Биогенный кремнезем - Википедия - Biogenic silica
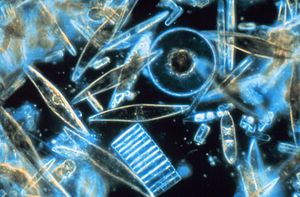
Биогенный кремнезем (bSi), также называемый опал, биогенный опал, или аморфный опаловый кремнезем, образует один из самых распространенных биогенных минералы. Например, микроскопические частицы кремнезема, называемые фитолиты можно найти в травах и других растениях. Кремнезем - аморфный металл окись сформированный комплексом неорганический полимеризация процессы. Это отличается от других основных биогенный минералы, включающие карбонат и фосфат, которые встречаются в природе в виде кристаллических ионо-ковалентных твердых веществ (например, соли ) чей осадки продиктовано равновесие растворимости.[1] Химически bSi гидратированный кремнезем (SiO2·пЧАС2O), который необходим для многих растений и животных.
Кремнезем в морской среде
Силикат, или же кремниевая кислота (ЧАС4SiO4), является важным питательное вещество В океане. В отличие от других основных питательных веществ, таких как фосфат, нитрат, или же аммоний, которые нужны практически всем морским планктон, силикат - необходимое химическое вещество для очень специфических биота, включая диатомеи, радиолярии, силикофлагеллаты, и кремнистые губки. Эти организмы извлекают растворенный силикат из поверхностных вод открытого океана для накопления твердого кремнезема (SiO2) или опаловые скелетные структуры (т. е. твердые части биоты).[2][3] Некоторые из наиболее распространенных кремнистых структур, наблюдаемых на поверхности клеток организмов, выделяющих кремнезем, включают: спикулы, чешуйки, твердые пластинки, гранулы, створки и другие сложные геометрические формы, в зависимости от рассматриваемых видов.[4]
Пять основных источников растворенный Кремнезем в морской среде можно выделить:[3]
- Речной приток растворенного кремнезема в океаны: 4,2 ± 0,8 × 1014 г SiO2 год−1
- Подводный вулканизм и связанный с ним гидротермальный эманации: 1,9 ± 1,0 × 1014 г SiO2 год−1
- Ледниковый выветривание: 2 × 1012 г SiO2 год−1
- Низкотемпературное подводное выветривание океана базальты
- Некоторое количество кремнезема может также ускользнуть из поровых вод, обогащенных кремнеземом. пелагические отложения на морское дно
Когда организм погибает, часть кремнистого скелетного материала растворяется, поскольку он оседает через столб воды, обогащая глубокие воды растворенным кремнеземом.[3] Некоторые из кремнистых чешуек также могут сохраняться со временем в виде микрофоссилий в глубокое море отложения, открывая окно в современное и древнее планктон /протисты сообщества.[4] Этот биологический процесс действует, по крайней мере, с ранних Палеозой время, чтобы регулировать баланс кремнезема в океане: радиолярии (Кембрийский /Ордовик -Голоцен ), диатомеи (Меловой -Голоцен ) и силикофлагелляты (Меловой -Голоцен ) образуют основных участников глобального биогенного цикла кремнезема на протяжении всего геологическое время. На диатомовые водоросли приходится 43% первичной продукции океана, и они ответственны за большую часть добычи кремнезема из океанических вод в современном океане на протяжении большей части последних пятидесяти миллионов лет. Напротив, океаны Юрский период и старшего возраста, характеризовались радиолярии как основной тип, использующий диоксид кремния.[2] В настоящее время радиолярии являются вторыми (после диатомовых) основными продуцентами взвешенного аморфного кремнезема в водах океана. Их распределение колеблется от Арктический к Антарктика, наиболее многочисленны в экваториальной зоне. В экваториальном Тихий океан воды, например, можно наблюдать около 16000 особей на кубический метр.[4]
Цикл силикатного цикла привлекает все большее внимание ученых в последнее десятилетие по следующим причинам. Во-первых, современный морской Широко распространено мнение, что в цикле кремнезема преобладают диатомовые водоросли для фиксации и экспорта твердые частицы (включая органический углерод ), от эвфотическая зона в глубокий океан с помощью процесса, известного как биологический насос. В результате диатомовые водоросли и другие организмы, выделяющие кремнезем, играют решающую роль в глобальном развитии. цикл углерода и иметь возможность влиять на атмосферный CO2 концентрации в различных временных масштабах, секвестирование CO2 В океане. Эта связь между биогенный кремнезем и органический углерод, вместе со значительно более высоким потенциалом сохранения биогенных кремнистых соединений по сравнению с органическим углеродом, делает записи о накоплении опала очень интересными для палеоокеанография и палеоклиматология. Во-вторых, накопление биогенного кремнезема на морском дне содержит много информации о том, где находится океан. экспортное производство произошло во временных масштабах от сотен до миллионов лет. По этой причине записи отложений опалов предоставляют ценную информацию о крупномасштабных океанографических реорганизациях в геологическом прошлом, а также о палеопродуктивности. И наконец, средняя океаническая Время жительства для силиката - примерно 10 000–15 000 лет. Это относительно короткое время пребывания делает концентрации силикатов в океане и потоки чувствительными к ледниковый /межледниковый возмущения, а значит отличный доверенное лицо для оценки изменений климата.[3][5]
Остатки диатомовых водорослей и других организмов, потребляющих диоксид кремния, обнаруживаются в виде опаловых отложений в пелагических глубоководных отложениях. Пелагические отложения, содержащие значительное количество кремнеземистых биогенных остатков, обычно называют кремнистый ил. Кремнистый ил особенно распространен в современном океане в высоких широтах северного и южного полушарий. Отличительной особенностью распределения кремнистого ила является ок. Полоса шириной 200 км, протянувшаяся через Южный океан. Некоторые экваториальные районы апвеллинг, куда питательные вещества в изобилии и продуктивность высока, также характеризуются местными кремнистыми илами. Кремнистые илы состоят в основном из остатков диатомовых водорослей и радиолярий, но могут также включать другие кремнистые организмы, такие как силикофлагелляты и губка спикулы. Ил диатомей встречается в основном в высокоширотных районах и вдоль некоторых окраин континентов, тогда как илы радиолярий более характерны для экваториальных районов. Кремнистые илы видоизменяются и при захоронении превращаются в слоистые. кремы.[2]
Диатомовые водоросли как в пресной, так и в соленой воде извлекают кремнезем из воды, чтобы использовать его в качестве компонента своих клеточных стенок. Точно так же некоторые голопланктонный простейшие (Радиолярии ), немного губки, а некоторые растения (лист фитолиты ) используют кремний в качестве конструкционного материала. Кремний, как известно, необходим цыплятам и крысам для роста и развития скелета. Кремний находится в человеке соединительной ткани, кости, зубы, кожа, глаза, железы и органы.
BSi - это кремнезем, получаемый при производстве растворенный кремнезем. BSi можно накапливать «напрямую» в морские отложения (через экспорт) или обратно в растворенный диоксид кремния в водной толще.
Все чаще изотопные отношения кислорода (O18: O16) и кремний (Si30: Si28) анализируются из BSi, сохранившейся в озерных и морских отложениях, чтобы получить записи прошлого изменение климата и круговорот питательных веществ (Де Ла Роча, 2006; Ленг и Баркер, 2006). Это особенно ценный подход, учитывая роль диатомеи в глобальном круговороте углерода. Кроме того, изотопный анализ BSi полезен для отслеживания прошлых изменений климата в таких регионах, как Южный океан, где мало биогенных карбонаты сохранены.
Баланс морского биогенного кремнезема
Реки и подводная лодка гидротермальный приток эманаций 6,1 × 1014 г SiO2 год−1 к морской среде. Примерно две трети этого количества кремнезема хранится в континентальная окраина и глубокое море депозиты. Кремнистые глубоководные отложения, расположенные под Антарктическая конвергенция (зона конвергенции ) содержат около 25% кремнезема, поступающего в океаны (т.е. 1,6 × 1014 г SiO2 год−1) и, следовательно, образуют один из основных поглотителей кремнезема на Земле. Наибольшие скорости накопления биогенного кремнезема в этом районе наблюдаются в Южной Атлантике, достигая 53 см. Кир.−1 в течение последних 18000 г. Кроме того, обширное биогенное накопление кремнезема было зарегистрировано в глубоководных отложениях Берингово море, Охотское море, и Субарктический Северная часть Тихого океана. Суммарные скорости накопления биогенного кремнезема в этих регионах составляют около 0,6 × 1014 г SiO2 год−1, что эквивалентно 10% растворенного кремнезема, поступающего в океаны. Континентальные окраинные области апвеллинга, такие как Калифорнийский залив, побережье Перу и Чили, характеризуются одними из самых высоких в мире темпов накопления биогенного кремнезема. Например, скорость накопления биогенного кремнезема 69 г SiO22/см2/ kyr зарегистрированы для Калифорнийского залива. Из-за ограниченного по горизонтали характера этих зон быстрого биогенного накопления кремнезема, на районы апвеллинга приходится только около 5% растворенного кремнезема, поступающего в океаны. Наконец, чрезвычайно низкие темпы накопления биогенного кремнезема наблюдались в обширных глубоководных отложениях Атлантического, Индийского и Тихого океанов, что делает эти океаны незначительными для глобального морского баланса кремнезема.[6]
Основные опускания кремнезема в современных океанах
Крупномасштабная океаническая циркуляция оказывает прямое влияние на опал осаждение. Тихий океан (характеризующийся бедными питательными веществами поверхностными водами и глубокими водами, богатыми питательными веществами) и Атлантический океан с циркуляцией благоприятствуют производство /сохранение кремнезема и карбоната соответственно. Например, отношения Si / N и Si / P увеличиваются от Атлантики до Тихого и Южного океана, в пользу опала по сравнению с карбонат производители. Следовательно, современная конфигурация крупномасштабной океанической циркуляции привела к локализации основных зон захоронения опалов в экваториальной части Тихого океана, в системах апвеллинга течения восточной границы и, безусловно, в наиболее важном - Южном океане.[5]
В водах современного Тихого океана и Южного океана обычно наблюдается увеличение отношения Si / N на средней глубине, что приводит к увеличению экспорта опала (~ увеличению производства опала). В Южном океане и северной части Тихого океана эта взаимосвязь между экспортом опала и отношением Si / N меняется с линейной на экспоненциальную для отношений Si / N более 2. Это постепенное увеличение важности силиката (Si) по сравнению с азотом (N) имеет огромные последствия для биологического производства океана. Изменение соотношения питательных веществ способствует выбору диатомеи в качестве основных продуцентов по сравнению с другими (например, кальцифицирующими) организмами. Например, микрокосм эксперименты показали, что диатомовые водоросли являются суперконкурентами DSi и доминируют над другими продуцентами DSi выше 2 мкМ. Следовательно, предпочтение будет отдаваться экспорту опала по сравнению с экспортом карбоната, что приведет к увеличению производства опала. Южный океан и северная часть Тихого океана также демонстрируют максимум биогенных силикатов / Cорганический отношения потоков и, таким образом, состоят в обогащении биогенным силикатом по сравнению с Cорганический экспортный поток. Это совокупное увеличение сохранности и экспорта опалов делает Южный океан самым важным. раковина для DSi сегодня.[5]
В современных водах Тихого и Южного океанов промежуточные и глубоководные воды характеризуются более высоким содержанием DSi по сравнению с Атлантическим океаном. Эта межбассейновая разница в DSi увеличивает потенциал сохранения опалов в Тихом и Южном океанах по сравнению с их атлантическими аналогами. Воды Атлантического океана, истощенные по DSi, производят относительно меньше окремненный организмов, что оказывает сильное влияние на сохранение их панцири. Этот механизм лучше всего проиллюстрирован при сравнении систем апвеллинга в Перу и северо-западной Африке. В растворение Коэффициент продуктивности в Атлантическом апвеллинге намного выше, чем в Тихоокеанском апвеллинге. Это связано с тем, что прибрежные воды источника апвеллинга намного богаче DSi у побережья Перу, чем у северо-западной Африки.[5]
Цикличность и накопление биогенного кремнезема в отложениях Южного океана
Отложения Южного океана являются основным поглотителем биогенного кремнезема (50-75% всей океанической массы 4,5 × 1014 г SiO2 год−1; DeMaster, 1981), но лишь незначительная раковина для органических углерод (<1% океанических 2 × 1014 г органического углерода в год−1). Эти относительно высокие скорости накопления биогенного кремнезема в отложениях Южного океана (преимущественно под полярным фронтом) по сравнению с органическим углеродом (60: 1 по весу) являются результатом преимущественного сохранения биогенного кремнезема в толще воды Антарктики. В отличие от того, что считалось ранее, эти высокие скорости накопления биогенного кремнезема не являются результатом высоких темпов накопления основное производство. Биологическое производство в Южном океане сильно ограничено из-за низкого уровня сияние в сочетании с глубокими смешанными слоями и / или ограниченным количеством микроэлементы, Такие как утюг.[7] Это преимущественное сохранение биогенного кремнезема по сравнению с органическим углеродом проявляется в постоянно увеличивающемся соотношении кремнезем / органический углерод в зависимости от глубины водной толщи. Около тридцати пяти процентов биогенного кремнезема, производимого в эвфотический зона выдерживает растворение в поверхностном слое; тогда как только 4% органического углерода уходит микробный деградация в этих приповерхностных водах. Следовательно, во время осаждения через толщу воды происходит значительное разделение органического углерода и кремнезема. Накопление биогенного кремнезема на морском дне составляет 12% от поверхностной продукции, тогда как скорость накопления органического углерода на морском дне составляет только <0,5% от поверхностной продукции. В результате на полярные отложения приходится большая часть накопления биогенного кремнезема в океане, но лишь небольшая часть потока осадочного органического углерода.[7]
BSi производство

Средняя суточная скорость BSi сильно зависит от региона:
- Прибрежный апвеллинг: 46 ммоль.м−2.d−1
- Суб-Арктический Тихий океан: 18 ммоль.м−2.d−1
- Южный океан: 3–38 ммоль.м−2.d−1
- посреди океана круговороты: 0,2–1,6 ммоль.м−2.d−1
Точно так же интегрированный годовой объем производства BSi сильно зависит от региона:
- Прибрежный апвеллинг: 3 × 1012 мол. год−1
- Субарктический Тихий океан: 8 × 1012 мол. год−1
- Южный океан: 17–37 × 1012 мол. год−1
- срединно-океанические круговороты: 26 × 1012 мол. год−1
Производство BSi контролируется:
- Растворенный кремнезем доступность, однако, постоянная половина насыщения Kμ для роста, ограниченного кремнием, ниже, чем Ks для поглощения кремния.
- Доступность света: нет необходимости в прямом освещении; поглощение кремния на 2-кратной глубине фотосинтез; Поглощение кремния продолжается ночью, но клетки должны активно расти.
- Микроэлементы доступность.
Роспуск BSi
Растворение BSi контролируется:
- Термодинамика из растворимость: Температура (от 0 до 25 ° C - увеличение в 50 раз).
- Скорость опускания: Структура пищевой сети - пастбища, фекальные гранулы, выброшенные кормовые структуры, агрегирование - быстрая транспортировка.
- Бактериальный деградация из органическая матрица (Бидл и Азам, 1999).
Сохранение BSi
Сохранность BSi измеряется:
- Скорость осаждения, в основном ловушки для наносов (Хондзё);
- Бентосный реминерализация нормы («переработка»), бентосные флюсовая камера (Берельсон);
- Концентрация BSi в отложениях, химическое выщелачивание в щелочной решение, зависит от сайта, необходимо различать литогенный по сравнению с биогенным Si, дифракция рентгеновских лучей.
Сохранение BSi контролируется:
- Скорость оседания;
- Грунтовая вода концентрация растворенного кремнезема: насыщение при 1,100 мкмоль / л;
- Покрытия поверхности: растворенный алюминий изменяет растворимость осажденных частиц биогенного кремнезема, растворенный кремнезем также может осадок с Алом как глина или покрытия Al-Si.
Опаловый кремнезем на Марсе
в Кратер Гусева Марса, Марсоход для исследования Марса Дух случайно обнаружил опаловый кремнезем. Одно из его колес ранее было обездвижено и, таким образом, эффективно закапывало марсианин. реголит когда он тащился за марсоходом. Более поздний анализ показал, что кремнезем свидетельствует о гидротермальных условиях.[8]
Рекомендации
- ^ Корадин, Т., Лопес, П.Дж. (2003). "Биогенное формирование рисунка из кремнезема: простая химия или тонкая биология?" ChemBioChem 3: 1-9.
- ^ а б c Боггс, С. (2005). «Принципы седиментологии и стратиграфии (4-е издание)». Pearson Education, Inc, 662p.
- ^ а б c d ДеМастер, Д.Дж. (1981). «Поставка и накопление кремнезема в морской среде». Geochimica et Cosmochimica Acta 45: 1715-1732.
- ^ а б c Эрлих и др. (2010). «Современные взгляды на обессиливание: растворение биокремнезема и абиотического кремнезема в естественных и искусственных средах». Chem. Ред. 110: 4656-4689.
- ^ а б c d Кортезе, Г., Жерсонде, Р. (2004). «Смещение отложений опала в Мировом океане за последние 15 млн лет». Письма о Земле и планетологии 224: 509-527.
- ^ ДеМастер, Д.Дж. (2002). «Накопление и круговорот биогенного кремнезема в Южном океане: пересмотр морского бюджета кремнезема». Deep-Sea Research Part II 49: 3155-3167
- ^ а б ДеМастер, Д. (1992). "Круговорот и накопление биогенного кремнезема и органических веществ в высокоширотных средах: море Росса". Океанография 5 (3): 147-153
- ^ [1] Ruff, S. W., et al. (2011). «Характеристики, распространение, происхождение и значение опалового кремнезема, обнаруженного марсоходом Spirit в кратере Гусева на Марсе». J. Geophys. Рез., 116, E00F23.
- Бжезинский, М.А. (1985). «Отношение Si: C: N морских диатомовых водорослей: межвидовая изменчивость и влияние некоторых переменных окружающей среды». Журнал психологии 21(3): 347-357.
- Де Ла Роша, C.L. (2006). «Опаловые прокси палеоэкологических условий». Глобальные биогеохимические циклы 20. Дои:10.1029 / 2005GB002664.
- Дагдейл, Р. К. и Ф. П. Вилкерсон (1998). «Силикатное регулирование новой добычи в экваториальном тихоокеанском апвеллинге». Природа 391(6664): 270.
- Дагдейл, Р. К., Ф. П. Вилкерсон и др. (1995). «Роль силикатного насоса в создании нового производства». Глубоководные исследования I 42 (5): 697-719.
- Ленг, М.Дж., Баркер, П.А. (2006). «Обзор изотопного состава кислорода озерной диатомовой кремнезема для реконструкции палеоклимата». Обзоры наук о Земле 75:5-27.
- Рагено, О., П. Трегер и др. (2000). «Обзор цикла Si в современном океане: недавний прогресс и недостающие пробелы в применении биогенного опала в качестве показателя палеопродуктивности». Глобальные и планетарные изменения 26: 317-365.
- Такеда, С. (1998). «Влияние доступности железа на соотношение потребления питательных веществ диатомовыми водорослями в океанических водах». Природа 393: 774-777.
- Вернер, Д. (1977). Биология диатомовых водорослей. Беркли и Лос-Анджелес, Калифорнийский университет Press.