Прогнатодон - Википедия - Prognathodon
| Прогнатодон | |
|---|---|
 | |
| P. overtoni ископаемое в Королевский музей Тиррелла | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Рептилии |
| Заказ: | Squamata |
| Надсемейство: | †Мосазавройда |
| Семья: | †Mosasauridae |
| Подсемейство: | †Mosasaurinae |
| Род: | †Прогнатодон Долло, 1889 год |
| Типовой вид | |
| Прогнатодон солвайи Долло, 1889 год | |
| Другие виды | |
Спорные виды
| |
| Синонимы | |
| |
Прогнатодон вымерший род морской ящерицы, принадлежащей мозазавр семья. Классифицируется как часть Mosasaurinae подсемейство, наряду с родами как Мозазавр и Clidastes. Прогнатодон извлечен из отложений возрастом от Кампанский к Маастрихтский в Средний Восток, Европа, Новая Зеландия, и Северная Америка.[1]
Прогнатодон означает «зуб передней челюсти», происходящий от латинский профи- («ранее» или «ранее»), Греческий гнатос («челюсть») и odṓn ("зуб"). Двенадцать номинальных видов Прогнатодон признаны, из Северная Америка, северный и западная африка, то Средний Восток, западная Европа и Новая Зеландия. Из-за иногда явных различий между ними и неполного характера многих экземпляров систематика рода и того, какие виды следует должным образом учитывать. Прогнатодон является спорным.[2]
Прогнатодон известен своими массивными челюстями и зубами. Его особые пищевые приспособления вызвали большой интерес к его экологии с момента его открытия, хотя прямые доказательства его диеты, такие как желудочные остатки, встречаются редко.[2]
Описание

Прогнатодон представляет собой один из самых крупных родов мозазавров с самым большим известным черепом (принадлежащим к P. currii) более 140 см в длину. Несмотря на огромные размеры, остатки этого рода часто бывают фрагментарными и неполными. На сегодняшний день известно очень мало экземпляров с сочлененными черепами и ни одного с полным скелетом.[2] Хотя многие виды были крупными, их размеры приближались или потенциально превышали 10 метров, например P. currii, П. сатуратор и P. overtoni,[2] многие виды были значительно меньше по размеру. В типовой вид, P. solvayi, был самым маленьким, едва достигая 5 метров в длину.[3]
Помимо размера и прочности, еще одна особенность, которая определяет Прогнатодон это форма склеротические кольца, присутствует у всех видов рода. Функция косточек склеры - поддерживать форму косточки. роговица и поддержать склера в области мышцы Брюке, отвечающей за аккомодацию в кружевной глаз. У типовых видов P. solvayiсклеротическое кольцо сохранилось лишь частично, состоящее из пяти склеральных косточек в каждом кольце.[4] Подобные склеротические кольца наблюдаются у нескольких других родов мозазавров, таких как Мозазавр.[5]
Последний опубликованный диагноз для рода Прогнатодон был предоставлен Lingham-Soliar and Nolf (1989),[2] и заявляет, что предчелюстная кость отсутствует рострум перед предчелюстными зубами. В префронтальный образует большую часть заднебоковой границы наружных ноздрей, а надглазничное крыло с тяжелыми треугольными крыльями соприкасается с заглазнично-лобной кзади по орбите медиальнее внешнего края лобной кости. Фронтальная кость над орбитами не вырезается, а срединный дорсальный гребень либо присутствует, либо отсутствует. В отверстие на теменный от маленького до умеренно большого, расположены спереди на небольшом выступе и плотно охватываются с обеих сторон короткими язычками спереди или на лобно-теменном шве.[5]
Края спинного теменный поверхности параллельны друг другу, а срединная линия черепа - к заднему основанию расходящихся поддерживающих ветвей, которые образуют прямоугольное поле медиальнее теменной кости. Вентральный отросток от посторбитофронтального к скуловому отросткам нечетко отделен от умеренно хорошо обнаженной дорсальной поверхности посторбитофронтального и вентропозаднего отростков на коже. скуловой слабо развита или отсутствует. Плоскоклеточное крыло до теменной кости большое. На дне базиально-затылочной кости имеется глубокая борозда для базилярная артерия.[5]
Супрастапедиальный отросток сливается с инфрастапедиальным отростком на квадратной кости, а барабанные крылья толстые. Ступенчатая ямка имеет форму от почти круглой до эллиптической. Задний отросток на дорсальной поверхности зубной присутствует, зарождается или отсутствует, и зубной камень резко обрывается перед первым зубным зубом. Сама зубная часть содержит от тринадцати до четырнадцати зубов, а крыловидный имеет от семи до восьми зубов. Медиальное крыло венечной кости соприкасается с угловым, передний отросток венечной кости отрывается от надугловой кости и соприкасается с задним отростком зубной кости или оканчивается надпочечной частью без соприкосновения с зубной частью. Ретроартикулярный отросток имеет прямоугольные очертания, перегибается медиально или шнурован латерально. Краевые зубы толстые, двояковыпуклые, гладкие или бороздчатые. Зигосфены и зигантра отсутствуют, зарождаются или большие и функциональные.[5]
Череп

Прогнатодон отмечен как имеющий очень крепкий череп. Череп также демонстрирует приспособление к очень мощной мускулатуре челюсти. Отношение между длиной надвисочного отверстия и общей длиной черепа ранее использовалось в качестве импровизированного измерения силы укуса мозазавра и является относительно высоким в Прогнатодон (0,22 дюйма P. overtoni и П. сатуратор) по сравнению с другими родами, такими как Мозазавр (0,19 дюйма М. hoffmannii ).[6]
Квадраты Прогнатодон, как и род Globidens, соединили супрастапедиальный и инфрастапедиальный отростки, что, возможно, является адаптацией для противодействия сильным силам, испытываемым костью во время укуса. Крепкая мускулатура челюсти в сочетании с относительно короткой и высокой зубной частью привели бы к очень сильному прикусу.[7]
Череп типового экземпляра Прогнатодон сатуратор почти завершена, отсутствует только передняя часть предчелюстной кости и зубные кости. Хотя большая часть передних краевых зубов отсутствует, наклон сохранившихся корней позволяет предположить, что П. сатуратор были выпученные зубы, что также наблюдается у P. solvayi. Дорсальный край зубной кости вогнутый, тогда как вентральный край верхней челюсти слегка выпуклый. Краевые зубы массивные, гладкие и округлые, в отличие от большинства других зубов мозазавра, которые обычно имеют фасеточные и сжатые с боков. Нижняя челюсть П. сатуратор очень высокий и массивный, даже больше, чем у других представителей этого рода. Это сочетается с массивным крыловидным отростком и различными другими частями черепа, такими как височная область и черепная коробка, которые все более толстые по сравнению с другими видами этого рода.[7]
Типовой образец P. lutugini, хотя и неполный, сохраняет значительную часть черепа. Первоначально утверждалось, что образец сохранил небольшую часть предчелюстная кость, хотя Д.В. Григорьев (2013)[8] отметил, что указанная кость в настоящий момент не может быть обнаружена и потенциально отсутствует. Обе крыловидные кости сохранились, хотя правый в настоящее время почти полностью реставрируется из гипс, с двумя оригинальными зубами. Левый крыловидный отросток был значительно более полным, но не имел всех отростков, кроме базисфеноидного. Задний альвеолярный край был очень маленьким, с зубцами, отходящими от тонкого, но выраженного вертикального гребня. Вентральная поверхность базисфеноидного отростка довольно гладкая и отверстие видны над шестым зубом на боковой поверхности крыловидного отростка, а также над положением между шестым и седьмым зубами на медиальной поверхности. В чешуйчатый кость представлена лишь несколькими фрагментами, но может быть отмечена тем, что она сжатая с боков и высокая, как у других видов Прогнатодон. Его задне-вентральная поверхность вогнута для контакта с квадратный. Зубные кости срастаются с задним концом блестящий и переднее лезвие предсуставного лезвия и имеет 13 зубов, по крайней мере, восемь зубов имеют субзубные крипты с некоторыми замещающими зубами, обнаруженными в типичном образце. Субзубные крипты располагаются кзади от функциональных зубов. В короноид имеет седловидную форму и хорошо развитый постеродорсальный отросток, который придает дорсальному краю указанной кости угол около 110 градусов между горизонтальным передним концом и субвертикальным задним крылом. Эта комбинация характеристик помогла определить, что P. lutugini действительно был разновидностью Прогнатодон, поскольку некоторые исследователи относят его к собственному роду ",Доллозавр".[8]
Зубы

Упомянутые виды Прогнатодон различаются по зубному ряду. Крепкие конические зубы с тупыми зубчатыми килями и гладкой эмалью часто считаются типичными. Прогнатодон зубы и обычно относят к роду. Несмотря на это, типовой вид P. solvayi отличается от зубов этого типа и вместо этого демонстрирует заметно сжатые лабиально-лингвистические и граничные зубы с мягкими фасетками. Также существует определенная степень вариации количества зубов, P. solvayi имеет 12 зубов на верхней челюсти и 13 на зубных, в то время как P. overtoni имеет 14 зубных зубов.[5]
Всех видов, P. solvayi имеет самые разные зубы от других представителей этого рода. Коронки зубов, как правило, большие и довольно сильно исчерченные, а передние зубы более выпуклые, чем у любых других мозазавров. Предчелюстные зубы почти горизонтальны, а передние зубные - лишь немного меньше. Помимо глубоких штрихов, фрагментарный материал P. solvayi показывает, что коронки зубов также могли быть призматическими с семью-восемью призмами на внешней поверхности.[5]
Зубы P. solvayi более тонкие на переднем конце, более широкие и треугольные к середине зубной ветви, уменьшающиеся в размерах к заднему концу, а последние два зуба довольно низкорослые. Коронки зубов только умеренно вздуты и в отличие от зубов P. overtoni, зубы постоянно увеличиваются в размерах в переднезаднем направлении и несколько реже загибаются назад. В P. overtoni и другие виды (например, P. giganteus), все зубы довольно однородны по размеру, за исключением основания предпоследнего зуба (которое больше).[5]
Еще один вид, известный своими расходящимися зубами, - это P. lutugini, где знания о зубах в основном основаны на отдельных образцах. Известные зубы сильно бикариновые со слабой зубчатостью на обоих килях. Затем кили делят зубы на лингвальную и губную поверхности, при этом язычная поверхность более выпуклая и большая по сравнению с губной. Зубы имеют небольшую заднюю и медиальную изогнутость и отличаются гладкой поверхностью (что отличается от других видов зубов). Прогнатодон) за исключением мелких морщинок на концах коронок зубов. Корни зубов довольно большие, примерно в 1,5 раза больше коронок зубов, и имеют бочкообразную форму. Самые большие известные зубные коронки достигают 5,5 см в высоту и 2,5 см в ширину у основания.[8]
Размер P. lutugini зубы также различаются в зависимости от положения во рту. Отмечается, что передние крыловидные зубы относительно большие и сопоставимы с краевыми зубами по размеру до 4,6 см в высоту.[8]
Относительно небольшое количество зубов по сравнению с другими мозазаврами (например, Прогнатодон сатуратор сохраняет 14 зубов в зубной части, 12 в верхней челюсти и 6 в крыловидном отростке) является характерным признаком всех видов Прогнатодон.[7]
Осевой скелет

Посткраниальные окаменелости Прогнатодон значительно реже, чем материал черепа и изолированные зубы.[2]
Типовой образец P. lutugini сохраняет несколько позвонков. Четыре шейные позвонки сохранились, у которых хорошо развиты передний и задний зигапофизы. Функциональные зигосфены и зигантра были обнаружены на двух шейных позвонках. На всех четырех позвонках обнаружены гипапофизарные ножки позвонков, очень короткие и заканчиваются небольшими сжатыми с боков овальными гранями. Эти фасетки наклонены кзади и расположены кзади на вентральных поверхностях центров. Синапофизы большие, расположены кпереди на центрах и не заходят ниже вентрального края центра. Мыщелки и семядоли дорсовентрально сдавлены слабо. Шейные позвонки почти такого же размера, как самые длинные. спинные позвонки и получите длину около 7,2 см и высоту 4 см. 26 спинные позвонки сохранились в экземпляре, хотя большинство из них плохо сохранились. Спинные позвонки немного длиннее шейных и, как и шейные, мыщелки и семядоли слабо сдавлены дорсовентрально. Максимальные размеры спинных позвонков около 8 см в длину и 5 см в высоту. Кроме того, типовой образец сохранил много ребро фрагменты, правда все неполные. Две бороздки проходят вдоль дистальной части ребер от суставной головки, но за серединой ребра остается только одна бороздка.[8]
Мягких тканей
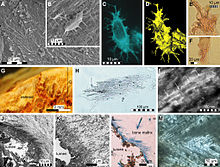
Находка исключительно хорошо сохранившегося экземпляра ERMNH HFV 197 из г. Маастрихтский депозиты в Харрана, центральный Иордания позволили детально изучить уникальные детали морфологии мягких тканей Прогнатодон. Ископаемое не только в значительной степени завершено и четко сформулировано, что редко для Прогнатодон образцов, но также сохраняет значительные части покровов и небольшой изгиб на последних нескольких хвостовых позвонках. Что наиболее важно, окаменелость сохраняет контуры мягких тканей хвостового плавника. Это помогло предоставить доказательства того, что мозазавры сходились с ихтиозавры, Metriorhynchid Thalattosuchians и киты в эволюции хвостовой двуустки в форме полумесяца для помощи в передвижении.[9]
Хвостовая лапа явно асимметрична. Нижняя доля плавника следует за хвостовыми позвонками и при жизни имела бы обтекаемое поперечное сечение, основанное на пропорциях осевого скелета и других мягких тканей. Верхняя доля плавника не поддерживается скелетом и сохранилась в виде небольшой, почти крыловидной структуры над несколькими последними хвостовыми позвонками. Форма хвостовой двуустки подобна форме хвостовой двуустки. кархархинид акулы, хотя и перевернутые, с маленькой верхней и большой нижней лопастями.[9]
На экземпляре также сохранились отпечатки чешуи, особенно вокруг контура хвостовой двуустки. Эти отпечатки, хотя и слабые, обнаруживают чешуйки «ромбовидной» формы.[9] Подобной формы чешуйки присутствуют на хвостовой лапе хорошо сохранившейся Платокарп экземпляр LACM 128319.[10]
Пропорции структур мягких тканей и их отношение к элементам скелета образца могут быть использованы для вывода формы и размера плавников у других видов животных. Прогнатодон и, возможно, также и в других родах мозазавров. В своем описании экземпляра Lindgren et al. (2013)[9] отметил, что образец странно мал по Прогнатодон стандартов по сравнению с некоторыми из более крупных экземпляров этого рода. Таким образом, предполагалось, что ERMNH HFV 197 - это ювенильная особь. Тот факт, что хвостовой плавник вырастет из-за увеличения веса более крупных особей, довольно логичен, и его можно наблюдать в других группах, которые имеют хвостовые плавники, например, в современных акулы и вымершие ихтиозавры. По этой причине вполне вероятно, что взрослый Прогнатодон, особенно у более крупных видов, хвостовые плавники были бы больше по сравнению с размером их тела.[9] Lindgren et al. (2013) особенно отметили, что у более крупных особей верхняя доля плавника, вероятно, увеличилась бы пропорционально.[9]
История открытия

Прогнатодон был впервые описан Луи Долло в 1889 г. по образцам, собранным в Бельгия. Существует некоторая путаница с правильным родовым названием таксона. Долло впервые упомянул таксон как "Прогнатодон"в некоторых предварительных заметках и дал предварительный диагноз, но заменил имя Прогнатодон с "Прогнатозавр"и использовал Прогнатозавр во всех его последующих статьях с упоминанием рода.[5]
Первое более позднее использование "Прогнатодон"был Дейл А. Рассел в обширной монографии о североамериканских мозазаврах 1967 г., где приоритетом Прогнатодон стало очевидным. Рассел также пересмотрел виды, отнесенные к Прогнатодон из Северная Америка , но лишь кратко прокомментировал бельгийские образцы.[4]
Хотя первоначальные останки этого рода были довольно обширными, а первоначальное описание было кратким, никаких дополнительных исследований типового материала не проводилось в течение столетия. Отсутствие исчерпывающего оригинального описания этого рода и видов, относящихся к нему из Бельгии, странным образом не является чем-то необычным для образцов мозазавров, обнаруженных в Формация Craie de Ciply из Бельгия. На извлечение и установку образцов обычно вкладывался большой объем работы, но их научное изучение оставалось ограниченным диагнозами и описаниями, в основном сосредоточенными на специфических точках их анатомии, таких как квадратный и барабанная перепонка из Plioplatecarpus houzeaui. Прогнатодон гигантский, названный Долло в 1904 г. - один из видов с наиболее краткими описаниями, по-видимому, предназначенный только для обозначения скелета мозазавра для выставки в зале музея.[5]

Первое комплексное исследование Прогнатодон образцы из Бельгии (включая типовой образец) были получены Теагартеном Лингхам-Солиар и Дирком Нольфом в 1989 г.[5] и диагноз в этом исследовании остается последним опубликованным исправленным диагнозом для этого рода.[2]
В 1998 году в Маастрихт известняковые карьеры. Вскоре после этого он получил прозвище «Бер» и был выставлен на обозрение в Маастрихтский музей естественной истории. Затем этот образец был идентифицирован как Прогнатодон, и получил видовое название Прогнатодон сатуратор. Этот образец был первым достаточно полным экземпляром мозазавра, извлеченным из Маастрихт площадь с 1957 г .; Скелет, выставленный в Музее естественной истории в Маастрихте, принадлежит животному, длина которого, вероятно, составляла 12 метров.[7]
Очень большой экземпляр найден в Израиль некоторое время неофициально называли "Оронозавр", но в итоге описали как новый вид Прогнатодон, P. currii.[11] Два экземпляра Prognathodon overtoni описан в 2011 г. с раннего конца Кампанский (около 74,5 млн лет) Формация Медвежьей Лапы в Альберта, Канада предоставили первые полностью сочлененные скелеты рода. Детальное изучение этих и ранее обнаруженных экземпляров позволило установить несколько признаков, отличающих Прогнатодон из близкородственных родов, таких как Лиодон и длинноносые мозазаврины. Сохранившиеся зубы и содержимое кишечника также позволили изучить предполагаемую палеоэкологию этого рода.[2]
Новая окаменелость, обнаруженная в 2008 г. и описанная в 2013 г., принадлежала молодому молодому человеку длиной 1,8 м. Прогнатодон был найден на сайте Иордании в Харране. Ископаемое было примечательно тем, что сохранило очертания хвостовых плавников мозазавра, показывая, что Прогнатодон, подобно Платокарп а позже у мозазавров также был двулопастный хвостовой двуусток, напоминающий опущенный хвост акулы, форма которого могла помочь существу всплыть на поверхность, а также атаковать добычу. Открытие также подтверждает теорию о том, что более поздние мозазавры были еще более приспособлены к образу жизни, который сначала занимали ихтиозавры.[9]
19 сентября 2012 года было объявлено, что девятью днями ранее в известняковом карьере недалеко от Маастрихта снова был найден скелет того, что похоже на мозазавра, в том же карьере, где был обнаружен типовой образец Mosasaurus hoffmanni. Карло Брауэр, оператор экскаватора ENCI в карьере, обнаружил зубы окаменелости в лопате своего экскаватора в понедельник утром, 10 сентября. В дни, прошедшие после открытия, сотрудники музея извлекли несколько больших участков черепа, а также часть тела и хвост примерно 13-метрового длинный скелет. На основе стратиграфия возраст экземпляра был оценен в 67,83 миллиона лет, что делает его примерно на полтора миллиона лет старше, чем "Bèr". Судя по тому, что было обнаружено, это, по-видимому, самый старый из известных экземпляров мозазаврид из района Маастрихта, и он принадлежит к роду Прогнатодон.[12] Образец получил прозвище Карло в честь обнаружившего его сотрудника ENCI.[13]
Палеобиология
Палеоэкология

Открытие хорошо сохранившихся образцов Prognathodon overtoni в Кампанский Формация Медвежьей Лапы из Альберта, Канада позволили детально изучить содержимое кишечника (включая фрагменты большой и маленькой рыбы, морская черепаха и потенциально головоногие моллюски ) и зубные ряды, которые позволяли размышлять об экологии Прогнатодон. Как и у большинства мозазавров, зубы этих особей имеют килевидную форму, причем кили расположены примерно параллельно челюсти. На неизношенных зубах верхушка острая, но тупая, с тонкими волнистыми анастомозирующими гребнями, составляющими до 25% высоты коронки. Такой орнамент потенциально мог укрепить зубы. Тупой кончик и шероховатая поверхность позволяют предположить, что зуб использовался для захвата довольно твердой добычи, а также наличие черепаха кости как содержимое кишечника подтверждает гипотезу о том, что Прогнатодон был приспособлен, чтобы сокрушать добычу с твердым панцирем.[2]
Зубы, однако, довольно высокие по сравнению с размером черепа, что говорит о том, что они использовались для пронзания добычи, а не для того, чтобы раздавить или схватить ее. Многие из полностью прорезавшихся зубов имеют зубцы на килях, образующие мелкие зубцы. Наличие зубчатых килей предполагает, что Прогнатодон вместо этого был оппортунистическим хищником, сопоставимым с современным косатки, а не специально приспособлен к сокрушению своей добычи. Такой хищник может питаться не только очень крупной добычей позвоночных, но и множеством других жертв. Тем не мение, P. overtoni у зубов отсутствует заостренный кончик, который в остальном характерен для условно-патогенных хищников с «режущими» зубами. Таким образом, зубы Прогнатодон похоже, показывают приспособления, которые обычно не встречаются вместе.[2]

Стоит отметить, что P. overtoni проявляет неоднозначность, как и другие мозазаврины, Такие как Globidens и Carinodens. Например, передние зубы более изогнутые и тонкие, чем задние, с постепенным изменением формы вдоль зубного ряда. Передние зубы имеют отношение длины коронки к ширине базальной коронки от 2,0 до 2,5, тогда как зубы в середине зубного ряда имеют отношение в диапазоне от 1,7 до 2,0. Эти соотношения соответствуют обоим мозазаврам с «режущими» и «дробящими» зубами. Хотя крепкие, зубы Прогнатодон далеко не такие широкие, как у типичных мозазавров с "сокрушающими" зубами, таких как Globidens.[2]
Самые задние зубы резко изогнуты и короткие, поэтому маловероятно, что они использовались для ловли добычи или обработки пищи. Зубы на верхняя челюсть и зубной Оба исследованных экземпляра имеют значительный износ. Вершины коронки необычно гладкие и отполированные, эта поломка и последующая полировка, вероятно, связаны с длительным контактом с продуктами питания. Поломка зубов несложная и почти горизонтальная, в отличие от типичных хищников с «режущими» зубами. Зубы могли быть достаточно прочными, чтобы предотвратить обширное разрушение, или, возможно, кривизна ограничивала его. Многие зубы изношены равномерно, что предполагает третью возможность; что он представляет собой постепенное стирание зубов в результате контакта с пищей. Несколько похожий износ наблюдается на зубах Globidens schurmanni, известно, что питались иноцерамид двустворчатые моллюски.[2]
Понятно, что износ зубьев не представляет собой простую поломку, поскольку это приведет к разному износу разных зубьев. В отличие от краевых зубов, крыловидные зубы, хотя и необычно большие для мозазавра, не изнашиваются. Это говорит о том, что краевые и крыловидные зубы выполняли разные функции, возможно, крыловидные зубы использовались для захвата добычи перед ее проглатыванием. Большие передние крыловидные зубы, характерные для Прогнатодон вероятно, обеспечивали эффективный захват крупных пищевых продуктов, указывая на то, что этот род был способен глотать добычу большими кусками.[2]
Один из экземпляров Альберты, ТМП 2007.034.0001, является первым Прогнатодон образец с сохраненным содержимым кишечника. В это содержимое входят останки очень большой (1,6 метра) рыбы, более мелкой рыбы, морская черепаха и возможные остатки головоногие моллюски. Эти предметы добычи сильно отличаются друг от друга и обычно являются предметами добычи для разных ниши хищников, но вместе с ними совместимы с крупным и оппортунистическим верхушечным хищником. Хотя они, казалось бы, могли питаться добычей, типичной для мозазавров с «режущими» зубами, например, крупной рыбы, крепкие зубы позволяют предположить, что и более твердую добычу, такую как морская черепаха, также обычно съедали.[2] Prognathodon overtoni, вероятно, схожий по экологии с другими видами этого рода, таким образом, вероятно, был условно-патогенным хищником, способным питаться практически всем, что есть в природе. Западный внутренний морской путь.[2]
Тафономия

Типовой образец Прогнатодон сатуратор сохранились некоторые тафономически интересные детали. Вероятной причиной смерти особи был возраст или болезнь из-за того, что морские хищники были достаточно большими, чтобы убить что-то своего размера, неизвестное из Маастрихтский. Степень сочленения образца предполагает, что животное достигло морского дна через несколько мгновений после смерти, где оно было съедено акулами перед тем, как быть захороненным в отложениях. Доказательства поедания акул включают находки связанных зубов акульих родов. Скваликоракс и Пликатосциллий среди костей мозазавра. Можно исключить возможность того, что мозазавры съели акул до его смерти (и, таким образом, представляли содержимое желудка), поскольку у них нет доказательств нападения со стороны желудочные кислоты. Количество акульих зубов (не говоря уже о постоянном размере и цвете зубов) также слишком велико, чтобы его можно было отнести к фоновому изобилию. На самом скелете также сохранились различные следы укусов, что является прямым доказательством того, что акула собирала падаль.[7]
Патология
А Прогнатодон образец, известный как NHMM 2012 (часто называемый «Карло»), описанный Бастиаансом и другие. в журнале Меловые исследования в 2020 году был обнаружен в Нидерландах недалеко от Маастрихта, и у него были выявлены серьезные лицевые деформации, включая разрушительную частичную ампутацию предчелюстной кости. В Прогнатодона Считается, что травмы являются результатом драки с другим мозазавром (вероятно, другим Прогнатодон).На этом образце видны признаки возобновления роста кости вокруг травмы, а также последующие инфекции, которые продолжались на момент смерти мозазавра, которые, возможно, были частично вызваны нанесенными травмами. Этот экземпляр является одним из немногих четко подтверждаемых случаев внутривидовой борьбы между мозазаврами.[14]
Классификация и виды
Современные филогенетические анализы постоянно Прогнатодон в пределах Mosasaurinae подсемейство, несмотря на это Прогнатодон исторически считался родом, разделяющим тесные отношения с Платокарп и Плиоплаткарпины. Луи Долло был одним из первых исследователей, работавших над систематикой мозазавров, первоначально поместив их в отдельный ящерица подотряд и разделив группу на две семьи, Mosasauridae и «Plioplatecarpidae». В этой ранней таксономии Mosasauridae содержали роды Clidastes, Мозазавр, Платокарп, Халисавр и Тилозавр а Plioplatecarpidae был монотипным, содержал только Плиоплаткарпус. В 1890 году после дальнейших открытий мозазавров (в том числе Прогнатодон), Долло пересмотрел свою таксономию, разделив Mosasauridae на три группы. Эти группы были основаны на том, насколько развит рострум на предчелюстной кости, размеру надстапедиального отростка квадратной кости и сливались ли гемальные дуги с центрами хвостовых позвонков. Прогнатодон был размещен рядом с Платокарп в «микроринхальной» группе. Две другие группы были «мегаринхами» (в том числе Тилозавр и Хайнозавр) и «мезоринхоза» (в том числе Мозазавр и Clidastes) группы.[4]
Долло понял, что Плиоплаткарпус общие персонажи с группой «микроринхов» в 1894 г. и отказались от двух предыдущих семейных систем, начав использовать только одно семейство мозазавров, Mosasauridae, и поместив Прогнатодон как тесно связанный с Платокарп и Плиоплаткарпус.[4]
Рассел (1967) сохранил Прогнатодон в пределах Plioplatecarpinae, но воздвигли трибу для рода и родственных Плезиотилозавр, то Прогнатодонтини. Он считал мозазавров в пределах Prognathodontini «явно происходящими из плиоплаткарпина», но оправдал племя тем, что их можно отличить от других плиоплаткарпинов по их массивным челюстям и крепким зубам.[4]
Горден Л. Белл-младший провел первый крупный филогенетический анализ мозазавров в 1997 году, используя новые методологии и включив другие таксоны, описанные после монографии Рассела 1967 года (особенно базальные мозазавроиды, такие как Эгиалозавр ). Белл выздоровел Прогнатодон среди Mosasaurinae, впервые в истории, как близкий родственник родов Globidens и Плезиотилозавр. Племя Prognathodontini было синонимом Globidensini, еще одно племя, созданное Расселом (1967) для Globidens. Белл также был первым, кто заметил, что его анализ восстановился. Прогнатодон, ранее считалось монофилетический, так как парафилетический.[15]
Взгляд на родство этого рода с другими родами мозазавров мало изменился с 1997 г., он обычно восстанавливается как в пределах Mosasaurinae, так и в парафилетических. Кау и Мадзия (2017) отметили, что включение Прогнатодон и Плезиотилозавр внутри Globidensini предполагает более тесную связь между родами, чем реальность ситуации. Хотя Прогнатодон и Плезиотилозавр обычно восстанавливаются как родственные роды, Cau and Madzia (2017) не воскрешали племя Prognathodontini в своем списке кладов мозазавров и их предпочтительных определениях, не давая никаких комментариев относительно того, почему нет.[16] В рамках анализа Кау и Мадзиа (а также предыдущих работ, таких как Simões et al., 2017[17]), Прогнатодон восстанавливается как родственная группа Mosasaurini, и кладу Мосазаурини + Прогнатодон сам был восстановлен как родственная группа Globidensini.[16]
Результаты всестороннего филогенетического анализа Mosasauridae, проведенного Simões et al. (2017)[17] показан ниже, свернут, чтобы отобразить только Mosasaurinae. Кладограмма показывает Прогнатодон как парафилетический, как пришли к заключению самых последних исследователей, и относит его к сестринской группе производным мозазавринам в Mosasaurini.
| Mosasaurinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Разновидность

Большое количество видов отнесено к Прогнатодон, хотя этот род широко считается парафилетический.[18] Неполный характер Прогнатодон экземпляров привело к неполному пониманию систематики рода в целом, включая как разнообразие альфа-уровня, так и общие характеристики. Более того, крайне редки ранние и средние Кампанский образцы Прогнатодон затмил раннюю часть своей эволюционной истории.[2]
Вопрос о том, что составляет Прогнатодон еще предстоит тщательно изучить. Линдгрен (2005)[18] указали, что крепкие и конические зубные коронки с тупыми, зазубренными килями и гладкой эмалью обычно относятся к роду, несмотря на родовой вид, P. solvayi отклоняется от этого описания, так как демонстрирует заметно сжатые лабио-лингвистические и слегка фасеточные краевые зубы.
Виды считаются действительными и внутри Прогнатодон, как было обнаружено Simões et al. (2017),[17] перечислены ниже. Примечательно, что исключение шести спорных видов значительно сокращает как временной, так и географический диапазон рода, ограничивая его до Европа и Средний Восток и удаление любого вхождения до Позднего Кампанский.
- Прогнатодон солвайи Долло, 1889 - Маастрихтский, Бельгия и Испания.[19]
- P. solvayi это типовой вид из Прогнатодон а также самый маленький из известных видов - всего 5 метров в длину.[3] P. solvayi отличается от всех других представителей рода в нескольких важных аспектах, что необычно для типового вида. Наиболее заметное различие, пожалуй, заключается в зубах, которые мягко огранены и лабиально-язычно сжаты.[18]
- Prognathodon currii Кристиансен и Бонд, 2002 - Поздно Кампанский, Израиль.
- P. currii один из самых крупных, если не самый крупный вид Прогнатодон и известен по единственному массивному черепу. Череп имеет сходство с Прогнатодон, в частности к типовым видам P. solvayi, но отличается от всех известных мозазавров тем, что лоб заметно шире, чем длинна. Хотя изначально неофициально называли как часть своего рода, "Оронозавр", многочисленные филогенетические анализы показали, что он прочно вошел в Прогнатодон.[20]
- Прогнатодон гигантский Долло, 1904 - Маастрихтский, Бельгия, Иордания и Сирия.[21]
- Прогнатодон хашими Каддуми, 2009 - Маастрихтский, Иордания.
- P. hashimi изначально был описан как отдельный род "Тенеразавр", но был переведен на Прогнатодон из-за отсутствия каких-либо истинных родоразделительных признаков. Типовой образец особенно сохраняет мягкие ткани, что является дополнительным доказательством наличия двулопастных хвостовых плавников у мозазавров, подобных некоторым знаменитым образцам Платокарп.[9]
- Prognathodon hudae Каддуми, 2009 - Маастрихтский, Иордания.
- P. hudae это вид среднего размера, который можно отличить от других видов Прогнатодон благодаря почти прямому дорсальному краю и 15 зубам в зубной кости.[3]
- Prognathodon lutugini Яковлев, 1901 - Поздно Кампанский, Украина.
- Хоть и не самый большой, Prognathodon lutugini был большим видом Прогнатодон Примерно 8 метров в длину.[18] Иногда его относят к собственному роду, «Доллозавр» и был взыскан с депозитов Позднего Кампанский возраст в Украина. Окаменелости описываются как "Доллозавр sp. »были извлечены из отложений аналогичного возраста в г. Швеция и Россия,[22] и поэтому может принадлежать P. lutugini также.
- Prognathodon lutugini отличается от всех остальных видов Прогнатодон на его пластинчато-угловой поверхности с отчетливыми горизонтальными языками и бороздками, форме мыщелка позвонка (слегка вдавленный с отношением высоты к ширине 0,75) и пропорциями длины позвонков (шейные позвонки почти равны по размеру самому длинному позвонки колонны). P. lutugini также отличается от всех других видов, кроме P. solvayi имея 13 зубов в его зубной части, хотя может быть исключен из-под утопления в P. solvayi за счет гладкой эмалевой поверхности.[8]
- Отсутствие передней зубной проекции отличает P. lutugini из П. кианда, а также P. lutugini наличие более крупных передних крыловидных зубов. Отсутствие медиальной бороздки на поверхности зубов отличает его от П. waiparaensis и P. solvayi. Кроме того, наличие зубцов килей на зубах отделяет их от П. кианда и P. currii. Форма мыщелка позвонков (не выше их ширины) отделяет его от П. сатуратор и P. rapax.
- P. lutugini первоначально был назван как вид Clidastes Яковлева в 1901 г. на основе типового экземпляра ЦНИГР 818, неполного черепа и скелета. Считая уникальным сочетание функциональных дополнительных сочленений и крупных крыловидных зубов, Яковлев в 1905 году создал новый род, Доллозавр, названный в честь бельгийского палеонтолога Луи Долло.[18] Lingham-Soliar (1989) показал, что эти характеристики не были уникальными для "Доллозавр", который он синонимизировал на уровне рода с Прогнатодон.[5]
- Линдгрен (2005)[18] отметил, что таксономическая ситуация P. lutugini (или же "Доллозавр") было несколько неясным, и что стоматологические и остеологические особенности предполагали размещение внутри Прогнатодон в то время как боковые ямки хвостовых позвонков предполагали более отдаленное родство. Кроме того, споры вокруг того, является ли род Прогнатодон является парафилетический или не сложный, лучше взгляните на классификацию упомянутого материала P. lutugini.
- Филогенетический анализ, проведенный в рамках повторного описания P. lutugini Автор: Д.В. Григорьев (2013)[8] поддержал направление P. lutugini к Прогнатодон и дополнительно предлагается о тесной связи с типовым видом P. solvayi.
Спорные и бывшие виды

Виды Прогнатодон Кианда, P. overtoni, P. rapax, П. сатуратор, P. stadtmani и П. waiparaensis признаны отдельными видами мозазавров, но их обозначение как видов Прогнатодон является спорным, и последние филогенетические анализы, такие как Madzia and Cau (2017)[16] и Simões et al. (2017),[17] восстановить указанный вид как не принадлежащий к роду Прогнатодон, большинство из них, таким образом, потенциально представляют роды сами по себе.
- Прогнатодон Кианда Schulp et al., 2008 - Маастрихтский, Ангола.
- П. кианда известно из Маастрихтский из Ангола и уникален тем, что имеет большое количество краевых зубов и относительно небольшие крыловидные зубы.[23] Этот вид, по-видимому, имеет близкие отношения с сомнительным родом Лиодон и регулярно оказывается за пределами Прогнатодон (и восстановлен как гораздо более базальный мозазаврин ) в большинстве филогенетических анализов.[8]
- Prognathodon overtoni Уиллистон, 1897 - Кампанский, Канада (Альберта) и США (Колорадо и Южная Дакота).[24]
- P. overtoni можно отличить от других видов, в том числе P. solvayi, гладкой эмалью его зубов и их непоколебимой природой.[4]
- Прогнатодон рапакс Хэй, 1902 - Маастрихтский, США (Мэриленд и Нью-Джерси).
- P. rapax легко отделяется от видов P. overtoni и P. solvayi знаками в квадрате и имеет позвоночные черты, очень напоминающие позвонки Мозазавр.[4]
- Прогнатодон сатуратор Дортангс и др., 2002 - Маастрихтский, Нидерланды.
- П. сатуратор массивно построенный и огромный вид из Маастрихтский из Нидерланды. П. сатуратор достигали длины примерно 9-10 метров, возможно, только короче P. overtoni (чья классификация как Прогнатодон также является спорным) и P. currii. Конкретное имя сатуратор является латинский для «тот, кто приносит удовлетворение», имея в виду, что типовой образец был первым достаточно полным образцом мозазавра, извлеченным из Маастрихт Площадь с 1957 года.[7]
- Наиболее очевидная отличительная черта П. сатуратор что отделяет его от других видов Прогнатодон его огромный размер, больше, чем у любого другого вида Прогнатодон известен на момент его описания и в целом является значительно более массивным и надежным. П. сатуратор также отсутствует высокий гребень на спинном срединном гребне, который есть у других представителей рода. Его можно легко отличить от большинства других видов по дальнейшим индивидуальным отличиям. Рано Кампанский P. stadtmani имеет гораздо более тонкую зубную часть, что также относится к П. waiparaensis. P. giganteus можно отличить от П. сатуратор прямым дорсальным краем зубов, маленьким венцом, прямоугольной задней частью нижней челюсти и тупым ретросуставным отростком, а также наличием гораздо менее массивной квадратной кости.[7]
- Типовой образец, NHMM 1998141, сохраняет почти полный череп, сочлененный шейный и передний спинные позвонки с шейными и спинными ребрами, изолированными пигалом и хвостовые позвонки, лопаточно-клювовидный и рассеянный элементы аппендикулярного скелета.[7]
- Прочность квадратной кости наряду с такой же прочной нижней челюстью позволяет предположить, что П. сатуратор мог достичь силы укуса, превышающей все другие мозазавры. По сравнению с квадратами рода Globidens и другие виды Прогнатодон, один из П. сатуратор еще сильнее построен. С известными примерами крупных морских черепах из региона Маастрихт, такими как Аллоплеврон, возможно (на основании найденных заживших следов укусов), что диапазон размеров жертвы П. сатуратор включая этих черепах длиной 2,9 метра.[7]
- Prognathodon waiparaensis Уэллс и Грегг, 1971 - Маастрихтский, Новая Зеландия.
- П. waiparaensis известен частичным скелетом, включая расчлененный череп, шейные позвонки и ребра, извлеченные из отложений маастрихтского возраста в Новая Зеландия.[25]
Три вида сомнительного рода Лиодон (L. сектор, L. compressidens и L. mosasauroides), двое с тонкой морфологией морфологии, были ненадолго переведены в Прогнатодон из-за сходства в соотношении сторон их краевых зубных рядов по краю челюсти к П. кианда.[23] С "Прогнатодон" Кианда многократно восстанавливается как мозазаврин более базальный, чем Прогнатодон[8] и, вероятно, отдельный род, филогенетическое размещение L. сектор, L. compressidens и L. mosasauroides остается крайне неопределенным, хотя они, вероятно, не представляют собой виды Прогнатодон.
Виды P. stadtmani был переописан как типовой вид своего рода, Гнатомортис, в 2020 году.[26]
Рекомендации
- ^ "Fossilworks: Prognathodon". fossilworks.org. Получено 2017-09-26.
- ^ а б c d е ж грамм час я j k л м п о п Кониси, Такуя; Бринкман, Дональд; Massare, Judy A .; Колдуэлл, Майкл У. (01.09.2011). «Новые исключительные образцы Prognathodon overtoni (Squamata, Mosasauridae) из верхнего кампана Альберты, Канада, а также систематика и экология этого рода». Журнал палеонтологии позвоночных. 31 (5): 1026–1046. Дои:10.1080/02724634.2011.601714. ISSN 0272-4634. S2CID 129001212.
- ^ а б c Каддуми, Хани Ф. «Новый вид Prognathodon (Squamata: Mosasauridae) из маастрихта Харрана-Иордания». Цитировать журнал требует
| журнал =(помощь) - ^ а б c d е ж грамм Рассел, Дейл. А. (6 ноября 1967 г.). «Систематика и морфология американских мозазавров» (PDF). Бюллетень музея естественной истории Пибоди (Йельский университет).
- ^ а б c d е ж грамм час я j k Т, Лингхам-Солиар; Д., Нольф (1989). «Мозазавр Prognathodon (Reptilia, Mosasauridae) из верхнего мела Бельгии». Bulletin van Het Koninklijk Belgisch Instituut voor Natuurwetenschappen. Aardwetenschappen = Bulletin de l'Institut Royal des Sciences Naturelles de Belgique. Sciences de la Terre.
- ^ Schulp, Anne S .; Jagt, John W. M .; Фонкен, Франс (10 сентября 2004 г.). «Новый материал мозазавра Carinodens belgicus из верхнего мела в Нидерландах». Журнал палеонтологии позвоночных. 24 (3): 744–747. Дои:10.1671 / 0272-4634 (2004) 024 [0744: NMOTMC] 2.0.CO; 2. ISSN 0272-4634.
- ^ а б c d е ж грамм час я «Большой мозазавр из верхнего мела в Нидерландах (доступен для скачивания PDF)». ResearchGate. Получено 2017-09-26.
- ^ а б c d е ж грамм час я Григорьев, Д. В. (2013). "Повторное описание Prognathodon lutugini (Squamata, Mosasauridae) " (PDF). Труды Зоологического института РАН.. 317 (3): 246–261.
- ^ а б c d е ж грамм час Линдгрен, Йохан; Каддуми, Хани Ф .; Полсин, Майкл Дж. (10.09.2013). «Сохранение мягких тканей ископаемой морской ящерицы с двулопастным хвостовым плавником». Nature Communications. 4: ncomms3423. Bibcode:2013 НатКо ... 4.2423L. Дои:10.1038 / ncomms3423. PMID 24022259.
- ^ Lindgren, J .; Caldwell, M.W .; Кониши, Т .; Чиаппе, Л.М. (2010). Фарке, Эндрю Аллен (ред.). «Конвергентная эволюция водных четвероногих животных: выводы из исключительного ископаемого мозазавра». PLOS ONE. 5 (8): e11998. Bibcode:2010PLoSO ... 511998L. Дои:10.1371 / journal.pone.0011998. ЧВК 2918493. PMID 20711249.
- ^ Christiansen, P .; Бонд, Н. (2002). «Новый вид гигантского мозазавра из позднего мела Израиля». Журнал палеонтологии позвоночных. 22 (3): 629. Дои:10.1671 / 0272-4634 (2002) 022 [0629: ANSOGM] 2.0.CO; 2.
- ^ Патологическая морда мозазавра из типового маастрихта (юго-восток Нидерландов), Bastiaans et al, 2014.
- ^ "Пресс-релиз Natuurhistorisch Museum Maastricht". 20 сентября 2012. Архивировано с оригинал 23 сентября 2012 г.. Получено 20 сентября 2012.
- ^ Бастиан, Дилан; Kroll, Jeroen J.F .; Корнелиссен, Дирк; Schulp, Anne S .; Ягт, Джон В. (Февраль 2020 г.). «Черепная палеопатология позднемелового мозазавра из Нидерландов». Меловые исследования. 112: 104425. Дои:10.1016 / j.cretres.2020.104425.
- ^ Bell GL. Jr. 1997. Филогенетический пересмотр Mosasauroida в Северной Америке и Адриатике. стр. 293-332. В: Callaway JM, Nicholls EL, (eds.), Древние морские рептилии, Academic Press, 501 стр.
- ^ а б c Madzia, D .; Кау А. (2017). «Выявление« слабых мест »в филогенетических деревьях: применение к номенклатуре мозазавроидов» (PDF). PeerJ. 5: e3782. Дои:10.7717 / peerj.3782. ЧВК 5602675. PMID 28929018.
- ^ а б c d Simões, Tiago R .; Вернигора, Оксана; Папарелла, Илария; Хименес-Уидобро, Паулина; Колдуэлл, Майкл У. (2017-05-03). «Филогения мозазавроидов с использованием нескольких филогенетических методов дает новое понимание эволюции водных адаптаций в группе». PLOS ONE. 12 (5): e0176773. Bibcode:2017PLoSO..1276773S. Дои:10.1371 / journal.pone.0176773. ISSN 1932-6203. ЧВК 5415187. PMID 28467456.
- ^ а б c d е ж Линдгрен, Йохан (2005). «Стоматологическая и позвоночная морфология загадочного мозазавра Dollosaurus (Reptilia, Mosasauridae) из нижнего кампана (верхний мел) на юге Швеции». Бюллетень геологического общества Дании. 52 (1): 17–25. ISSN 0011-6297.
- ^ "Fossilworks: Prognathodon solvayi". fossilworks.org. Получено 2017-09-26.
- ^ Кристиансен, Пер; Бонд, Нильс (2002). «Новый вид гигантского мозазавра из позднего мела Израиля». Журнал палеонтологии позвоночных. 22 (3): 629–644. Дои:10.1671 / 0272-4634 (2002) 022 [0629: ansogm] 2.0.co; 2. JSTOR 4524254.
- ^ "Окаменелости: Prognathodon giganteus". fossilworks.org. Получено 2017-09-26.
- ^ Аверьянов, А.О .; Ярков, А. А. (2004). «О появлении гигантской летающей рептилии (Pterosauria) в конце позднего мела Нижнего Поволжья». Палеонтологический журнал. 38 (6): 669–671.
- ^ а б Schulp, Anne S .; Polcyn, Майкл Дж .; Матеуш, Октавио; Джейкобс, Луи Л .; Мораис, Мария Луиза (2008). «Новый вид Prognathodon (Squamata, Mosasauridae) из маастрихта Анголы и сходство с родом мозазавров Liodon». Труды второй встречи мозазавров.
- ^ "Fossilworks: Prognathodon overtoni". fossilworks.org. Получено 2017-09-26.
- ^ "Fossilworks: Prognathodon waiparaensis". fossilworks.org. Получено 2017-10-11.
- ^ Лайвли, Дж. Р. (2020). «Переописание и филогенетическая оценка 'Прогнатодон' Stadtmani: значение для монофилии Globidensini и гомологии характера у Mosasaurinae ". Журнал палеонтологии позвоночных: e1784183. Дои:10.1080/02724634.2020.1784183.
| Mosasaurinae |
|  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Halisaurinae | |||||||||||
| Расселлозаврина |
| ||||||||||
| Родственные группы и роды | |||||||||||
| Статьи по Теме | |||||||||||