыми оболочками на правой стороне тела, и его отверстие направлено дорсально.[15] Половое отверстие отдельное.[10]
Джошуа Алдер и Олбани Хэнкок (1851)[3] описал ткани Фиона пинната как очень жесткий и твердый.



Пищеварительная система

Пищеварительная система Канал, ведущий ото рта к щечной массе, очень короткий и суженный; и, непосредственно перед тем, как он открывается в щечную массу, он получает с обеих сторон внизу очень тонкий проток от большого, много фолликулированного, слюнных желез. Эти железы лежат под желудком и проходят почти до середины тела. То, что на правой стороне, значительно меньше, чем другое, и несколько трубчатое, - особенно по направлению к его окончанию; тот, что слева, очень сложен по форме, неравномерно и обильно саккулирован. Положение этих желез необычно, но есть и другие виды, такие как Doto fragilis, которые открываются в канал рта раньше, чем образование со стороны рта.[3]
Щечное образование небольшое, довольно длинное, тонкое и неправильной формы. Есть две роговые пластинки или челюсти. по бокам от щечной массы. Он немного удлинен сзади для приема задней части радула, а вокруг расположены мышцы. Мышцы с дорсальной стороны сильно развиты, образуют плотную массу, волокна проходят поперечно, а их конечности вставлены в дорсальные края челюстей. Эти мышцы помогают движению челюстей. Мышцы для перемещения всей буккальной массы вперед состоят из уплощенных и изолированных полос, конечности которых прикреплены к заднему краю челюстей и к мышцам, образующим стенки канала рта.[3]
Фиона пинната имеет две роговые челюсти (нижние челюсти) с зубчатой режущей кромкой.[10] Задняя часть уплощена.[8] Роговые пластинки немного меньше размера буккального образования, сильно удлинены, хорошо изогнуты и имеют яйцевидную форму. (Когда они полностью изолированы, они сильно напоминают по форме створки небольшого двустворчатого моллюска из рода Mytilus ). Они гладкие, блестящие, коричневато-янтарного цвета, наиболее темные по направлению к передней конечности, которая служит опорой для режущего лезвия. Это крыловидный отросток небольшого размера, оканчивающийся снизу свободной точкой и имеющий дугообразный вперед режущий край, ровный и почти под прямым углом к общему направлению пластины. Выше небольшой отросток или точка опоры - точка, в которой две пластины сочленены. Сразу за этой точкой находится дорсальный край пластинок, который отражается и расширяется в дугообразную лопасть для прикрепления мышц.[3] Длина челюсти 2,8 мм. Максимальная ширина и максимальная высота челюсти - 1,3 мм.[18]
 Рисунок челюстей Фиона пинната, также показывая радулу между ними. |  Рисунок внешнего вида одной челюсти Фиона пинната. Режущая кромка находится вверху справа. Слева вверху - расширенная доля дорсального края для прикрепления мышц. |  Рисунок изнутри одной челюсти Fiona pinnata. Режущая кромка находится вверху слева. В центре вверху видна точка сочленения губок. Справа вверху расширенная доля дорсального края для прикрепления мышц. |
В радула опирается на мясистый выступ, который поднимается от дна ротовой полости и простирается в переднезаднем направлении от пищевод к переднему отверстию. Радула длинная, линейная, в форме ремня и состоит из полукруглых и серповидных зубцов (крошечных зубцов) оранжевого цвета.[3] В радуле 40 рядов зубов: 15 самых старых зубцов на переднем конце, затем 22 зубца после угла и три неполных зубца в влагалище на заднем конце радулы.[3][18] Формула радулы: 0 + 1 + 0, что означает, что в каждом ряду есть только один центральный зубчик.[8] В центре остроконечный позвоночник, а 6[8] или 6-7 более мелких шипов с каждой стороны зубца. У основания внешнего края зубца также иногда встречаются крошечные шипы. Все шипы немного изогнуты, а их острия направлены назад, к отверстию пищевода.[3] Общая длина радулы 2,6 мм.[18]
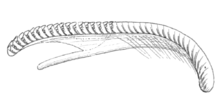 Рисунок сбоку радулы Фиона пинната. Передний конец слева. |  Рисование двух рядов из радулы Фиона пинната. Каждый ряд состоит только из одного зубца, поэтому просматриваются только два зубца. |  Рисунок одиночного зубчика Фиона пинната. |
В пищевод представляет собой короткую и довольно тонкую трубку. Он ведет от верхней части буккальной массы к переднему краю отчетливой грушевидной формы и открывается в нее. желудок. Желудок имеет широкий конец вперед, расположен над репродуктивной системой и лежит в передней части висцеральной полости. Внутренняя поверхность желудка не пластинчатая. В кишечник ведет от заднего конца желудка, слегка наклоняется вправо и переходит назад в трубчатый анус. Анус располагается немного правее средней линии спины, сразу за сердцем. Кишечная трубка довольно короткая, одинакового диаметра на всем протяжении и продольно внутренне складчатая.[3]
Печеночный аппарат у этого животного очень своеобразен. Хвостовой конец желудка получает два желчные протоки, по одному с каждой стороны кишечника. Эти протоки или печеночные каналы почти такой же ширины, как кишечник, и они расходятся по мере выхода из желудка, очень быстро переходят в кожу по бокам спины, где каждый открывается в широкий канал, который простирается почти на всю длину тело. Каналы получают многочисленные ветви, которые сообщаются с железами мозговых оболочек и по мере приближения к боковому расширению сбоку от тела. Эти каналы разделены по несколько раз и расположены неравномерно. Передние отделы больших печеночных каналов связаны с двумя фолликулированными железистыми телами, сильно и неравномерно отвердевшими. Эти железы соединены с кожей, по одной с каждой стороны около области желудка, и, вероятно, образуют внутренние стенки этих частей каналов. Печеночные каналы почти полностью находятся внутри кожи. Печеночные железы большие, почти заполняют головной мозг. Они слабо и неравномерно отсыпаны, с внутренней поверхностью вкладывающей мембраны, выстланной темным зернистым веществом; выше это вещество очень много, образует плотную массу; ниже мембрана некоторых цератов полностью лишена ее.[3]
Расположение печеночных каналов отличается от того, что преобладает у Eolidida. В Эолис, Эмблтония, Дото, Дендронотус, Ломонот, и Антиопа, главные каналы свободно лежат в висцеральной полости, и во всех них есть срединный задний ствол. В этом отношении Oithona казалось бы напоминать Hermaa, в котором все печеночные разветвления, по-видимому, связаны с кожей, и есть только два основных ствола, которые проходят по бокам спины. Однако очевидно, что одна только пищеварительная система в достаточной мере различает Oithona из всех вышеперечисленных родов, даже не исключая HermaaВ центральной части каудального конца тела, позади яичника, также имеется железистое вещество красноватого цвета, фолликулированное и явно разветвленное, соединенное с ветвями печеночных каналов внутри кожи. Эти ветви в задней части тела, вероятно, образуют своего рода сеть трубок на спинной поверхности. Об этом, возможно, можно судить по внешнему виду ветвей, присутствующих при разделении кожи спины по средней линии.[3] Также MacFarland (1966, стр. 357 ).
Репродуктивная система
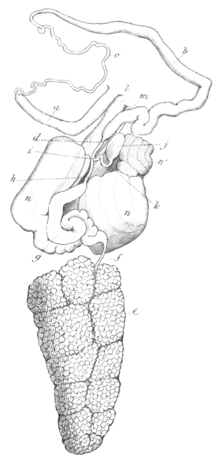
Репродуктивная система: На правой стороне головы за ротовым щупальцем есть два генитальных отверстия: отверстие для полового члена и (гермафродитная) половая пора. Репродуктивная система и слизистые такие же, как у рода Эолидия. Единственное отличие состоит в том, что в Фиона пинната.[3]
При вскрытии кожи спины обнаруживается, что репродуктивные органы, как обычно, занимают большую часть висцеральной полости, при этом желудок и кишечник располагаются сверху, а щечная масса спереди. Пенис расположен впереди других частей и в втянутом состоянии длинный, довольно тонкий и прямой. Пенис во время спаривания имеет коническую форму. Наружный конец полового члена ведет через стенку висцеральной полости к наружному отверстию, и по пути оболочка или внешнее покрытие полового члена прочно прикрепляется к мышцам кожи.[3]
В овотестис (железа гермафродита) желтого цвета с белыми точками.[18] Овотестис заполняет заднюю часть висцеральной полости и состоит из больших неправильных долек, почти полностью состоящих из яиц и упакованных в плотную массу, немного сужающуюся сзади и усеченную спереди.
Яичко представляет собой толстую трубку телесного цвета, свернутую в два-три раза. На одной конечности он сужается в длинный тонкий проток или семявыносящий проток, который соединяется с внутренним концом полового члена. Другой конец яичка внезапно сокращается в такой же тонкий проток, но намного короче, и этим протоком соединяется с яйцеводом.[требуется проверка ]
Спермовод отходит от переднего края яичника в виде тонкой трубки, но, почти сразу же расширяясь, равен диаметру яичка. Эта расширенная часть семенного канала лежит между долями слизистая железа, и сначала несколько мешковато и извилисто. Затем семявыводящий канал проходит вперед и внезапно сокращается до своего первоначального диаметра, а затем продвигается к передней границе слизистой железы и принимает проток от яичка, как описано выше. Затем он немного отклоняется назад и вскоре присоединяется к протоку от сперматеки. Сперматека представляет собой небольшой перепончатый мешок овальной формы, лежащий между долями и у переднего края слизистой железы. Проток, который короткий и тонкий, проходит от одного конца мешка, и в точке соединения протока с яйцеводом к нему присоединяется трубка, которая выходит из внешнего отверстия непосредственно внутри женского отверстия. Эта трубка представляет собой влагалище или копулятивный канал и прикрепляется к верхней стенке женского канала. Непосредственно перед тем, как влагалище достигает протока сперматеки и яйцевода, оно дает ответвление, которое погружается в женский канал, и до сих пор его можно рассматривать как часть яйцевода, поскольку именно по этой ветви яйца находят свое путь к женской розетке.[3]
Слизистая железа, выделяющая слизеподобную оболочку яиц, состоит из двух боковых долей, разделенных на верхней поверхности глубокой трещиной. Эти доли являются полупеллюцидными и образованы грубо извитой трубкой, то есть с правой стороны передней части опаковой и телесного цвета. Две доли открываются в женский канал, широкий и довольно длинный.[3]

Система кровообращения и дыхания


В сердечно-сосудистая система и дыхательная система уникальна для этого животного, потому что почти все эти сосуды отчетливо видны на коже спины, возвышаясь над общей поверхностью. В сердце расположен примерно посередине спины, где он образует большую овальную припухлость непосредственно под кожей, имея под ней половые органы. От заднего конца сердца по средней линии спины до хвостового конца тела проходит широкий приподнятый, но округлый гребень. Этот гребень с обеих сторон соединен многочисленными одинаково приподнятыми ветвями, которые разделяются и разделяются по мере приближения к паллиально-подобному расширению по бокам тела. На всех этих ветвях и их отделах, резко возвышающихся над общей поверхностью кожи, вдоль них расположены жаберные мозговые оболочки, и от них отходят веточки, которые проходят вверх по краю широкого волнообразного перепончатого расширения мозговых оболочек. .[3]
Открыв сердце сверху, желудочек и ушная раковина занимают четко очерченный овал перикард. Желудочек большой и мускулистый, неправильной эллиптической формы, выделяющий спереди аорту, которая обычным образом снабжает ветвями различные органы. К нему присоединяется ушная раковина сзади, немного с левой стороны. Ушная раковина нежная по сравнению с желудочком, но, тем не менее, обильно снабжена мышечными волокнами; он лежит по диагонали в перикарде, причем левая сторона выдвинута почти к передней части того органа, где он принимает ствол вены от кожи. Правая сторона ушной раковины тянется назад и принимает аналогичную стволовую вену из кожи этой стороны почти на заднем конце перикарда.[3]
При открытии дорсальной стенки ушной раковины ее полость оказывается непрерывной с полостью большого заднего приподнятого срединного гребня или стволовой вены, о которой говорилось ранее, а при открытии этой стволовой вены наблюдаются входящие в нее различные боковые ветви. любая сторона. Следовательно, очевидно, что эта стволовая вена, полностью лежащая внутри кожи, является большой задней афферентной или жаберно-сердечной веной, и что все возвышающиеся ветви, идущие к ней от цераты, также являются афферентными сосудами. Таким образом, керата используется для дыхания как специализированный орган дыхания.[3]Оксигенированная кровь из сердца ведет к аорте, к синусам, где она насыщает ткани кислородом. Обезоксигенированная кровь направляется к эфферентным жаберным сосудам головного мозга. Эти эфферентные сосуды можно увидеть на поперечном срезе головного мозга как широкий канал, проходящий вверх по противоположному краю. Из эфферентных сосудов кровь поступает в афферентный сосуд, где получает кислород.[3][18] Кожа спины также частично служит органом дыхания.[3]
Выделительная система
Выделительная система: Почечная пора находится между сердцем и анусом.[18]
Нервная система

Нервная система: The церебральные ганглии помещаются в начало пищевода. Как правило, имеется четыре пары надэзофагеальных ганглиев, хотя на первый взгляд видны только три. Мозговой и жаберный отделы полностью срослись. Жаберные ганглии образуют две овальные центральные массы, лежащие на верхней поверхности пищевода, по одной с каждой стороны от срединной линии, по которой они соединяются на передней конечности короткой, но отчетливой комиссурой. Их задние конечности расходятся и слегка двулопастные, отмечая границы двух ганглиев, из которых состоит каждая масса: передняя доля указывает на цереброид, а задняя - на жаберную. (Жаберные ганглии также срастаются Onchidoris bilamellata И в Lamellidoris aspera.) Педиальные ганглии неправильно округлены, по величине равны цереброидным и жаберным вместе взятым. Они лежат по бокам пищевода и соединяются с нижней поверхностью центральных образований. Четвертая пара ганглиев - обонятельные: они хорошо развиты, хотя намного меньше тех, что были описаны выше, и соединены короткой комиссурой с верхней поверхностью передних краев цереброидных ганглий.[3]
Инфраэзофагеальные ганглии располагаются в обычном положении на буккальном образовании ниже пищевода. Щечные узлы едва ли крупнее обонятельных и имеют овальную форму, их внутренние конечности соединяются через срединную линию короткой комиссурой; их внешние конечности получают связь от каждого из узлов мозга. Двухминутные эллиптические ганглии на передней границе щечных ганглиев почти сидячие; это гастро-цезофагеальные узлы. Таким образом, всего имеется шесть пар ганглиев; четыре над пищеводом и два под ним.[3]
Первая пара нервов идет от обонятельных узлов и большие, но небольшой длины; они разделяются на несколько нитей, когда входят в основание спинных щупалец. Вторая пара проходит от нижней поверхности переднего края цереброидных узлов, недалеко от их соединения с обонятельными ганглиями; эти нервы идут на снабжение верхней поверхности канала рта. Третья и четвертая пары нервов исходят из одних и тех же узлов, но значительно позади второй пары; они также попадают в канал рта; третий, вероятно, посылает ветвь к оральным щупальцам. Рядом с корнем четвертой пары проходит прочный тяж: эти тяжи огибают пищевод и соединяются с внешними конечностями щечных узлов, образуя передний воротничок. Пятая пара нервов, по-видимому, выходит из внешнего края жаберных ганглиев и идет к коже сбоку от головы. Шестая пара маленькие, выходят из верхней поверхности жаберных узлов; эти нервы идут к коже по бокам спины. Седьмой, намного крупнее шестого, выходит из заднего края тех же ганглиев и питает дорсальную кожу и, по-видимому, аналогично cerata. Это жаберные нервы. Восьмая и девятая пары - большие нервы; они выходят из внешней границы педальных узлов и идут к стопе. Задние края этих ганглиев объединены толстой короткой комиссурой, состоящей из двух или трех тяжей, которые, проходя ниже пищевода, образуют большой воротничок пищевода. Десятая пара нервов отходит от заднего края щечных нервов; они проходят через щечную массу и идут к языку. Одиннадцатая пара, отходящая от наружных конечностей буккальных ганглиев, распределяется между мышцами буккального образования. Двенадцатая пара идет от вершины желудочно-пищеводных узлов и, прикладываемая к пищеводу, каждая разделяется на две ветви, одна из которых снабжает верхнюю часть этой трубки, а другая, проходя по ней, идет к желудку в виде в другом голожаберники. Тринадцатая пара крупная; это печеночные нервы; они исходят из буккального образования и, вероятно,[требуется проверка ] (как в роде Эолидия ) связаны по своему происхождению с ганглиями, которые следует рассматривать как принадлежащие к симпатической системе. Сразу же после выхода из буккального образования они соединяются с щечными ганглиями в точке их соединения с желудочно-пищеводным, а затем, выгибаясь наружу и вверх, проходят изнутри переднего воротникового отдела пищевода и направляются к кровоснабжающим железам головного мозга.[3]
Другой нерв включает «генитальный нерв» - единственный нерв, отходящий от тонкого воротничка, концы которого соединены с нижней поверхностью центральных масс, там, где они соединены с педиальными ганглиями. Другой нерв, который, по-видимому, также распространялся на гениталии; это, по-видимому, исходит из правого жаберного ганглия в месте его соединения с педиальной. Эти два нерва, вероятно, идут от висцеральных ганглиев.[3][требуется проверка ][ненадежный источник? ]
Экология
Это голожаберник пелагический аналогично голожаберному Glaucus atlanticus. В отличие от некоторых других пелагических животных, этот вид не может плавать или даже плавать в воде сам по себе.[15] таким образом, хотя это пелагический, он не считается планктонный.
Фиона пинната был даже обнаружен как у взрослых, так и у молодых морские черепахи с Канарских островов.[20]
Кормление
Фиона пинната нападает и охотится на ракушки рода Лепас: гусиная шея Lepas anatifera, Lepas anserifera, Lepas fascicularis, Лепас Хилли, и Лепас тестудината, которые растут на плавающих обломках.[14] Может атаковать других ракушек, но только поврежденных: Pollicipes polyterus и Balanus glandula.[14] Он также может поедать ракообразных из этого рода Алепа[21] (см. также Beeman & Williams 1980) и книдарийцы Велелла Велелла[21] и Порпита порпита.[14] Цвет пищеварительной железы в цератах меняется на ярко-синий, когда животное питается. Велелла.[22] Некоторые авторы отмечают, что Фиона пинната не питается сифонофором Физалия физалис[14] (см. также Bayer 1963), но некоторые авторы упоминают Физалия как его добыча.[15]
Жизненный цикл
Стадион велигер личинка Фиона пинната длится пять дней.[14] Затем он проходит метаморфоза в слизняк. Новозеландский малаколог Ричард Кардеу Уиллан (1979)[14] опубликовал теорию о том, что велигер может отсрочить свою метаморфозу, если не найдет подходящей плавучей среды обитания, к которой можно было бы прикрепиться.
Фиона пинната растет очень быстро. У него один из самых высоких темпов роста среди всех голожаберников (по сравнению с бентосными голожаберниками, единственными, для которых скорость роста известна). Единственный вид, который, как известно, растет быстрее этого, - это Дориделла обскура.[14]
Фиона пинната может вырасти от 8 до 20 мм за 4 дня.[14]

Рисунок переднего вида молодых велигер из Фиона пинната.

Рисунок правой стороны молодого велигера.
Рисунок правой стороны велигера.

Рисунок правой стороны велигера.




Вид спереди хорошо развитого велигера.

Вид сверху хорошо развитого велигера.

Правая сторона велигера перед вылуплением.

Велигер перед вылуплением сверху.




{kind=link}
Рекомендации
Эта статья включает текст из ссылок, являющийся общественным достоянием.[3][8][10]
- ^ а б (на немецком) Эшшольц, Ф. (1831). Зоологический атлас 4. 19 стр., Плс. 16-20. Г. Реймер, Берлин. стр.14, Табл.19, рисунок 1.
- ^ Форскол, П. (1775). Описание Animalium. Mölleri, Hauniae. 20 + xxxv + 164 с., 1 карта.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z аа ab Алдер Дж. & Хэнкок А. (Октябрь 1851 г.). «Описание двух новых видов голожаберных моллюсков, один из которых формирует тип нового рода». Летопись и журнал естественной истории (серия 2) 8 (46): 290 -302, плс. 9-10.
- ^ а б c Алдер, Джошуа; Хэнкок, Олбани (1855). Монография британского голожабернистого моллюска: с изображениями всех видов. Луч Общество. Часть 7, Fam. 3, пл. 38а.
- ^ Берг, Л. С. Р. (1857). "Anatomisk Untersögelse af Фиона атлантика, Бхх. " Videnskabelige meddelelser fra den Naturhistoriske forening в Kjöbenhavn (18-21): 273-335, приложение стр. 336-337, пожалуйста. 2-3.
- ^ (на итальянском) Коста, А. 1866. "Acquisti fatti durante l'anno 1863". Annuario del Museo Zoologico della Università di Napoli 3 (за 1863 год) (2): 13 -41.
- ^ (на итальянском) Коста, А. (1867 г.). "Acquisti fatti durante l'anno 1864". Annuario del Museo Zoologico della Università di Napoli 4 (на 1964 год): 29 -30. Таблица 1, рисунок 1-3.
- ^ а б c d е ж грамм час Хаттон Ф. В. (1882) "Заметки о некоторых Branchiate Mollusca". Сделки Королевского общества Новой Зеландии 14: 162-167, Плита 6, изображение F и I.
- ^ (на немецком) Берг, Л. С. Р. (1894 г.). "Die Opisthobranchien. Отчеты о дноуглубительных работах у западного побережья Центральной Америки до Галапагосских островов, западного побережья Мексики и в Калифорнийском заливе, отвечающие за Александр Агассиз, продолженный Пароход Рыбной комиссии США "Альбатрос", в течение 1891 г. лейтенант. Командир З. Л. Таннер, США, командует. Вестник Музея сравнительной зоологии, Гарвард 25 (10): 125-233, пожалуйста. 1-12. Описание вида на странице 130 -132. Таблица 1, рисунки 13-15.
- ^ а б c d е ж грамм час Сутер Х. (1913) Наставник новозеландского моллюска. Веллингтон, 1120 стр., Стр. 586-587.
- ^ а б Фиона пинната (Эшшольц, 1831 г.). Malacolog версии 4.1.1. База данных по морским моллюскам Западной Атлантики. Проверено 17 декабря 2009 года.
- ^ а б Трики, Дж. С., Тиль, М. и Уотерс, Дж., 2016. Трансокеанское распространение и загадочное разнообразие в космополитическом голожаберном спуске на плотах. Систематика беспозвоночных 30 (3): 290. DOI: 10.1071 / IS15052
- ^ Целла, К; Carmona Barnosi, L .; Екимова, Я; Чичвархин А; Щепетов, Д; Гослинер, Т. М. (2016). Радикальное решение: филогения голожаберного семейства Fionidae. PLoS ONE. 11 (12): e0167800.
- ^ а б c d е ж грамм час я j k Уиллан Р. К. (1979) "Новозеландские данные о местонахождении эолидного голожаберника. Фиона пинната (Эшшольц) ". Тане 25: . PDF
- ^ а б c d Фиона пинната (Эшшольц, 1831 г.). Форум морских слизней. Проверено 17 декабря 2009 года.
- ^ Бэрд, W. (1843). "Род Оитона". Зоолог. 1. Лондон: Джон Ван Ворт. С. 59–61.
- ^ Финли Х. Дж. (23 декабря 1926 г.). «Дальнейший комментарий к систематике моллюсков Новой Зеландии». Сделки и разбирательства Королевского общества Новой Зеландии 57(1927): 320 -485.
- ^ а б c d е ж грамм час я МакФарланд, Ф. М. (8 апреля 1966 г.). «Исследования опистожаберных моллюсков Тихоокеанского побережья Северной Америки». Мемуары Калифорнийской академии наук 6: 354 -358. 546 с. Плита 68, фигуры 23-28. Плита 70, рисунки 11-12.
- ^ Беренс Д. В. (последнее изменение: 7 августа 2002 г.) Фиона пинната Слагсайт. Проверено 17 декабря 2009 года.
- ^ Лоза А. Л. и Лопес-Хурадо Л. Ф. (январь 2008 г.) «Сравнительное исследование эпибионтов пелагических и половозрелых самок логгерхедовых черепах на Канарских островах и островах Зеленого Мыса». стр. 100. В: Маст Р. Б., Хатчинсон Б. Дж. и Хатчинсон А. Х. (ред.). (2007). Материалы двадцать четвертого ежегодного симпозиума по биологии и сохранению морских черепах. Технический меморандум NOAA NMFS-SEFSC-567, 205 стр.
- ^ а б Макдональд Г. Р. и Нибаккен Дж. У. (последнее изменение: 14 декабря 2009 г.) "Список всемирных пищевых привычек голожаберников" В архиве 2010-09-13 на Wayback Machine. Доступ 20 декабря 2009 г. htm В архиве 2009-04-11 на Wayback Machine
- ^ Томпсон, Т. и Браун, Г.Х., 1984. Биология опистожаберных моллюсков, том II. Общество лучей. 229 страниц 41 тарелка, 40 рисунков, стр. 125.
дальнейшее чтение
- Байер, Ф. М. (1963). «Наблюдения за пелагическими моллюсками, ассоциированными с сифонофорами. Велелла и Физалия". Бюллетень морских наук Персидского залива и Карибского бассейна, Университет Майами 13 (3): 454-466.
- Биман Р. Д. и Уильямс Г. К. (1980). Глава 14. Opisthobranchia и Pulmonata: морские слизни и союзники. С. 308–354, плс. 95–111. В: Роберт Х. Моррис, Дональд П. Эбботт и Юджин К. Хадерли. Приливные беспозвоночные Калифорнии, ix + 690 стр., 200 пл. Stanford University Press. См. Страницу 338.
- Берг, Л. С. Р. (1859 г.). "Вклады в монографию рода Фиона, Hanc ". Копенгаген, стр. 1–20. Pls. 1-2.
- Биери, Р. (1966). «Кормовые предпочтения и нормы улитки, Янтина пролонгата, ракушка, Lepas anseriferaголожаберники, Glaucus atlanticus и Фиона пинната, и пищевая сеть в морском нейстоне ". Публикации Морской биологической лаборатории Сето 14: 161–170, плс. III-IV.
- Берн, Р.Ф. (1966). "Описание австралийской Eolidacea (Mollusca: Opisthobranchia). 4. Роды Плевролидия, Фиона, Леархис, и Cerberilla с острова Лорд-Хау ". Журнал Малакологического общества Австралии (10): 21–34.
- Холлеман, Дж. Дж. (1972) "Наблюдения за ростом, питанием, размножением и развитием опистобранха. Фиона пинната (Эшшольц) ". Велигер 15 (2): 142–146.
- Джеффрис, Дж. Г. (1869) Британская конхология: или описание моллюсков, которые сейчас населяют Британские острова и окружающие их моря.. Дж. Ван Ворст, Лондон. Том 5, Страница 35. Тарелка 2, фигура 2.
- Уильямс, М. N. (1978) Щечные железы некоторых эолидных голожаберников (ультраструктура и гистохимия). Неопубликовано. Диссертация на степень магистра, Оклендский университет. 96 стр.
внешняя ссылка
- Пелагические улитки: биология голопланктонных брюхоногих моллюсков Кэрол М. Лалли, Рональд В. Гилмер. через книги Google
- Пауэлл А. В. Б., Новозеландский моллюск, William Collins Publishers Ltd, Окленд, Новая Зеландия, 1979 г. ISBN 0-00-216906-1
- Кастил Д. Б. (Апрель 1904 г.) «Клеточная линия и раннее развитие личинок Фиона марина, голожаберный моллюск ". Труды Академии естественных наук Филадельфии 1 (6): 325-405.
- фото 1
- фото 2[постоянная мертвая ссылка ]
{kind=link}