Сквален-гопенциклаза - Squalene-hopene cyclase
| Сквален-гопенциклаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
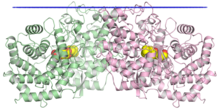 В кристаллографический структура димера сквален-гопенциклазы, при этом положение мембраны обозначено синим, два мономера - зеленым и розовым, а миметик субстрата в центральной полости - желтым.[1] | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 5.4.99.17 | ||||||||
| Количество CAS | 76600-69-6 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
Сквален-гопенциклаза (SHC) (ЕС 5.4.99.17 ) или же гопан-22-ол гидролиза прокариотический фермент в терпен семейство циклазы / мутазы. Он катализирует взаимное превращение ациклической молекулы сквалена в пентациклический тритерпен гопен и гопанол в соотношении 5: 1.[2][3][4][5][6] Этот фермент катализирует следующие химические реакции.
- сквален хмель-22 (29) -ен

- сквален + H2О гопан-22-ол
Сквален-гопенциклаза важна, потому что ее продукты, гопеноиды, очень похожи на стерины у эукариот в том, что они конденсируют липидные мембраны и снижают проницаемость. В случае прокариот они обеспечивают стабильность при высоких температурах и экстремальной кислотности благодаря жестким кольцевым структурам.[7] Действительно, повышенная регуляция сквален-гопенциклазы происходит у некоторых бактерий в присутствии горячей или кислой среды.[8][9]

Вступление
Сквален-гопенциклаза обнаруживается в большом количестве бактерий, но наиболее легко выделяется из термофильных бактерий. Alicyclobacillus acidocaldarius.[10] Он катализирует превращение ациклической молекулы сквален в пентациклические тритерпены Hopene и гопанол.

Считается, что сквален-гопенциклаза является эволюционным предшественником многих классов эукариотических и прокариотических организмов. стерол циклазы.[6] Оксидоскваленциклазы, которые являются эукариотическими аналогами SHC, для их реакции требуется кислород, демонстрируя гораздо более позднюю эволюцию, когда атмосфера начала накапливать кислород. Сквален-гопенциклаза функционирует в гипоксической среде, что предполагает гораздо более раннее ее присутствие.[11]
Структура
Сквален-гопенциклаза представляет собой связанный с мембраной белок 70-75 кДа, состоящий из 631 аминокислоты и семи повторов PTFB. Он существует как монотопный гомодимер.[1]
Механизм
Формирование скелета гопена - одна из самых сложных одностадийных реакций в биохимии.[12] За один этап разрывается или образуется 13 ковалентных связей, устанавливается 9 хиральных центров и образуется 5 колец.[13] Сквален-гопенциклаза катализирует превращение ациклической молекулы сквалена в пентациклические тритерпены гопена и гопанола. Эти продукты представлены в соотношении 5: 1. Синтез гопена начинается со связывания сквалена во всей конформации до кресла и сопровождается образованием пяти связей C-C.[14] Эти последовательные стадии реакции образования кольца инициируются электрофильной атакой кислотного протона на одну из двух концевых двойных связей. Полициклическое образование завершается, когда протон отщепляется от альтернативной концевой метильной группы сквалена посредством акцепции молекулой воды.[5] Эта база известна как передняя вода. Другие молекулы воды работают на усиление поляризации (задние воды) и построить водородные связи между семью остатками - T41, E45, E93, R127, Q262, W133 и Y267. Передняя вода также играет роль в определении конечного продукта. Если он хранит протон, генерируемый либо из метильной группы 29, либо из 30 с образованием гопена. Однако гопанол производится в меньших количествах, если вместо того, чтобы принимать протон, вода вносит гидроксил в катион C-22 A-кольца.[15]

Во время образования колец от A до D конформационные изменения очень незначительны. Следовательно, реакция не требует промежуточного соединения и может протекать в одну стадию. Однако образованию кольца E препятствует энтропийный барьер, который может объяснить его отсутствие в тетрациклическом стероиды.[5]
Активный сайт
SHC активный сайт расположен в центральной полости в области белка, прилегающей к мембране, и к нему получает доступ субстрат через неполярный канал.[16] Активный центр особенно окружен ароматическими остатками, образующими полость, которая удобно прилегает к молекуле сквалена, когда она свернута в продуктивную конформацию. В каталитический механизм использует связанные аспартат и гистидин остатки, чтобы инициировать циклизация реакция со стороны протонирующий в C3 и депротонирует в C29, проходя дискретную серию карбокатион промежуточные звенья.[1][17] Фермент может быть инактивирован мутацией каталитических аспартатов.[18]
Термодинамика
Этот фермент необычно экзотермический с выделением энергии 40-50 ккал / моль, что значительно превышает энергию стабилизации белка. Считается, что это плавит боковой липидный канал, через который выходит объемный продукт. Некоторые ученые считают, что для сохранения структурной целостности фермента 7-8 нетандемных повторяющихся мотивов QW (Q - глутамин и W есть триптофан ), которые соединяют многочисленные поверхности α спирали укрепить структуру белка и предотвратить денатурацию.[1]

Рекомендации
- ^ а б c d Wendt KU, Poralla K, Schulz GE (сентябрь 1997 г.). «Структура и функция скваленциклазы». Наука. 277 (5333): 1811–5. Дои:10.1126 / science.277.5333.1811. PMID 9295270.
- ^ Хосино Т., Сато Т. (февраль 2002 г.). «Сквален-гопенциклаза: каталитический механизм и распознавание субстрата». Химические коммуникации (4): 291–301. Дои:10.1039 / b108995c. PMID 12120044.
- ^ Хосино Т., Накано С., Кондо Т., Сато Т., Миёси А. (май 2004 г.). «Сквален-гопенциклаза: окончательная реакция депротонирования, конформационный анализ циклизации (3R, S) -2,3-оксидосквалена и дополнительные доказательства потребности в изопропилиденовой части как для инициирования каскада полициклизации, так и для образования 5-членное Е-кольцо ». Органическая и биомолекулярная химия. 2 (10): 1456–70. Дои:10.1039 / b401172d. PMID 15136801.
- ^ Сато Т., Куда М., Хосино Т. (март 2004 г.). «Эксперименты по сайт-направленному мутагенезу на предполагаемом сайте депротонирования сквален-гопенциклазы из Alicyclobacillus acidocaldarius». Биология, биотехнология и биохимия. 68 (3): 728–38. Дои:10.1271 / bbb.68.728. PMID 15056909.
- ^ а б c Reinert DJ, Balliano G, Schulz GE (январь 2004 г.). «Превращение сквалена в пентакарбоциклический гопен». Химия и биология. 11 (1): 121–6. Дои:10.1016 / j.chembiol.2003.12.013. PMID 15113001.
- ^ а б Пирсон А., Будин М., Брокс Дж. Дж. (Декабрь 2003 г.). «Филогенетические и биохимические доказательства синтеза стеролов у бактерии Gemmata obscuriglobus». Труды Национальной академии наук Соединенных Штатов Америки. 100 (26): 15352–7. Дои:10.1073 / pnas.2536559100. ЧВК 307571. PMID 14660793.
- ^ Kannenberg, E .; Поралла, К. (1999). «Биосинтез и функция гопаноидов в бактериях». Naturwissenschaften. 86 (4): 168–176. Дои:10.1007 / s001140050592.
- ^ Уриссон Г., Ромер М., Поралла К. (1987). «Прокариотические гопаноиды и другие суррогаты политерпеноидных стеролов». Ежегодный обзор микробиологии. 41: 301–33. Дои:10.1146 / annurev.mi.41.100187.001505. PMID 3120639.
- ^ Sahm H, Rohmer M, Bringer-Meyer S, Sprenger GA, Welle R (1993). «Биохимия и физиология гопаноидов у бактерий». Успехи микробной физиологии. 35: 247–73. Дои:10.1016 / с0065-2911 (08) 60100-9. ISBN 9780120277353. PMID 8310881.
- ^ Seckler, B .; Поралла, К. (1986). «Характеристика и частичная очистка сквален-гопенциклазы из Bacillus acidocaldarius». Biochimica et Biophysica Acta (BBA) - Общие предметы. 881 (3): 356–363. Дои:10.1016/0304-4165(86)90027-9.
- ^ Rohmer, M .; Бувье, G .; Уриссон, Г. (1979). «Молекулярная эволюция биомембран: структурные эквиваленты и филогенетические предшественники стеринов». Труды Национальной академии наук. 76 (2): 847–851. Дои:10.1073 / pnas.76.2.847. ЧВК 383070. PMID 284408.
- ^ Зиденбург Г., Ендроссек Д. (июнь 2011 г.). «Сквален-гопенциклазы». Прикладная и экологическая микробиология. 77 (12): 3905–15. Дои:10.1128 / aem.00300-11. ЧВК 3131620. PMID 21531832.
- ^ Кори Э.Дж., Мацуда С.П., Бартель Б. (декабрь 1993 г.). «Выделение гена Arabidopsis thaliana, кодирующего циклоартенолсинтазу, путем функциональной экспрессии в дрожжевом мутанте, лишенном ланостеринсинтазы, с помощью хроматографического скрининга». Труды Национальной академии наук Соединенных Штатов Америки. 90 (24): 11628–32. Дои:10.1073 / пнас.90.24.11628. ЧВК 48037. PMID 7505443.
- ^ Чжэн Ю.Ф., Абэ И., Прествич Г.Д. (апрель 1998 г.). «Кинетика ингибирования и аффинное мечение бактериального сквалена: гопенциклазы тиа-замещенными аналогами 2,3-оксидосквалена». Биохимия. 37 (17): 5981–7. Дои:10.1021 / bi9727343. PMID 9558334.
- ^ [1]
- ^ Гао Ю., Хонзатко РБ, Петерс Р.Дж. (октябрь 2012 г.). «Структуры терпеноидсинтазы: пока неполный взгляд на комплексный катализ». Отчеты о натуральных продуктах. 29 (10): 1153–75. Дои:10.1039 / C2NP20059G. ЧВК 3448952. PMID 22907771.
- ^ Хосино Т., Сато Т. (февраль 2002 г.). «Сквален-гопенциклаза: каталитический механизм и распознавание субстрата». Химические коммуникации (4): 291–301. Дои:10.1039 / B108995C. PMID 12120044.
- ^ Feil, C .; Sussmuth, R .; Jung, G .; и другие. (1996). «Сайт-направленный мутагенез предполагаемых остатков активного сайта в сквален-гопенциклазе». Европейский журнал биохимии. 242 (1): 51–55. Дои:10.1111 / j.1432-1033.1996.0051r.x. PMID 8954152.
внешняя ссылка
- Сквален-гопен + циклаза в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)