Грибные тела - Википедия - Mushroom bodies

В грибовидные тела или же тела pedunculata представляют собой пару структур в мозг из насекомые, Другой членистоногие, и немного кольчатые червя (особенно тряпка Platynereis dumerilii ).[2] Также известно, что они играют роль в обонятельное обучение и память. У большинства насекомых грибовидные тела и боковой рог две высшие области мозга, которые получают обонятельную информацию от усик через проекционные нейроны.[3] Впервые они были идентифицированы и описаны французским биологом. Феликс Дюжарден в 1850 г.[4][5]
Структура
Грибные тела обычно описывают как нейропилы, т.е. как плотные сети нейронный процессы (дендрит и терминалы аксонов ) и глия. Они получили свое название от их примерно полусферической формы. чашечка, выпуклость, которая соединяется с остальной частью мозга центральным нервным трактом или цветонос.
Большая часть наших текущих знаний о грибовидных телах основана на исследованиях нескольких видов насекомых, особенно таракан Periplaneta americana, то пчела Apis mellifera, то саранча и плодовая муха Drosophila melanogaster. Исследования грибовидных тел плодовых мушек были особенно важны для понимания генетической основы функционирования грибовидных тел, поскольку их геном был секвенирован, и существует огромное количество инструментов для управления экспрессией их генов.
в мозг насекомого, цветоносы грибовидных тел проходят через средний мозг. В основном они состоят из длинных, плотно упакованных нервных волокон Клетки Кеньона, внутренние нейроны грибовидных тел. Эти клетки были обнаружены в грибовидных телах всех исследованных видов, хотя их количество варьируется. Например, у плодовых мушек их около 2500, а у тараканов - около 200000.
Функция
Грибные тела самые большие в Перепончатокрылые, которые, как известно, особенно тщательно контролируют обонятельный поведение. Однако, поскольку грибовидные тела также встречаются в аносмический примитивные насекомые, их роль, вероятно, выходит за рамки обонятельной обработки. Анатомические исследования предполагают роль в обработке визуальный и механосенсорный ввод у некоторых видов.[6] В Перепончатокрылые в частности, подобласти нейропиля грибовидного тела специализируются на получении обонятельных, визуальных или обоих типов сенсорных сигналов.[7] У перепончатокрылых обонятельные органы располагаются в чашечках. У муравьев можно выделить несколько слоев, соответствующих разным скоплениям клубочков в усиковые доли, возможно, для обработки разных классов запахов.[3][8] Есть две основные группы проекционные нейроны делит усиковую лопасть на две основные области: переднюю и заднюю. Группы проекционных нейронов разделены, иннервируя группы клубочков отдельно и отправляя аксоны отдельными маршрутами, либо через медиально-антенно-протоцеребральный тракт (m-APT), либо через латерально-антенно-протоцеребральный тракт (l-APT), и соединяются с двумя слоями в чашечки грибовидных тел. В этих слоях топографически представлена организация двух эфферентных областей антеннальной доли, устанавливающая грубую одотопный карта антеннальной доли в районе губа грибовидных тел.[3][8]
Известно, что грибовидные тела участвуют в учусь и объем памяти, особенно для запах, и поэтому являются предметом текущих интенсивных исследований. Исследования показывают, что у более крупных насекомых грибовидные тела обладают и другими функциями обучения и памяти, такими как ассоциативная память, сенсорная фильтрация, блок управления двигателем, и место памяти. Исследования показывают, что грибовидные тела обычно действуют как своего рода детектор совпадений, интегрируя мульти-модальный входы[3] и создание новых ассоциаций, таким образом предполагая их роль в обучении и памяти.[9] Недавняя работа также демонстрирует доказательства участия грибовидного тела во врожденном обонятельном поведении через взаимодействие с боковым рогом,[10][11] возможно, используя частично стереотипные сенсорные реакции выходных нейронов грибовидного тела (MBON) у разных людей.[12] Хотя связи между проекционными нейронами и клетками Кеньона случайны (т. Е. Не стереотипны между людьми),[13] Стереотипность в ответах MBON стала возможной благодаря плотной конвергенции многих ячеек Kenyon в несколько MBON наряду с другими свойствами сети.[12]
Информация о запахах может быть закодирована в грибовидном теле с помощью характеристик реагирующих нейронов, а также времени их спайков.[14] Опыты на саранче показали, что Клетки Кеньона синхронизировать свою активность с частотой 20 Гц нейронные колебания и особенно чувствительны к спайкам проекционных нейронов в определенных фазах колебательного цикла.[15]
Drosophila melanogaster

Мы знаем, что строение грибовидного тела важно для обонятельный учусь и объем памяти в Дрозофила потому что их абляция разрушает эту функцию.[17] Грибовидное тело также способно сочетать информацию о внутреннем состоянии тела и обонятельной информации для определения врожденного поведения.[18] Точные роли конкретных нейронов, составляющих грибовидные тела, до сих пор неясны. Однако эти структуры широко изучаются, потому что об их генетический состав. Есть три конкретных класса нейронов, которые составляют доли грибовидного тела: α / β, α ’/ β’ и γ нейроны, которые все имеют различную экспрессию генов. Тема текущих исследований - какие из этих субструктур грибовидного тела участвуют в каждой фазе и процессе обучения и памяти.[19] Дрозофила грибовидные тела также часто используются для изучения обучения и памяти, и ими манипулируют из-за их относительно дискретной природы. Обычно тесты обонятельного обучения состоят из раздельного воздействия на мух двух запахов; один сопряжен с импульсами электрического удара ( условный раздражитель, или CS +), а второй - нет (безусловный стимул, или США). После этого периода обучения мух помещают в Т-образный лабиринт с двумя запахами, размещенными по отдельности на обоих концах горизонтальных Т-образных рычагов. Рассчитан процент мух, избегающих CS +, при этом высокий уровень избегания считается свидетельством обучения и памяти.[20]
Следы сотовой памяти
Недавние исследования, сочетающие кондиционирование запаха и визуализацию клеток, выявили шесть следов в памяти, которые совпадают с молекулярный изменения в Дрозофила обонятельная система. Три из этих следов связаны с ранним формированием поведенческой памяти. Один такой след был визуализирован в усик (AL) автор: синапто-pHфторин репортерные молекулы. Сразу после кондиционирования дополнительный набор проекционные нейроны в наборе из восьми клубочки в AL становится синаптически активированным условным запахом и длится всего 7 минут.[21] Второй след обнаруживается GCaMP экспрессия, и, следовательно, увеличение Ca2+ приток в аксоны α ’/ β’ нейронов грибовидного тела.[22] Это более продолжительный след, который сохраняется до одного часа после кондиционирования. Третий след памяти - это снижение активности переднего парного латерального нейрона, который действует как супрессор формирования памяти через одну из своих тормозных функций. ГАМКергический рецепторы. Снижение кальций ответ нейронов APL и последующее снижение ГАМК высвобождение на грибовидные тела сохранялось до 5 минут после кондиционирования запаха.[23]
Трассировка промежуточной памяти зависит от выражения амн ген расположен в дорсальных парных медиальных нейронах. Увеличение кальций приток и синаптическое высвобождение, которые иннервируют грибовидные тела, становятся обнаруживаемыми примерно через 30 минут после сочетания электрического шока с запахом и сохраняются в течение как минимум часа.[24] Обе карты долговременной памяти зависят от активности и синтеза белка. CREB и CaMKII, и существуют только после кондиционирования с промежутками. Первый след обнаруживается в α / β нейронах между 9 и 24 часами после кондиционирования и характеризуется увеличением кальций приток в ответ на условный запах.[25] Второй след долговременной памяти формируется в γ-грибовидных телах и обнаруживается по увеличению кальций приток от 18 до 24 часов после кондиционирования [26]
динамика цАМФ
Циклический аденозинмонофосфат (цАМФ или циклический АМФ) - это второй мессенджер, который участвует в облегчении грибовидного тела кальций приток в Drosophila melanogaster нейроны грибовидного тела. Повышение цАМФ вызывает пресинаптическую пластичность у дрозофилы. На уровни цАМФ влияют как нейротрансмиттеры, Такие как дофамин и октопамин, и сами запахи. Дофамин и октопамин выделяются грибовидным телом. интернейроны, в то время как запахи непосредственно активируют нейроны в обонятельном пути, вызывая приток кальция через потенциалзависимые кальциевые каналы.[27]
В классическое кондиционирование парадигма, спаривание нейронов деполяризация (через ацетилхолин приложение для представления запах или же CS ) с последующим применением дофамина (чтобы представить шок или нас ), приводит к синергетическому увеличению лагерь в долях грибовидного тела.[27] Эти результаты предполагают, что доли грибовидного тела являются критическим местом интеграции CS / US через действие цАМФ. Этот синергетический эффект первоначально наблюдался в Аплизия, где приток кальция сопряжен с активацией G протеин сигнализация серотонин генерирует аналогичное синергетическое увеличение цАМФ.[28]
Кроме того, это синергетическое увеличение цАМФ опосредуется и зависит от брюква аденилилциклаза (rut AC), которая чувствительна как к кальцию (что является результатом открытия потенциалозависимых кальциевых каналов запахами), так и G протеин стимуляция (вызванная дофамином).[27] В то время как прямое соединение нейронов деполяризация и дофамин (ацетилхолин, а затем дофамин) приводит к синергическому увеличению цАМФ, прямому спариванию нейронных деполяризация октопамин оказывает субаддитивный эффект на цАМФ.[27] Более конкретно, это означает, что эта пара производит значительно меньше цАМФ, чем сумма каждого стимула по отдельности в долях. Следовательно, rut AC в нейронах грибовидного тела работает как детектор совпадений с дофамином и октопамином, действующими двунаправленно, чтобы влиять на уровни цАМФ.[27]
Динамика ПКА
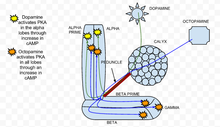
PKA, также известный как протеинкиназа А, было обнаружено, что он играет важную роль в обучении и памяти в Дрозофила.[29] Когда кальций входит в камеру и связывается с кальмодулин, это стимулирует аденилатциклаза (AC), который кодируется ген брюквы (колея).[30] Эта активация переменного тока увеличивает концентрацию лагерь, который активирует PKA.[30] Когда дофамин, аверсивный обонятельный стимулятор, он активирует ПКА, в частности, в вертикальных долях грибовидного тела.[30] Эта пространственная специфика регулируется тупицей (dnc) PDE, цАМФ-специфичный фосфодиэстераза. Если ген тупицы отменен, как обнаружено в dnc мутант, пространственная специфичность не сохраняется. Напротив, стимуляция аппетита, создаваемая октопамин приложение увеличивает ПКА во всех долях.[30] в колея мутант, генотип, в котором брюква отсутствует, ответы на дофамин и октопамин были значительно снижены и близки к экспериментальному шуму.
Ацетилхолин, который представляет собой условный раздражитель, приводит к сильному увеличению активации ПКА по сравнению со стимуляцией только дофамином или октопамином.[30] Эта реакция отменяется в колея мутанты, которые демонстрируют, что PKA важна для сенсорной интеграции.[30] Специфичность активации альфа-доли в присутствии дофамина сохраняется, когда дофамин находится в сочетании с ацетилхолином.[30] По сути, во время парадигмы кондиционирования, когда условный стимул сочетается с безусловным стимулом, ПКА проявляет повышенную активацию. Это показывает, что PKA требуется для условного обучения в Drosophila melanogaster.
Смотрите также
Рекомендации
- ^ Jenett A .; Schindelin J. E .; Гейзенберг М. (2006). «Протокол виртуального мозга насекомого: создание и сравнение стандартизированной нейроанатомии». BMC Bioinformatics. 7: 544. Дои:10.1186/1471-2105-7-544. ЧВК 1769402. PMID 17196102.
- ^ Томер, Р .; Denes, A. S .; Tessmar-Raible, K .; Арендт, Д. (2010). «Профилирование путем совмещения изображений показывает общее происхождение тел кольчатых грибов и паллиума позвоночных». Клетка. 142 (5): 800–809. Дои:10.1016 / j.cell.2010.07.043. PMID 20813265. S2CID 917306.
- ^ а б c d Gronenberg, W .; Лопес-Рикельме, Г.О. (февраль 2014 г.). «Мультисенсорная конвергенция грибовидных тел муравьев и пчел». Acta Biologica Hungarica. 55 (1–4): 31–37. Дои:10.1556 / ABiol.55.2004.1-4.5. PMID 15270216.
- ^ Дюжарден, Ф. (1850). "Mémoire sur le système nerveux des insectes". Анна. Sci. Nat. Zool. 14: 195–206.
- ^ Штраусфельд Н. Дж .; Hansen L; Li Y; Gomez R. S .; Ито К. (1998). «Эволюция, открытие и интерпретация грибовидных тел членистоногих». Учиться. Mem. 5 (1–2): 11–37. Дои:10.1101 / мкм. 5.1.11 (неактивно 11.11.2020). ЧВК 311242. PMID 10454370.CS1 maint: DOI неактивен по состоянию на ноябрь 2020 г. (связь)
- ^ Зарс, Троя (декабрь 2000 г.). «Поведенческие функции грибовидных тел насекомых». Curr Opin Neurobiol. 10 (6): 790–5. Дои:10.1016 / S0959-4388 (00) 00147-1. PMID 11240291. S2CID 5946392.
- ^ Моббс, П. Г. (1982). «Мозг пчелы Apis Mellifera. I. Связи и пространственная организация грибовидных тел». Философские труды Лондонского королевского общества B. 298 (1091): 309–354. Bibcode:1982РСПТБ.298..309М. Дои:10.1098 / рстб.1982.0086.
- ^ а б Лопес-Рикельме, Г.О. (июнь 2014 г.). «Одотопическое афферентное представление организации гломерулярных усиков в грибовидных телах муравьев (Hymenoptera: Formicidae): сравнение двух видов». TIP Revista Especializada en Ciencias Químico-Biológicas. 15 (1): 15–31. Дои:10.1016 / S1405-888X (14) 70317-1.
- ^ Талли, Т; Куинн, WG (сентябрь 1985 г.). «Классическое кондиционирование и удержание у нормальных и мутантных Drosophila melanogaster». J Comp Physiol A. 157 (2): 263–77. Дои:10.1007 / bf01350033. PMID 3939242. S2CID 13552261.
- ^ Долан, Майкл-Джон; Фрехтер, Шахар; Бейтс, Александр Шакил; Дэн, Чунтао; Хуовиала, Пааво; Roberts, Ruairí СП; Шлегель, Филипп; Дхаван, Безмятежный; Табано, Реми; Дионн, Хизер; Кристофору, Кристина; Близко, Кари; Сатклифф, Бен; Джулиани, Бьянка; Ли, Фэн; Коста, Марта; Ирке, Гудрун; Мейснер, Джеффри Уилсон; Бок, Дави Д.; Асо, Йошинори; Рубин, Джеральд М; Джефферис, Грегори SXE (21 мая 2019 г.). «Нейрогенетическое рассечение бокового рога дрозофилы выявляет основные результаты, различные поведенческие функции и взаимодействия с грибовидным телом». eLife. 8: e43079. Дои:10.7554 / eLife.43079. ISSN 2050-084X. ЧВК 6529221. PMID 31112130.
- ^ Льюис, LP; Сиджу, КП; Асо, Y; Фридрих, AB; Bulteel, AJ; Рубин, GM; Грюнвальд Кадоу, ИК (31 августа 2015 г.). «Высший мозговой контур для немедленной интеграции противоречивой сенсорной информации у дрозофилы». Текущая биология. 25 (17): 2203–14. Дои:10.1016 / j.cub.2015.07.015. PMID 26299514. S2CID 16276500.
- ^ а б Миттал, Аруш Мохит; Гупта, Дикша; Сингх, Амрита; Lin, Andrew C .; Гупта, Нитин (24 февраля 2020 г.). «Множественные свойства сети преодолевают случайную связанность, чтобы обеспечить стереотипные сенсорные реакции». Nature Communications. 11 (1): 1023. Bibcode:2020НатКо..11.1023M. Дои:10.1038 / s41467-020-14836-6. ЧВК 7039968. PMID 32094345.
- ^ Карон, SJ; Рута, В; Abbott, LF; Аксель, Р. (2 мая 2013 г.). «Случайная конвергенция обонятельных входов в грибовидном теле дрозофилы». Природа. 497 (7447): 113–7. Bibcode:2013Натура.497..113C. Дои:10.1038 / природа12063. ЧВК 4148081. PMID 23615618.
- ^ Гупта, Нитин; Стопфер, Марк (6 октября 2014 г.). «Временной канал информации в разреженном сенсорном кодировании». Текущая биология. 24 (19): 2247–56. Дои:10.1016 / j.cub.2014.08.021. ЧВК 4189991. PMID 25264257.
- ^ Гупта, Нитин; Сингх, Свикрити Саран; Стопфер, Марк (15.12.2016). «Окна колебательной интеграции в нейронах». Nature Communications. 7: 13808. Bibcode:2016 НатКо ... 713808G. Дои:10.1038 / ncomms13808. ISSN 2041-1723. ЧВК 5171764. PMID 27976720.
- ^ Дэвис, Рональд (2011). «Следы памяти дрозофилы». Нейрон. 70 (1): 8–19. Дои:10.1016 / j.neuron.2011.03.012. ЧВК 3374581. PMID 21482352.
- ^ Макгуайр, Шон; Ле, Фыонг; Дэвис, Рональд (август 2001 г.). "Роль Дрозофила передача сигналов грибовидного тела в обонятельной памяти ". Наука. 17 (293): 1330–33. Bibcode:2001Научный ... 293.1330М. Дои:10.1126 / science.1062622. PMID 11397912. S2CID 23489877.
- ^ Bräcker, L.B .; Siju, K. P .; Varela, N .; Aso, Y .; Zhang, M .; Hein, I .; Кадоу, И.С.Г. (2013). «Существенная роль грибовидного тела в зависимом от контекста избегании СО2 у дрозофилы». Текущая биология. 23 (13): 1228–1234. Дои:10.1016 / j.cub.2013.05.029. PMID 23770186. S2CID 15112681.
- ^ Йылдызоглу, Тугче; Вайслогель, Ян-Марек; Мохаммад, Фархан; Чан, Эдвин С.-Й .; Assam, Pryseley N .; Кларидж-Чанг, Адам (8 декабря 2015 г.). "Оценка обработки информации в системе памяти: применение метааналитических методов в генетике". PLOS Genet. 11 (12): e1005718. Дои:10.1371 / journal.pgen.1005718. ISSN 1553-7404. ЧВК 4672901. PMID 26647168.
- ^ Акалал, Давид-Бенджамин; Уилсон, Кертис; Цзун, Линь; Танака, Нобуаки; Ито, Кей; Дэвис, Рональд (сентябрь 2006 г.). «Роль нейронов грибовидного тела дрозофилы в обонятельном обучении и памяти». Обучение и память. 13 (1): 659–68. Дои:10.1101 / лм. 221206. ЧВК 1783621. PMID 16980542.
- ^ Ю, Динхуэй; Пономарев, Артем; Дэвис, Рональд (май 2004 г.). «Измененное представление пространственного кода запахов после обонятельного классического кондиционирования; образование следов памяти путем синаптического набора». Нейрон. 42 (3): 437–49. Дои:10.1016 / S0896-6273 (04) 00217-X. PMID 15134640. S2CID 5859632.
- ^ Ван, Ялин; Мамира, Акира; Чан, Анн-шын; Чжун, И (апрель 2008 г.). «Визуализация раннего следа памяти в грибовидном теле дрозофилы». Журнал неврологии. 28 (17): 4368–76. Дои:10.1523 / jneurosci.2958-07.2008. ЧВК 3413309. PMID 18434515.
- ^ Сюй, Лю; Дэвис, Рональд (январь 2009 г.). «ГАМКергический передний парный латеральный нейрон подавляется и подавляется обонятельным обучением». Природа Неврология. 12 (1): 53–59. Дои:10.1038 / нн.2235. ЧВК 2680707. PMID 19043409.
- ^ Ю, Динхуэй; Кин, Алекс (декабрь 2005 г.). «Нейроны DPM дрозофилы образуют отложенный и специфичный для ветвей след памяти после обонятельного классического кондиционирования». Клетка. 123 (5): 945–57. Дои:10.1016 / j.cell.2005.09.037. PMID 16325586. S2CID 14152868.
- ^ Ю, Динхуэй; Акалал, Бенджамин-Давид (декабрь 2006 г.). «Нейроны грибовидного тела дрозофилы a / b образуют специфичный для ветвей долговременный след клеточной памяти после разнесенного обонятельного кондиционирования». Нейрон. 52 (1): 845–55. Дои:10.1016 / j.neuron.2006.10.030. ЧВК 1779901. PMID 17145505.
- ^ Акалал, Бенджамин-Давид; Ю, Динхуэй (декабрь 2010 г.). «Поздняя фаза, следовые формы долговременной памяти в γ-нейронах грибовидных тел дрозофилы после обонятельного классического кондиционирования». Журнал неврологии. 30 (49): 16699–16708. Дои:10.1523 / jneurosci.1882-10.2010. ЧВК 3380342. PMID 21148009.
- ^ а б c d е Томчик, Сет; Дэвис, Рональд (ноябрь 2009 г.). «Динамика связанной с обучением передачи сигналов цАМФ и интеграции стимулов в обонятельный путь дрозофилы». Нейрон. 64 (4): 510–21. Дои:10.1016 / j.neuron.2009.09.029. ЧВК 4080329. PMID 19945393.
- ^ Абрамс, Томас; Карл, Кевин; Кандел, Эрик (сентябрь 1991). «Биохимические исследования конвергенции стимулов во время классического кондиционирования при аплизии: двойная регуляция аденилатциклазы Ca2 + / кальмодулином и медиатором». Журнал неврологии. 11 (9): 2655–65. Дои:10.1523 / JNEUROSCI.11-09-02655.1991. ЧВК 6575265. PMID 1679120. S2CID 16477962.
- ^ Скулакис, EM; Кальдерон, Д; Дэвис, Р.Л. (1993). «Преимущественная экспрессия в грибовидных телах каталитической субъединицы протеинкиназы А и ее роль в обучении и памяти». Нейрон. 11 (2): 197–201. Дои:10.1016 / 0896-6273 (93) 90178-т. PMID 8352940. S2CID 23105390.
- ^ а б c d е ж грамм Герваси, Николас; Ченио, Поль; Preat, Томас (февраль 2010 г.). «Динамика PKA в учебном центре дрозофилы: обнаружение совпадений с помощью аденилилциклазы брюквы и пространственной регуляции с помощью фосфодиэстеразы Dunce». Нейрон. 65 (4): 516–529. Дои:10.1016 / j.neuron.2010.01.014. PMID 20188656. S2CID 14318460.
дальнейшее чтение
- Мартин Гейзенберг (1998). "Что делают грибовидные тела для мозга насекомых? Введение". Учиться. Mem. 5 (1): 1–10. ЧВК 311238. PMID 10454369.
- S.E. Фарбах (2006). «Строение грибовидных тел мозга насекомых». Анну. Преподобный Энтомол. 51 (3): 209–232. Дои:10.1006 / jtbi.2001.2504. PMID 12183125.
- Лопес-Рикельме, Херман Октавио (2008). Hormigas como sistemas modelo para el comportamiento complete. Базы нейробиологического исследования в кимике и дивизион дель трабахо в лас гормигас (Кандидат наук.). Universidad Nacional Autónoma de México. Дои:10.13140 / RG.2.1.3145.1689.