Бабакотия - Википедия - Babakotia
| Бабакотия | |
|---|---|
 | |
| Восстановление жизни | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Strepsirrhini |
| Семья: | †Палеопропитеки |
| Род: | †Бабакотия Годфри и другие., 1990[2] |
| Разновидность: | †Б. радофилай |
| Биномиальное имя | |
| †Бабакотия радофилай Годфри и другие., 1990[1] | |
 | |
| Субфоссильные участки дляБабакотия радофилай[3] | |
Бабакотия вымерший род среднего размера лемур, или же Strepsirrhine примат, из Мадагаскар который содержит один вид, Бабакотия радофилай. Вместе с Палеопропитек, Археоиндрис, и Мезопропитек, он образует семейство Palaeopropithecidae, широко известное как ленивые лемуры. Название Бабакотия исходит из Малагасийский имя для Индри, Бабакото, с которым он и все остальные лемуры-ленивцы тесно связаны. Благодаря сочетанию морфологические признаки которые показывают промежуточные стадии между медленно передвигающимися мелкими лемурами-ленивцами и поддерживающими большими лемурами-ленивцами, это помогло определить взаимосвязь между обеими группами и близкородственными и вымершими обезьяны лемуры.
Бабакотия радофилай и все остальные лемуры-ленивцы имеют много общих черт с живыми ленивцы, демонстрируя конвергентная эволюция. У него были длинные предплечья, изогнутые пальцы и очень подвижные тазобедренные и голеностопные суставы. Его череп был более крепким, чем у индрииды, но не так сильно, как у более крупных лемуров-ленивцев. Его зубной ряд подобен таковому у всех других индриидов и лемуров-ленивцев. Он жил в северной части Мадагаскара и разделял ареал по крайней мере с двумя другими видами лемуров-ленивцев, Palaeopropithecus ingens и Mesopropithecus dolichobrachion. Бабакотия радофилай был в первую очередь листоедом (фоливор ), хотя он также ел фрукты и твердые семена. Это известно только из окаменелость остается и, возможно, вымерли вскоре после прибытия людей на остров, но этого недостаточно. радиоуглеродное датирование было сделано с этим видом, чтобы знать наверняка.
Этимология
Название рода Бабакотия происходит от Малагасийский общее название для Индри, Бабакото, близкий родственник Бабакотия. В название вида, радофилай, был выбран в честь французского математика и экспатриант Жан Радофилао, заядлый спелеолог кто нанес на карту пещеры, где останки Бабакотия радофилай были впервые найдены.[4]
Классификация и филогения
Бабакотия радофилай является единственным членом род Бабакотия и принадлежит семья Палеопропитеки, который включает три других рода ленивые лемуры: Палеопропитек, Археоиндрис, и Мезопропитек. Эта семья в свою очередь принадлежит к инфраотряд Лемурообразные, который включает всех малагасийских лемуров.[5][1]
Первые окаменелые останки Бабакотия радофилай были обнаружены в рамках серии экспедиций, последовавших за открытиями Жана Радофилао и двух англо-малагасийских разведывательных экспедиций в 1981 и 1986–1987 гг.[6][7] Вторая волна исследований была начата в 1980-х гг. биологический антрополог Элвин Л. Саймонс[8] который раскопал в 1988 году в пещере, известной как Анцироандоха в Массив Анкарана Северный Мадагаскар представляет собой почти полный скелет и череп в дополнение к останкам примерно дюжины других людей.[3] Сразу после обнаружения был идентифицирован как лемур-ленивец (палеопропитецид).[4] Бабакотия вместе с Мезопропитек помог разрешить спор об отношениях между лемурами-ленивцами, обезьяны лемуры (семейство Archaeolemuridae) и живущие индрииды. У обезьяньих лемуров были черепа, которые больше напоминали индриид, но их зубы были очень специализированными и не похожи на зубы индриидов. С другой стороны, более крупные лемуры-ленивцы сохранили зубной ряд похожи на живых индриидов, но отличались более крепкими и специализированными черепами. Бабакотия и Мезопропитек не только разделяли зубы индриидов, но и индриидоподобные черепа, что свидетельствует о том, что лемуры-ленивцы были наиболее тесно связаны с живыми индриидами, а лемуры-обезьяны были сестринская группа как для.[9] Кроме того, открытие Бабакотия помогли продемонстрировать, что предки индриидов не были «рикошетными прыгунами» (быстро подпрыгивая с дерева на дерево), как живые индрииды, а вертикальными лазаньями и висячими кормушками и, возможно, случайными прыгунами.[10]
Анатомия и психология
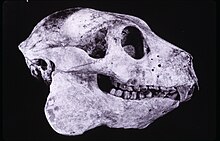
Весом от 16 до 20 кг (от 35 до 44 фунтов), Бабакотия радофилай был лемуром средних размеров и заметно меньше крупных лемуров-ленивцев (Археоиндрис и Палеопропитек), но крупнее мелких лемуров-ленивцев (Мезопропитек).[5][1] Во многих отношениях он имел промежуточный уровень адаптации к сдерживающее поведение между большими лемурами ленивца и маленькими лемурами ленивца.[1][3][11][9][12] Сюда входят его высокомобильные тазобедренные и голеностопные суставы, а также другие специалисты в области позвоночника, таза и конечностей.[12] Его передние конечности были на 20% длиннее, чем Задние конечности, повышая его межмембральный индекс (~ 119) чем Мезопропитек (От ~ 97 до 113), предполагая, что это было конвергентно похожий на древесный ленивцы.[12][13] Было уменьшено предплюсна и изогнутые, удлиненные пальцы, приспособленные для захвата и подсказки поддерживающего поведения.[12][14] Его задние лапы были уменьшены, что делало его хорошо приспособленным для лазания и висения (как у других палеопропитецидов), но не для прыжков (как у индриидов).[5][1][3] Кости запястья, найденные в 1999 году, еще раз продемонстрировали, что этот вид может лазать по высоте.[15] Кроме того, анализ его полукружные каналы,[16] поясничных позвонков и его остистые отростки указывают на медленные движения и приспособления к лазанию (антипроноградные), но не обязательно на ленивое висение, вертикальное цепляние или прыжки.[17] Следовательно, скорее всего, это был медленный альпинист, похожий на Лорис а также проявлял некоторое сдерживающее поведение, как ленивец.[8]
| Бабакотия размещение в филогении лемуров[18][19][11] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Все лемуры-ленивцы имеют относительно крепкие черепа по сравнению с индриидами.[1] но несмотря на то, что поделился черепной особенности с более крупными лемурами-ленивцами,[9] его череп все еще напоминает череп индри.[5] Черты черепа, общие с другими лемурами-ленивцами, включают относительно небольшие орбиты, крепкий скуловые дуги, и в основном прямоугольный твердое небо.[1] Принимаемые во внимание малые орбиты с учетом относительного размера зрительный канал предположить, что Бабакотия был низкий Острота зрения, что характерно для лемуров.[20] Длина черепа в среднем составляет 144 мм (5,7 дюйма).[3]
В зубная формула из Бабакотия радофилай был таким же, как и другие лемуры и индрииды-ленивцы: либо 2.1.2.31.1.2.3[1][9] или же 2.1.2.32.0.2.3 × 2 = 30.[3] Неясно, был ли один из зубов в постоянный прикус является резец или же собачий, в результате чего эти две стоматологические формулы противоречат друг другу.[21] В любом случае, отсутствие нижнего клыка или резца приводит к получению четырехзубого зуба. частый гребень вместо более типичного стрепсеринового гребня с шестью зубцами. Бабакотия радофилай несколько отличался от индриид несколько удлиненной премоляры. Его щечные зубы имели широкие срезанные гребни и зубчатые эмаль.[3]
Распространение и экология
Как и все лемуры, Бабакотия радофилай был эндемичный на Мадагаскар. Его останки были найдены только в известняковые пещеры на массиве Анкарана в пределах Анкарана заповедник и в Anjohibe, что указывает на диапазон на крайнем севере и северо-западе острова.[1][3][22] Ограниченный ареал этого древесного приматы, особенно в то время, когда большая часть острова была покрыта лесом, могло быть связано со спецификой среды обитания, конкурентное исключение, или какой-то другой неизвестный фактор.[23] Это было сочувствующий (произошло вместе) с Palaeopropithecus maximus и Mesopropithecus dolichobrachion.[8][24][25]
Исходя из его размера, морфология его коренные зубы, и анализ микроизоляций на его зубах, Бабакотия радофилай вероятно был фоливор, дополняя свой рацион фруктами и твердыми семена.[5][11][20][26] У всех лемуров-ленивцев, включая Бабакотия радофилай, то постоянные зубы извергались рано - признак, наблюдаемый у индриидов, который улучшает выживаемость молоди в течение первого сухого сезона после отлучение от груди.[12]
Вымирание
Поскольку он вымер относительно недавно и известен только по окаменелым останкам, он считается современной формой малагасийского лемура.[22] Бабакотия радофилай жил во время Голоцен эпоха и считается, что он исчез вскоре после прибытия людей на остров, возможно, в течение последних 1000 лет.[1] Единственный радиоуглеродная дата сообщается, что это относится к 3100–2800 гг. до н. э.[27]
Рекомендации
- ^ а б c d е ж грамм час я j Новак, Р. (1999). "Семья Palaeopropithecidae: ленивые лемуры". Млекопитающие мира Уокера (6-е изд.). Издательство Университета Джона Хопкинса. стр.89–91. ISBN 978-0-8018-5789-8.
- ^ McKenna, M.C .; Белл, С. (1997). Классификация млекопитающих: выше уровня вида. Издательство Колумбийского университета. п. 335. ISBN 978-0-231-11013-6.
- ^ а б c d е ж грамм час Годфри, Л.Р .; Юнгерс, W.L. (2002). «Глава 7: Четвертичные ископаемые лемуры». В Hartwig, W.C. (ред.). Летопись окаменелостей примаса. Издательство Кембриджского университета. С. 97–121. ISBN 978-0-521-66315-1.
- ^ а б Годфри, Л.Р .; Simons, E.L .; Chatrath, P.J .; Ракотосамиманана Б. (1990). «Новый ископаемый лемур (Бабакотия, Приматы) с северного Мадагаскара ". Comptes Rendus de l'Académie des Sciences. 2. 81: 81–87.
- ^ а б c d е Миттермайер, Р.А.; Konstant, W.R .; Hawkins, F .; Louis, E.E .; Langrand, O .; Ratsimbazafy, J .; Rasoloarison, R .; Ganzhorn, J.U .; Rajaobelina, S .; Tattersall, I .; Мейерс, Д. (2006). «Глава 3: Вымершие лемуры». Лемуры Мадагаскара. В иллюстрациях С.Д. Нэш (2-е изд.). Консервейшн Интернэшнл. С. 37–51. ISBN 978-1-881173-88-5.
- ^ Wilson, J.M .; Стюарт, П.Д .; Ramangason, G.-S .; Деннинг, A.M .; Хатчингс, М. (1989). «Экология и сохранение коронованного лемура в Анкаране, Северный Мадагаскар, с примечаниями о лемуре Сэнфорда, других симпатриях и субфоссильных лемурах». Folia Primatologica. 52 (1–2): 1–26. Дои:10.1159/000156379. PMID 2807091.
- ^ Wilson, J.M .; Годри, Л.Р .; Simons, E.L .; Стюарт, П.Д .; Вийом-Рандриамантена, М. (1995). «Прошлая и настоящая фауна лемуров в Анкаране, Северный Мадагаскар» (PDF). Примат сохранения. 16: 47–52.
- ^ а б c Годфри, Л.Р .; Юнгерс, W.L. (2003). "Вымершие лемуры-ленивцы Мадагаскара" (PDF). Эволюционная антропология. 12 (6): 252–263. Дои:10.1002 / evan.10123. S2CID 4834725.
- ^ а б c d Миттермайер, Р.А.; Таттерсолл, И.; Konstant, W.R .; Meyers, D.M .; Маст, Р. Б. (1994). «Глава 4: Вымершие лемуры». Лемуры Мадагаскара. В иллюстрациях С.Д. Нэш (1-е изд.). Консервейшн Интернэшнл. С. 33–48. ISBN 978-1-881173-08-3.
- ^ Jungers, W.L .; Годфри, Л.Р .; Simons, E.L .; Chatrath, P.S .; Ракотосамиманана Б. (1991). «Филогенетическое и функциональное сродство Бабакотия (Приматы), ископаемый лемур с севера Мадагаскара " (PDF). Труды Национальной академии наук. 88 (20): 9082–9086. Bibcode:1991PNAS ... 88.9082J. Дои:10.1073 / пнас.88.20.9082. ЧВК 52656. PMID 1924371.
- ^ а б c Годфри, Л.Р .; Юнгерс, W.L. (2003). «Субфоссильные лемуры». В Goodman, S.M .; Бенстед, Дж. П. (ред.). Естественная история Мадагаскара. Издательство Чикагского университета. С. 1247–1252. ISBN 978-0-226-30306-2.
- ^ а б c d е Саймонс, Э. (1997). «Глава 6: Лемуры: старое и новое». В Goodman, S.M .; Паттерсон, Б.Д. (ред.). Природные изменения и антропогенное воздействие на Мадагаскаре. Пресса Смитсоновского института. С. 142–166. ISBN 978-1-56098-682-9.
- ^ Годфри, Л.Р .; Jungers, W.L .; Reed, K.E .; Simons, E.L .; Чатрат, П.С. (1997). «Глава 8: Субфоссильные лемуры». В Goodman, S.M .; Паттерсон, Б.Д. (ред.). Природные изменения и антропогенное воздействие на Мадагаскаре. Пресса Смитсоновского института. С. 218–256. ISBN 978-1-56098-682-9.
- ^ Jungers, W.L .; Годфри, Л.Р .; Simons, E.L .; Чатрат, П.С. (1997). «Кривизна фаланга и позиционное поведение у вымерших лемуров-ленивцев (приматы, Palaeopropithecidae)» (PDF). Труды Национальной академии наук. 94 (22): 11998–12001. Bibcode:1997PNAS ... 9411998J. Дои:10.1073 / пнас.94.22.11998. ЧВК 23681. PMID 11038588.
- ^ Hamrick, M.W .; Simons, E.L .; Юнгерс, W.L. (2000). «Новые кости запястья малагасийских гигантских окаменелых лемуров». Журнал эволюции человека. 38 (5): 635–650. Дои:10.1006 / jhev.1999.0372. PMID 10799257.
- ^ Уокер, А .; Ryan, T.M .; Silcox, M.T .; Simons, E.L .; Спур, Ф. (2008). "Система полукружных каналов и движение: случай вымерших лемуроидов и лорисоидов". Эволюционная антропология. 17 (3): 135–145. Дои:10.1002 / evan.20165. S2CID 83737480.
- ^ Shapiro, L.J .; Seiffert, C.V.M .; Годфри, Л.Р .; Jungers, W.L .; Simons, E.L .; Randria, G.F.N. (2005). «Морфометрический анализ поясничных позвонков вымерших малагасийских Strepsirrhines». Американский журнал физической антропологии. 128 (4): 823–839. Дои:10.1002 / ajpa.20122. PMID 16110476.
- ^ Horvath, J.E .; Weisrock, D.W .; Embry, S.L .; Fiorentino, I .; Balhoff, J.P .; Kappeler, P .; Wray, G.A .; Willard, H.F .; Йодер, А.Д. (2008). «Разработка и применение филогеномного инструментария: решение эволюционной истории лемуров Мадагаскара» (PDF). Геномные исследования. 18 (3): 489–499. Дои:10.1101 / гр.7265208. ЧВК 2259113. PMID 18245770. Получено 24 февраля 2010.
- ^ Орландо, Л .; Calvignac, S .; Schnebelen, C .; Douady, C.J .; Годфри, Л.Р .; Хэнни, К. (2008). «ДНК вымерших гигантских лемуров связывает археолемурид с современными индриидами». BMC Эволюционная биология. 8: 121. Дои:10.1186/1471-2148-8-121. ЧВК 2386821. PMID 18442367.
- ^ а б Годфри, Л.Р .; Jungers, W.L .; Шварц, Г. (2006). «Глава 3: Экология и исчезновение субфоссильных лемуров Мадагаскара». In Gould, L .; Sauther, M.L (ред.). Лемуры: экология и адаптация. Springer. С. 41–64. ISBN 978-0-387-34585-7.
- ^ Анкель-Саймонс, Ф. (2007). «Глава 7: Зубы». Анатомия приматов (3-е изд.). Академическая пресса. С. 224–283. ISBN 978-0-12-372576-9.
- ^ а б Суссман, Р.В. (2003). «Глава 4: Ночные лемурообразные». Экология и социальная структура приматов. Пирсон Custom Publishing. С. 107–148. ISBN 978-0-536-74363-3.
- ^ Burney, D.A .; James, H.F .; Grady, F.V .; Rafamantanantsoa, J .; Рамилисонина; Wright, H.T .; Коварт, Дж. Б. (1997). «Изменение окружающей среды, исчезновение и деятельность человека: свидетельства из пещер на северо-западе Мадагаскара». Журнал биогеографии. 24 (6): 755–767. Дои:10.1046 / j.1365-2699.1997.00146.x. HDL:2027.42/75139. JSTOR 2846113.
- ^ Годфри, Л.Р .; Уилсон, Джейн М .; Simons, E.L .; Стюарт, Пол Д .; Вийом-Рандриамантена, М. (1996). «Анкарана: окно в прошлое Мадагаскара». Лемуры Новости. 2: 16–17.
- ^ Уилсон, Джейн М .; Годфри, Л.Р .; Simons, E.L .; Стюарт, Пол Д .; Вийом-Рандриамантена, М. (1995). «Прошлая и настоящая фауна лемуров в Анкаране, Северный Мадагаскар». Примат сохранения. 16: 47–52.
- ^ Rafferty, K.L .; Тефорд, М.Ф .; Юнгерс, W.L. (2002). «Молярные микрокожи субфоссильных лемуров: улучшение разрешения диетических выводов». Журнал эволюции человека. 43 (5): 645–657. Дои:10.1006 / jhev.2002.0592. PMID 12457853.
- ^ Simons, E.L .; Burney, D.A .; Chatrath, P.S .; Годфри, Л.Р .; Jungers, W.L .; Ракотосамиманана, Б. (1995). "AMS 14C Даты вымерших лемуров из пещер в массиве Анкарана, Северный Мадагаскар ". Четвертичное исследование. 43 (2): 249–254. Bibcode:1995QuRes..43..249S. Дои:10.1006 / qres.1995.1025.