Основное производство - Primary production
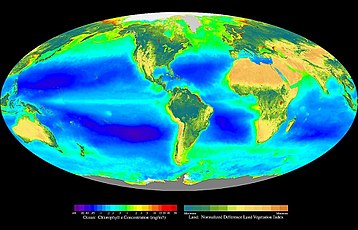
В экология, основное производство это синтез органические соединения из атмосферного или водного углекислый газ. В основном это происходит в процессе фотосинтез, который использует свет в качестве источника энергии, но также происходит через хемосинтез, который использует окисление или восстановление неорганических химических соединений в качестве источника энергии. Почти все жизнь на Земле прямо или косвенно зависит от первичного производства. Организмы, ответственные за первичное производство, известны как первичные производители или же автотрофы, и формируют основу пищевая цепочка. В наземные экорегионы, это в основном растения, пока в водные экорегионы водоросли преобладают в этой роли. Экологи различают первичное производство как сеть или же валовой, первый учитывает потери для таких процессов, как клеточное дыхание, последнее нет.
Обзор
Первичное производство - это производство химическая энергия в органических соединениях живыми организмы. Основным источником этой энергии является Солнечный свет но мельчайшая доля первичного производства обусловлена литотрофный организмов, использующих химическую энергию неорганический молекулы.
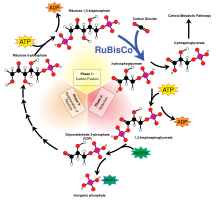
Независимо от источника, эта энергия используется для синтеза сложных Органические молекулы из более простых неорганических соединений, таких как углекислый газ (CO2) и воды (ЧАС2О). Следующие два уравнения являются упрощенными представлениями фотосинтеза (вверху) и (одна форма) хемосинтез (Нижний):
- CO2 + H2O + свет → CH2О + О2
- CO2 + O2 + 4 часа2S → CH2O + 4 S + 3 H2О
В обоих случаях конечной точкой является полимер из уменьшенный углевод, (CH2O)п, как правило, молекулы, такие как глюкоза или другой сахара. Эти относительно простые молекулы затем могут быть использованы для дальнейшего синтеза более сложных молекул, включая белки, сложные углеводы, липиды, и нуклеиновые кислоты, или быть вздохнул выполнять работай. Потребление первичных производителей гетеротрофный организмы, такие как животные, затем переносит эти органические молекулы (и запасенную в них энергию) вверх по пищевой сети, заправляя все земной шар живые системы.
Валовая первичная продукция и чистая первичная продукция
Валовая первичная продукция (GPP) - это количество химической энергии, обычно выражаемое в виде углерода. биомасса, которые первичные производители создают за определенный период времени. Некоторая часть этой фиксированной энергии используется первичными производителями для клеточное дыхание и поддержание существующих тканей (например, «дыхание роста» и «поддерживающее дыхание ").[1][2] Оставшаяся фиксированная энергия (т.е. масса фотосинтата) обозначается как чистая первичная продукция (АЭС).
- NPP = GPP - дыхание [растениями]
Чистая первичная продукция - это скорость, с которой все автотрофы в экосистема производят чистую полезную химическую энергию. Как уже отмечалось, он равен разнице между скоростью, с которой растения в экосистеме производят полезную химическую энергию (GPP), и скоростью, с которой они используют часть этой энергии во время дыхания. Чистая первичная продукция доступна для направления на рост и воспроизводство первичных производителей. Таким образом, он доступен для употребления травоядными животными.
Как валовая, так и чистая первичная продукция обычно выражается в единицах массы на единицу площади за единицу времени. В наземных экосистемах масса углерода на единицу площади в год (г С м−2 год−1) чаще всего используется в качестве единицы измерения. Обратите внимание, что иногда проводится различие между «производством» и «производительностью», причем первое - это количество произведенного материала (г C m−2), последнее - скорость его производства (г C м−2 год−1), но эти термины чаще используются как синонимы.
Наземное производство
На суше почти все первичное производство сейчас осуществляется сосудистые растения, с небольшой долей из водорослей и несосудистые растения Такие как мхи и печеночники. Перед эволюция Из сосудистых растений несосудистые растения, вероятно, играли более значительную роль. Первичное производство на суше - это функция многих факторов, но в основном локальных гидрология и температура (последний до некоторой степени зависит от света, в частности, фотосинтетически активного излучения (ФАР), источника энергии для фотосинтеза). Хотя растения покрывают большую часть поверхности Земли, они сильно сокращаются там, где температура слишком высока или когда необходимые растительные ресурсы (в основном вода и PAR) ограничены, например, пустыни или же полярные регионы.
Вода «потребляется» растениями в процессе фотосинтеза (см. Выше) и испарение. Последний процесс (на который приходится около 90% водопотребления) обусловлен испарение воды из листья растений. Транспирация позволяет растениям переносить воду и минеральная питательные вещества от почва в районы произрастания, а также охлаждает растение. Распространение водяного пара из листа, сила, которая вызывает транспирацию, регулируется структурами, известными как устьица. Эти структуры также регулируют диффузию углекислого газа из атмосферы в лист, так что уменьшение потери воды (частичное закрытие устьиц) также снижает приток углекислого газа. Некоторые растения используют альтернативные формы фотосинтеза, называемые Метаболизм крассуловой кислоты (CAM) и C4. Они используют физиологический и анатомический адаптации для повышения эффективности водопользования и обеспечения возможности увеличения первичной продукции в условиях, которые обычно ограничивают фиксацию углерода за счет C3 растения (большинство видов растений).
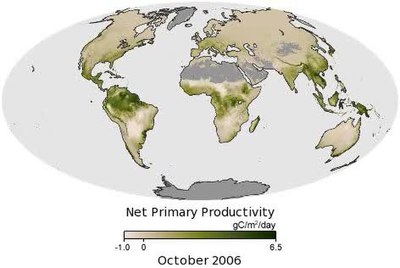
Как показано на анимации, бореальные леса Канады и России демонстрируют высокую продуктивность в июне и июле, а затем медленное сокращение осенью и зимой. Круглый год тропические леса в Южной Америке, Африке, Юго-Восточной Азии и Индонезии отличаются высокой продуктивностью, что неудивительно с учетом обилия солнечного света, тепла и осадков. Однако даже в тропиках урожайность колеблется в течение года. Например, бассейн Амазонки демонстрирует особенно высокую продуктивность примерно с августа по октябрь - период засушливого сезона в этом районе. Поскольку деревья имеют доступ к обильному запасу грунтовых вод, который накапливается в сезон дождей, они растут лучше, когда дождливое небо очищается и позволяет большему количеству солнечного света достигать леса.[3]
Океаническое производство

В обратном порядке на суше, в океанах, почти весь фотосинтез осуществляется водорослями, при этом небольшую долю вносят сосудистые растения и другие группы. Водоросли включают в себя широкий спектр организмов, от отдельных плавающих клеток до прикрепленных водоросли. В их число входят фотоавтотрофы из самых разных групп. Эубактерии являются важными фотосинтезаторами как в океанических, так и в наземных экосистемах, а некоторые археи находятся фототрофный, ни один из них не использует фотосинтез с выделением кислорода.[4] Номер эукариоты вносят значительный вклад в первичную продукцию в океане, в том числе зеленые водоросли, бурые водоросли и красные водоросли, и разнообразная группа одноклеточных групп. Сосудистые растения также представлены в океане такими группами, как морские травы.
В отличие от наземных экосистем, большая часть первичной продукции в океане осуществляется свободноживущими. микроскопические организмы называется фитопланктон. Более крупные автотрофы, такие как морские травы и макроводоросли (водоросли ) обычно ограничиваются прибрежный зона и прилегающие мелководья, где они могут прикреплять на нижележащий субстрат, но все еще в фотическая зона. Есть исключения, например Саргассум, но подавляющее большинство свободно плавающих видов продукции происходит внутри микроскопических организмов.

Факторы, ограничивающие первичную продукцию в океане, также сильно отличаются от факторов на суше. Наличие воды, очевидно, не является проблемой (хотя ее соленость возможно). Точно так же и температура, влияя на метаболический ставки (см. Q10 ), в океане распространяется менее широко, чем на суше, поскольку теплоемкость изменения температуры буферов морской воды и образование морской лед изолирует это при более низких температурах. Однако наличие света, источника энергии для фотосинтеза и минеральных веществ. питательные вещества, строительные блоки для нового роста, играют решающую роль в регулировании первичной продукции в океане.[5] Доступные модели земной системы предполагают, что текущие биогеохимические изменения океана могут вызвать сокращение океанских АЭС от 3% до 10% от текущих значений в зависимости от сценария выбросов.[6]
Свет

Солнечная зона океана называется фотическая зона (или эвфотическая зона). Это относительно тонкий слой (10–100 м) у поверхности океана, где достаточно света для фотосинтеза. Для практических целей толщина световой зоны обычно определяется глубиной, на которой свет достигает 1% от его поверхностного значения. Свет ослабленный вниз по толщине воды своим поглощение или же рассеяние самой водой, а также растворенным или твердым веществом в ней (включая фитопланктон).
Чистый фотосинтез в водной толще определяется взаимодействием между фотической зоной и смешанный слой. Турбулентное перемешивание к ветер энергия на поверхности океана гомогенизирует водную толщу по вертикали до тех пор, пока турбулентность рассеивается (создавая вышеупомянутый смешанный слой). Чем глубже смешанный слой, тем ниже среднее количество света, перехватываемого внутри него фитопланктоном. Смешанный слой может варьироваться от более мелкого, чем фотическая зона, до гораздо более глубокого, чем фотическая зона. Когда он намного глубже световой зоны, это приводит к тому, что фитопланктон проводит слишком много времени в темноте, чтобы мог произойти чистый рост. Максимальная глубина смешанного слоя, в которой может происходить чистый рост, называется критическая глубина. Пока имеется достаточное количество питательных веществ, чистая первичная продукция происходит всякий раз, когда смешанный слой мельче критической глубины.
Как сила ветра, так и доступность света на поверхности океана зависят от различных пространственных и временных масштабов. Наиболее характерным из них является сезонный цикл (вызвано последствия Земли осевой наклон ), хотя магнитуды ветра также имеют сильные пространственные компоненты. Следовательно, первичное производство в умеренный такие регионы, как Североатлантический является очень сезонным и меняется как в зависимости от падающего света на поверхность воды (снижается зимой), так и от степени перемешивания (увеличивается зимой). В тропический регионы, такие как круговороты в середине главного бассейны, свет может незначительно меняться в течение года, и смешивание может происходить только эпизодически, например, во время больших штормы или же ураганы.
Питательные вещества

Смешивание также играет важную роль в ограничении первичной продукции питательными веществами. Неорганические питательные вещества, такие как нитрат, фосфат и кремниевая кислота необходимы фитопланктону для синтезировать их клетки и клеточные механизмы. Потому что гравитационный опускание твердых частиц (например, планктон, мертвый или фекальный материал), питательные вещества постоянно теряются из фотической зоны и пополняются только путем смешивания или апвеллинг глубокой воды. Это усугубляется тем, что в летнее время солнечное нагревание и уменьшение ветра увеличивает вертикальную стратификацию и приводит к сильной термоклин, поскольку это затрудняет унос более глубокой воды при перемешивании ветром. Следовательно, между событиями перемешивания первичная продукция (и возникающие в результате процессы, которые приводят к оседанию твердых частиц) постоянно действует, чтобы потреблять питательные вещества в смешанном слое, и во многих регионах это приводит к истощению питательных веществ и снижению продуктивности смешанного слоя летом (даже в наличие обильного света). Однако до тех пор, пока световая зона достаточно глубока, первичная продукция может продолжаться ниже смешанного слоя, где ограниченная светом скорость роста означает, что питательных веществ часто больше.
Утюг
Еще один относительно недавно обнаруженный фактор, играющий значительную роль в первичной продукции океана, - это микронутриент утюг.[7] Это используется как кофактор в ферменты участвует в таких процессах, как снижение нитратов и азотфиксация. Основным источником железа в океанах является пыль с Земли. пустыни, поднятый и доставленный ветром, как эоловая пыль.
В районах океана, удаленных от пустынь или недоступных для пылевых ветров (например, Южный и Северная часть Тихого океана океанов), нехватка железа может серьезно ограничить объем первичной продукции, которая может произойти. Эти области иногда называют HNLC (С высоким содержанием питательных веществ и низким содержанием хлорофилла), поскольку дефицит железа ограничивает рост фитопланктона и оставляет избыток других питательных веществ. Некоторые ученые предложили введение железа в эти области как средство увеличения первичной продуктивности и улавливания двуокиси углерода из атмосферы.[8]
Измерение
Методы измерения первичной продукции различаются в зависимости от того, является ли валовая или чистая продукция желаемой мерой, а также от того, в центре внимания находятся наземные или водные системы. Валовое производство почти всегда труднее измерить, чем чистое, из-за дыхания, которое представляет собой непрерывный и непрерывный процесс, в ходе которого потребляются некоторые продукты первичного производства (например, сахара), прежде чем их можно будет точно измерить. Кроме того, наземные экосистемы, как правило, более сложны, поскольку значительная часть общей продуктивности перемещается в подземные органы и ткани, где ее сложно измерить с точки зрения логистики. Мелководные водные системы также могут столкнуться с этой проблемой.
Масштаб также сильно влияет на методы измерения. Скорость ассимиляции углерода тканями, органами, целыми растениями или образцами планктона растений может быть определена количественно биохимические методы, но эти методы явно не подходят для крупномасштабных наземных полевых условий. Здесь чистая первичная продукция почти всегда является желаемой переменной, а методы оценки включают различные методы оценки изменений сухой массы биомассы с течением времени. Оценки биомассы часто преобразуются в показатель энергии, такой как килокалории, с помощью эмпирически определен коэффициент преобразования.
Наземный

В наземных экосистемах исследователи обычно измеряют чистую первичную продукцию (NPP). Несмотря на простое определение, полевые измерения, используемые для оценки продуктивности, варьируются в зависимости от исследователя и биома. В полевых оценках редко учитываются подземная продуктивность, травоядность, оборот, подстилка, летучие органические соединения, корневые экссудаты и выделение симбиотический микроорганизмы. Оценка АЭС на основе биомассы приводит к заниженной оценке АЭС из-за неполного учета этих компонентов.[9][10] Однако многие полевые измерения хорошо коррелируют с АЭС. Существует ряд исчерпывающих обзоров полевых методов, используемых для оценки АЭС.[9][10][11] Оценки экосистемное дыхание, общий диоксид углерода, производимый экосистемой, также может быть получен с измерения потока газа.
Основной неучтенный пул - подземная продуктивность, особенно производство и оборот корней. Подземные компоненты АЭС трудно измерить. BNPP (подземная АЭС) часто оценивается на основе соотношения ANPP: BNPP (наземная АЭС: подземная АЭС), а не прямых измерений.
Валовая первичная продукция может быть оценена на основе измерений чистого экосистемного обмена (NEE) диоксида углерода, производимого метод вихревой ковариации. В ночное время этот метод измеряет все компоненты дыхания экосистемы. Это дыхание масштабируется до дневных значений и далее вычитается из NEE.[12]
Луга

Чаще всего предполагается, что для измерения ЧПП используется пиковая биомасса на корню. В системах с устойчивой стоячей подстилкой обычно регистрируется живая биомасса. Измерения пиковой биомассы более надежны, если система преимущественно однолетняя. Однако многолетние измерения могли бы быть надежными, если бы была синхронная фенология, обусловленная сильным сезонным климатом. Эти методы могут недооценивать ANPP на пастбищах на целых 2 (умеренный ) до 4 (тропический ) сложить.[10] Повторные измерения живой и мертвой биомассы на корню дают более точные оценки всех пастбищ, особенно тех, которые характеризуются высокой текучестью, быстрым разложением и межвидовой изменчивостью во времени пика биомассы. Водно-болотные угодья продуктивность (болота и топи) измеряется аналогичным образом. В Европа, ежегодное кошение делает очевидным годовой прирост биомассы водно-болотных угодий.
Леса
Методы, используемые для измерения продуктивности лесов, более разнообразны, чем методы измерения продуктивности пастбищ. Прирост биомассы в зависимости от насаждения аллометрия плюс опадная подстилка считается подходящим, хотя и неполным, учетом чистой первичной продукции надземных водоемов (ANPP).[9] Полевые измерения, используемые в качестве прокси для ANPP, включают годовой опад подстилки, диаметр или прирост базальной площади (DBH или BAI) и приращение объема.
Водный
| Часть серии о |
| Цикл углерода |
|---|
 |
В водных системах первичная продукция обычно измеряется с использованием одного из шести основных методов:[13]
- вариации концентрации кислорода в запечатанной бутылке (разработано Гаардером и Граном в 1927 году)
- включение неорганических углерод-14 (14C в виде бикарбонат натрия ) в органическое вещество[14][15]
- Стабильные изотопы кислорода (16О, 18O и 17O)[16][17]
- кинетика флуоресценции (техника все еще является темой исследования)
- Стабильные изотопы углерода (12C и 13C)[18]
- Соотношение кислород / аргон [19]
В методе, разработанном Гаардером и Граном, используются вариации концентрации кислорода в различных экспериментальных условиях для определения валовой первичной продукции. Обычно три одинаковых прозрачных сосуда заполняются пробой воды и закупоренный. Первый анализируется немедленно и используется для определения начальной концентрации кислорода; обычно это делается путем выполнения Винклер-титрование. Два других сосуда инкубируют, по одному на свету и затемненных. По прошествии определенного времени эксперимент заканчивается, и измеряется концентрация кислорода в обоих сосудах. Поскольку фотосинтез не происходил в темном сосуде, он обеспечивает определенную экосистемное дыхание. Световой сосуд разрешает как фотосинтез, так и дыхание, поэтому обеспечивает меру чистого фотосинтеза (то есть производство кислорода посредством фотосинтеза вычитает потребление кислорода посредством дыхания). Затем получают валовую первичную продукцию, добавляя потребление кислорода в темном сосуде к чистому производству кислорода в светлом сосуде.
Техника использования 14Включение C (добавлено как обозначено Na2CO3), чтобы сделать вывод о первичной продукции, сегодня чаще всего используется, потому что он чувствителен и может использоваться во всех океанских средах. В качестве 14C - это радиоактивный (через бета-распад ), относительно просто измерить его включение в органический материал с помощью таких устройств, как сцинтилляционные счетчики.
В зависимости от выбранного времени инкубации можно оценить чистую или валовую первичную продукцию. Валовую первичную продукцию лучше всего оценить, используя относительно короткое время инкубации (1 час или меньше), поскольку потеря инкорпорированного 14C (путем дыхания и выделения / экссудации органических материалов) будет более ограниченным. Чистая первичная продукция - это доля валовой продукции, остающаяся после того, как в результате этих процессов потерь была израсходована часть фиксированного углерода.
Процессы убытков могут составлять от 10 до 60% зарегистрированных 14C в зависимости от периода инкубации, условий окружающей среды (особенно температуры) и экспериментальных разновидность использовал. Помимо тех, которые вызваны физиологией самого объекта эксперимента, необходимо также учитывать потенциальные потери из-за активности потребителей. Это особенно верно в экспериментах с использованием естественных скоплений микроскопических автотрофов, где невозможно изолировать их от потребителей.
Методы на основе стабильных изотопов и O2Отношения / Ar имеют то преимущество, что позволяют оценить частоту дыхания на свету без необходимости инкубации в темноте. Среди них метод тройных изотопов кислорода и O2/ Ar обладают дополнительным преимуществом, так как не требуют инкубации в закрытых контейнерах и O2/ Ar можно даже непрерывно измерять в море с помощью масс-спектрометрии на входе уравновешивающего устройства (EIMS)[20] или масс-спектрометрия на входе в мембрану (MIMS).[21] Однако, если желательны результаты, относящиеся к углеродному циклу, вероятно, лучше полагаться на методы, основанные на изотопах углерода (а не кислорода). Важно отметить, что метод на основе стабильных изотопов углерода - это не просто адаптация классического 14C, но совершенно другой подход, который не страдает проблемой отсутствия учета рециркуляции углерода во время фотосинтеза.
Глобальный
Как первичное производство в биосфера является важной частью цикл углерода, оценка его в глобальном масштабе важна в Наука о земных системах. Однако количественное определение первичного производства в этом масштабе затруднено из-за диапазона среда обитания на Земле, и из-за воздействия Погода события (наличие солнечного света, воды) от его изменчивости. С помощью спутниковое -производные оценки Нормализованный разностный индекс растительности (NDVI) для наземных местообитаний и морской поверхности хлорофилл для океанов, по оценкам, общая (фотоавтотрофная) первичная продукция Земли составила 104,9 петаграммы углерода в год (Пг C год−1; эквивалентно не-SI Gt C год−1).[22] Из них 56,4 пг C в год−1 (53,8%) были продуктом наземных организмов, а остальные 48,5 пг C в год−1, приходилось на океаническую продукцию.
Масштабирование оценок GPP на уровне экосистемы на основе ковариация вихря измерения чистого обмена экосистемами (см. выше) на региональные и глобальные значения с использованием пространственных деталей различных переменных-предикторов, таких как климатические переменные и данные дистанционного зондирования fAPAR или же LAI привело к валовому первичному производству на суше 123 ± 8 Гт углерода (НЕ двуокиси углерода) в год в течение 1998-2005 гг. [23]
В ареал По оценкам, производство земли составляло примерно 426 г C м−2 год−1 (без учета участков с постоянным ледяным покровом), а для океанов - 140 г С м−2 год−1.[22] Еще одно существенное различие между сушей и океанами заключается в их постоянных запасах - хотя на долю океанических автотрофов приходится почти половина всей продукции, на долю океанических автотрофов приходится только около 0,2% общей биомассы.
Оценки
Первичную продуктивность можно оценить с помощью множества прокси. Один из них, имеющий особое отношение к геологической летописи, - это барий, концентрация которого в морских отложениях возрастает в соответствии с первичной продуктивностью на поверхности.[24][25][26]
Человеческое воздействие и присвоение
Человеческие общества являются частью цикла АЭС Земли, но оказывают на него непропорционально большое влияние.[27] В 1996 году Хосеп Гари разработал новый индикатор устойчивого развития, основанный именно на оценке человеческого присвоения АЭС: он придумал его «HANPP» (человеческое присвоение чистого первичного производства) и представил его на вступительной конференции Европейского общества защиты окружающей среды. Экологическая экономика.[28] С тех пор HANPP получила дальнейшее развитие и широко применяется в исследованиях по экологической экономике, а также в анализе политики в области устойчивого развития. HANPP представляет собой пример воздействия человека на природу и может применяться в различных географических масштабах, а также в глобальном масштабе.
Широкая степень использования ресурсов планеты людьми, в основном за счет землепользование, приводит к различным уровням воздействия на фактическая АЭС (АЭСдействовать). Хотя в некоторых регионах, например Нил Долина, орошение привел к значительному увеличению первичного производства, на большей части планеты наблюдается заметная тенденция Снижение АЭС из-за изменения земель (ΔNPPLC) 9,6% по всей суше.[29] В дополнение к этому, конечное потребление людьми увеличивает общую HANPP. [27] до 23,8% потенциальная растительность (АЭС0).[29] По оценкам, в 2000 г. 34% свободной ото льда площади суши Земли (12% пахотная земля; 22% пастбище ) был посвящен человеческому земледелию.[30] Это непропорционально большое количество снижает доступную для других видов энергию, оказывая заметное влияние на биоразнообразие, потоки углерода, воды и энергии, и экосистемные услуги,[29] и ученые задаются вопросом, насколько большой может быть эта доля до того, как эти службы начнут отказываться.[31] Ожидается также сокращение ЧЭС в океане в результате продолжающегося изменения климата, потенциально влияющего на морские экосистемы (~ 10% глобального биоразнообразия) и товары и услуги (1-5% от глобального общего количества), которые предоставляют океаны.[6]
Смотрите также
Рекомендации
- ^ Амтор, Дж. и Baldocchi, D.D. (2001). Дыхание и чистая первичная продукция наземных высших растений. В Земная глобальная производительность, Academic Press, 33-59
- ^ Sigman, D.M .; Хайн, М. (2012). «Биологическая продуктивность океана» (PDF). Знание природы образования. 3 (6): 1–16. Получено 2015-06-01.
«Валовая первичная продукция» (GPP) относится к общей скорости производства органического углерода автотрофами, а «дыхание» относится к энергоемкому окислению органического углерода обратно в диоксид углерода. «Чистая первичная продукция» (NPP) - это GPP за вычетом собственной скорости дыхания автотрофов; таким образом, это скорость, с которой полный метаболизм фитопланктона производит биомассу. «Вторичная продукция» (SP) обычно относится к скорости роста гетеротрофной биомассы.
- ^ а б «Чистая первичная производительность: глобальные карты». earthobservatory.nasa.gov. 26 марта 2018 г.. Получено 26 марта 2018.
- ^ Шефер Г., Энгельхард М., Мюллер В. (1 сентября 1999 г.). «Биоэнергетика архей». Microbiol. Мол. Биол. Rev. 63 (3): 570–620. Дои:10.1128 / MMBR.63.3.570-620.1999. ЧВК 103747. PMID 10477309.
- ^ Sigman, D.M .; Хайн, М. (2012). «Биологическая продуктивность океана» (PDF). Знание природы образования. 3 (6): 1–16. Получено 2015-06-01.
Глубокий максимум хлорофилла (DCM) происходит в месте контакта, где есть достаточно света для фотосинтеза и при этом значительный приток питательных веществ снизу.
- ^ а б Mora, C .; и другие. (2013). "Уязвимость биотики и человека к прогнозируемым изменениям в биогеохимии океана в 21 веке". PLOS Биология. 11 (10): e1001682. Дои:10.1371 / journal.pbio.1001682. ЧВК 3797030. PMID 24143135.
- ^ Martin, J. H .; Фитцуотер, С. Э. (1988). «Дефицит железа ограничивает рост фитопланктона в северо-восточной части Тихого океана в Субарктике». Природа. 331 (6154): 341–343. Bibcode:1988Натура.331..341М. Дои:10.1038 / 331341a0.
- ^ Купер, Д.Дж .; Watson, A.J .; Найтингейл, П. (1996). "Большое уменьшение океана - поверхность CO
2 легкомыслие в ответ на на месте удобрение железом ». Природа. 383 (6600): 511–513. Bibcode:1996Натура.383..511C. Дои:10.1038 / 383511a0. - ^ а б c Clark, D.A .; Brown, S .; Kicklighter, D.W .; Chambers, J.Q .; Thomlinson, J.R .; Ни, Дж. (2001). «Измерение чистой первичной продукции в лесах: концепции и полевые методы». Экологические приложения. 11 (2): 356–370. Дои:10.1890 / 1051-0761 (2001) 011 [0356: MNPPIF] 2.0.CO; 2. ISSN 1051-0761.
- ^ а б c Scurlock, J.M.O .; Johnson, K .; Олсон, Р.Дж. (2002). «Оценка чистой первичной продуктивности на основе измерений динамики биомассы пастбищ». Биология глобальных изменений. 8 (8): 736–753. Bibcode:2002GCBio ... 8..736S. Дои:10.1046 / j.1365-2486.2002.00512.x.
- ^ Leith, H .; Уиттакер, Р. Х. (1975). Первичная продуктивность биосферы. Нью-Йорк: Springer-Verlag. ISBN 978-0-387-07083-4.
- ^ Райхштейн, Маркус; Фальге, Ева; Балдокки, Деннис; Папале, Дарио; Обине, Марк; Бербиджье, Поль; и другие. (2005). «О разделении сетевого обмена экосистемами на ассимиляцию и экосистемное дыхание: обзор и улучшенный алгоритм». Биология глобальных изменений. 11 (9): 1424–1439. Bibcode:2005GCBio..11.1424R. Дои:10.1111 / j.1365-2486.2005.001002.x. ISSN 1354-1013.
- ^ Марра, Дж. (2002), стр. 78-108. В: Уильямс, П. Дж. ЛеБ., Томас, Д. Н., Рейнольдс, С. С. (ред.), Продуктивность фитопланктона: Ассимиляция углерода в морских и пресноводных экосистемах. Блэквелл, Оксфорд, Великобритания
- ^ Стиман-Нильсен, Э. (1951). «Измерение продукции органического вещества в море с помощью углерода-14». Природа. 167 (4252): 684–685. Bibcode:1951Натура.167..684Н. Дои:10.1038 / 167684b0. PMID 14826912.
- ^ Стиман-Нильсен, Э. (1952). «Использование радиоактивного углерода (C14) для измерения органического производства в море». J. Cons. Int. Explor. Мер. 18 (2): 117–140. Дои:10.1093 / icesjms / 18.2.117.
- ^ Бендер, Майкл; и другие. (1987). «Сравнение 4 методов определения продукции планктонного сообщества». Лимнология и океанография. 32 (5): 1085–1098. Bibcode:1987LimOc..32.1085B. Дои:10.4319 / lo.1987.32.5.1085.
- ^ Луз и Баркан, Б; Баркан, Э (2000). «Оценка продуктивности океана по тройному изотопному составу растворенного кислорода». Наука. 288 (5473): 2028–2031. Bibcode:2000Sci ... 288.2028L. Дои:10.1126 / science.288.5473.2028. PMID 10856212.
- ^ Карвалью и Эйр (2012). «Измерение планктонного дыхания СО2 на свету». Лимнология и океанография: методы. 10 (3): 167–178. Дои:10.4319 / lom.2012.10.167.
- ^ Крейг и Хейворд (1987). «Перенасыщение кислородом в океане: биологический и физический вклад». Наука. 235 (4785): 199–202. Bibcode:1987 Наука ... 235..199C. Дои:10.1126 / science.235.4785.199. PMID 17778634.
- ^ Cassar, N .; Б.А. Барнетт; М.Л. Бендер; Дж. Кайзер; R.C. Хамме; Б. Тилбрук (2009). «Непрерывный высокочастотный растворенный O2/ Измерение Ar методом масс-спектрометрии на входе в уравновешивающий агрегат » Анальный. Chem. 81 (5): 1855–1864. Дои:10.1021 / ac802300u. PMID 19193192.
- ^ Kaiser, J .; М. К. Ройер; Б. Барнетт; М.Л. Бендер (2005). «Оценка продуктивности морской среды на основе непрерывных измерений отношения O-2 / Ar с помощью масс-спектрометрии на входе мембраны». Geophys. Res. Латыш. 32 (19): н / д. Bibcode:2005GeoRL..3219605K. CiteSeerX 10.1.1.584.1430. Дои:10.1029 / 2005GL023459.
- ^ а б Филд, C.B .; Behrenfeld, M.J .; Randerson, J.T .; Фальковски, П. (1998). «Первичная продукция биосферы: интеграция компонентов суши и океана». Наука. 281 (5374): 237–240. Bibcode:1998Научный ... 281..237F. Дои:10.1126 / science.281.5374.237. PMID 9657713.
- ^ Пиво, гр .; Reichstein, M .; Tomelleri, E .; Ciais, P .; Юнг, М .; Carvalhais, N .; и другие. (2010). «Валовое поглощение двуокиси углерода землей: глобальное распределение и ковариация с климатом» (PDF). Наука. 329 (5993): 834–838. Bibcode:2010Sci ... 329..834B. Дои:10.1126 / science.1184984. ISSN 0036-8075. PMID 20603496.
- ^ Пракаш Бабуа, C .; Brumsack, H.-J .; Schnetger, B .; Бёттчер, M.E. (2002). «Барий как показатель продуктивности в отложениях континентальной окраины: исследование восточной части Аравийского моря». Морская геология. 184 (3–4): 189–206. Bibcode:2002МГеол.184..189П. Дои:10.1016 / S0025-3227 (01) 00286-9.
- ^ E.D. Гольдберг, G.O.S. Аррениус Химия пелагических отложений Геохим. Cosmochim. Acta, 13 (1958), стр. 153-212
- ^ Даймонд, Джек; Сьюсс, Эрвин; Лайл, Митч (1992). «Барий в глубоководных отложениях: геохимический показатель палеопродуктивности» (PDF). Палеоокеанография. 7 (2): 163–181. Bibcode:1992ПалОк ... 7..163Д. Дои:10.1029 / 92PA00181.
- ^ а б Vitousek, P.M .; Эрлих, П.Р .; Ehrlich, A.H .; Матсон, П. (1986). «Присвоение человеком продуктов фотосинтеза» (PDF). Бионаука. 36 (6): 368–373. Дои:10.2307/1310258. JSTOR 1310258. Архивировано из оригинал (PDF) на 2011-03-05.
- ^ Гари, J.A. (1996). «HANPP рассчитывается по растительному покрову как индикатор экологической устойчивости». Материалы Международной конференции: Экология, общество, экономика.: 1–18. HDL:10419/183215.
- ^ а б c Haberl, H .; Erb, K.H .; Krausmann, F .; Gaube, V .; Bondeau, A .; Plutzar, C .; Gingrich, S .; Lucht, W .; Фишер-Ковальский, М. (2007). «Количественная оценка и составление карты освоения человеком чистой первичной продукции в наземных экосистемах Земли». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 104 (31): 12942–12947. Bibcode:2007PNAS..10412942H. Дои:10.1073 / pnas.0704243104. ЧВК 1911196. PMID 17616580.
- ^ Ramankutty, N .; Эван, A.T .; Monfreda, C .; Фоли, Дж. (2008). «Сельское хозяйство на планете: 1. Географическое распределение сельскохозяйственных земель в мире в 2000 году». Глобальные биогеохимические циклы. 22 (1): GB1003. Bibcode:2008GBioC..22.1003R. Дои:10.1029 / 2007GB002952.
- ^ Foley, J.A .; Monfreda, C .; Ramankutty, N .; Закс, Д. (2007). «Наша доля планетарного пирога». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 104 (31): 12585–12586. Bibcode:2007ПНАС..10412585F. Дои:10.1073 / pnas.0705190104. ЧВК 1937509. PMID 17646656.