Водоросли - Seagrass

Морские травы являются (единственными) цветущие растения которые растут в морской среды. Существует около 60 видов полностью морских водорослей, которые относятся к четырем семьи (Posidoniaceae, Zosteraceae, Hydrocharitaceae и Cymodoceaceae ), все по порядку Двулистные (в классе однодольные ).[1] Морские травы произошли от наземные растения который повторно заселил океан 70–100 миллионов лет назад.
Название водоросли Стебли от многих видов с длинными и узкими листьями, которые растут за счет расширения корневища и часто распространяются на большие "луга "напоминающие луга; многие виды внешне напоминают земных травы семьи Poaceae.
Как все автотрофный растения, морские травы фотосинтезировать, в затопленном фотическая зона, и большинство из них встречается в неглубоких и защищенных прибрежных водах, закрепленных на песчаном или иловом дне. Большинство видов подвергаются подводным опыление и завершают свой жизненный цикл под водой.
Морские травы образуют плотную под водой водоросли луга которые являются одними из самых продуктивных экосистем в мире. Они функционируют как важные поглотители углерода и обеспечить среду обитания и пищу для разнообразных морская жизнь сравнимо с коралловые рифы.
Эволюция

Наземные растения развились, возможно, уже 450 миллионов лет назад из группы зеленые водоросли.[2] Затем морские травы произошли от наземных растений, которые мигрировали обратно в океан.[3][4] Примерно 70-100 миллионов лет назад три независимые линии морских водорослей (Hydrocharitaceae, Cymodoceaceae complex и Zosteraceae) произошли от одной линии однодольные цветущие растения.[5]
Другие растения, колонизировавшие море, такие как болото, периодически затопляемое морской водой растения мангровые заросли, и морские водоросли, имеют более разнообразные эволюционные линии. Несмотря на свое низкое видовое разнообразие, морские травы смогли колонизировать континентальные шельфы всех континентов, кроме Антарктиды.[6]
Таксономия
| Семья | Изображение | Роды | Описание |
|---|---|---|---|
| Zosteraceae | Семья Zosteraceae, также известный как семейство водорослей, включает два рода, включающих 14 морских видов. Он находится в умеренный и субтропический прибрежный воды, с наибольшим разнообразием в Корее и Японии. Итого по видам: | ||
 | Филлоспадикс | 6 видов | |
 | Зостера | 16 видов | |
| Hydrocharitaceae | Семья Hydrocharitaceae, также известный как ленточные травы, включают Канадский водоросль и лягушонок. В семейство входят как пресные, так и морские водные виды, хотя из шестнадцати известных в настоящее время родов только три являются морскими. [7] Они встречаются по всему миру в самых разных средах обитания, но в основном тропические. Итого по видам: | ||
 | Энхалус | 1 вид | |
 | Halophila | 19 видов | |
 | Талассия | 2 вида | |
| Posidoniaceae | Семья Posidoniaceae содержит один род от двух до девяти морских видов, обитающих в морях Средиземноморье и вокруг южного побережья Австралия. Итого по видам: от 2 до 9 | ||
 | Посидония | От 2 до 9 видов | |
| Cymodoceaceae | Семья Cymodoceaceae, также известный как ламантин, включает только морские виды.[8] Некоторые систематики не признают это семейство. Итого по видам: | ||
 | Амфиболис | 2 вида | |
 | Cymodocea | 4 вида | |
 | Halodule | 6 видов | |
 | Syringodium | 2 вида | |
 | Талассодендрон | 3 вида | |
| Всего видов: | |||
Приливные и сублиторальные водоросли
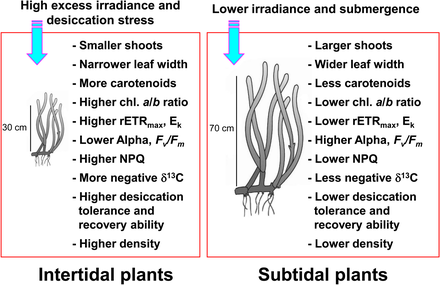
Морские травы, встречающиеся в приливной и сублиторальной зонах, подвергаются очень изменчивым условиям окружающей среды из-за приливных изменений.[10][11] Морские травы в приливная зона регулярно подвергаются воздействию воздуха и, как следствие, подвергаются экстремально высоким и низким температурам, высоким световым ингибиторам сияние, и высыхание стресс по сравнению с сублиторальными водорослями.[11][12][13] Такие экстремальные температуры могут привести к значительному усыханию водорослей, когда они подвергаются воздействию воздуха во время отлива.[14][15][16] Напряжение высыхания во время отлива считается основным фактором, ограничивающим распространение водорослей в верхней приливной зоне.[17] Морские травы, обитающие в приливной зоне, обычно меньше, чем в сублиторальной зоне, чтобы свести к минимуму воздействие стресса от появления всходов.[18][19] Морские травы в приливной зоне также демонстрируют светозависимые реакции, такие как снижение фотосинтетической эффективности и усиление светозащиты в периоды высокой освещенности и воздействия воздуха.[20][21]

Напротив, морские травы в сублиторальная зона адаптироваться к условиям пониженного освещения, вызванным ослаблением и рассеянием света из-за перекрывающего водяного столба и взвешенных частиц.[23][24] Морские травы в глубокой сублиторальной зоне обычно имеют более длинные листья и более широкие листовые пластинки, чем те, что находятся в неглубокой сублиторальной зоне или приливной зоне, что способствует большему фотосинтезу, что, в свою очередь, приводит к большему росту.[13] Морские травы также реагируют на снижение освещенности увеличением хлорофилл содержание и уменьшение соотношение хлорофилла a / b усилить поглощение света эффективность за счет эффективного использования большого количества длин волн.[25][26][27] Поскольку морские травы в приливной и сублиторальной зонах находятся в очень разных условиях освещения, они демонстрируют совершенно разные фотоаклиматические реакции, чтобы максимизировать фотосинтетическую активность и светозащиту от избыточного излучения.
Морские травы усваивают большое количество неорганический углерод для достижения высокого уровня производства.[28][29] Морские макрофиты, включая водоросли, используйте CO2 и HCO−
3 (бикарбонат ) для фотосинтетического восстановления углерода.[30][31][32] Несмотря на воздействие воздуха во время отлива, морские травы в приливной зоне могут продолжать фотосинтез, используя CO.2 в воздухе.[33] Таким образом, состав источников неорганического углерода для фотосинтеза морских водорослей, вероятно, варьируется между приливными и сублиторальными растениями. Потому что стабильный отношения изотопов углерода изменения тканей растений на основе источников неорганического углерода для фотосинтеза,[34][35] Морские травы в приливной и сублиторальной зонах могут иметь разные диапазоны соотношений стабильных изотопов углерода.
Микробиом водорослей
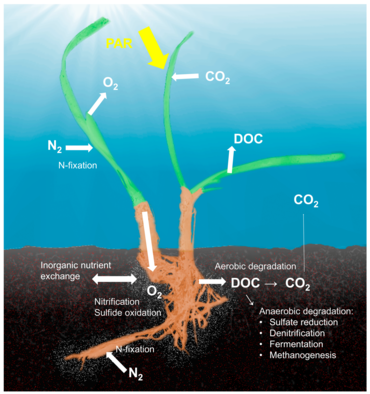
Морской холобионт
Концепция холобионт, который подчеркивает важность и взаимодействие микробного хозяина с ассоциированными микроорганизмами и вирусами и описывает их функционирование как единую биологическую единицу,[38] был исследован и обсужден для многих модельных систем, хотя есть существенная критика концепции, которая определяет различные симбиозы хозяина и микроба как единую биологическую единицу.[39] Концепции холобионта и гологенома эволюционировали с момента первоначального определения,[40] и нет сомнений в том, что симбиотические микроорганизмы имеют решающее значение для биологии и экологии хозяина, обеспечивая витамины, энергию и неорганические или органические питательные вещества, участвуя в защитных механизмах или управляя эволюцией хозяина.[41] Хотя большая часть работ по взаимодействию хозяев и микробов была сосредоточена на системах животных, таких как кораллы, губки или люди, существует значительный объем литературы по холобионтам растений.[42] Связанные с растениями микробные сообщества влияют на оба ключевых компонента приспособленности растений, роста и выживания,[43] и формируются доступностью питательных веществ и защитными механизмами растений.[44] Было описано несколько мест обитания микробов, связанных с растениями, включая ризоплан (поверхность корневой ткани), ризосфера (периферия корней), эндосфера (внутри растительной ткани) и филлосфера (общая площадь надземной поверхности).[36]
Луга с водорослями
Грядки из водорослей / Луга могут быть как моноспецифичными (состоящими из одного вида), так и смешанными. В умеренный области, обычно преобладают один или несколько видов (например, угорь Зостера Марина в Северной Атлантике), тогда как тропический кровати обычно более разнообразные, до тринадцати разновидность записано в Филиппины.
Грядки из водорослей разнообразны и продуктивны экосистемы, и может служить приютом для сотен ассоциированных видов со всех тип, например подростки и взрослые рыбы, эпифитный и свободная жизнь макроводоросли и микроводоросли, моллюски, щетинистые черви, и нематоды. Первоначально считалось, что немногие виды питаются непосредственно водорослями. листья (частично из-за их низкого содержания питательных веществ), но научные обзоры и улучшенные методы работы показали, что водоросли травоядный является важным звеном в пищевой цепи, питая сотни видов, в том числе зеленые черепахи, дюгони, ламантины, рыбы, гуси, лебеди, морские ежи и крабы. Некоторые виды рыб, которые посещают / питаются водорослями, выращивают молодь в прилегающих мангровые заросли или же коралловые рифы.
Морская трава улавливает отложения и замедляет движение воды, вызывая осаждение взвешенных отложений. Преимущества улавливания осадка коралл за счет уменьшения наносов и улучшения фотосинтеза как кораллов, так и морских водорослей.[45]

Пуховики с белыми пятнами, часто встречается в зонах водорослей
Подводные кадры луга из водорослей, бык и морской угорь





Хотя морские травы часто упускаются из виду, они обеспечивают ряд экосистемные услуги[46][47]. Морские травы считаются инженеры экосистемы.[48][4][3] Это означает, что растения изменяют экосистему вокруг себя. Это регулирование происходит как в физической, так и в химической форме. Многие виды морских водорослей образуют обширную подземную сеть корней и корневище который стабилизирует отложения и снижает береговую эрозию.[49] Эта система также помогает насыщать осадок кислородом, обеспечивая гостеприимную среду для организмы, обитающие в донных отложениях.[48] Морские травы также улучшают качество воды за счет стабилизации тяжелых металлов, загрязнителей и избыточных питательных веществ.[50][4][3] Длинные лезвия морской травы замедляют движение воды, что снижает энергию волн и обеспечивает дополнительную защиту от прибрежных вод. эрозия и штормовая волна. Кроме того, поскольку морские травы являются подводными растениями, они производят значительное количество кислорода, насыщающего водную толщу кислородом. На эти луга приходится более 10% всех запасов углерода в океане. На гектар в нем содержится вдвое больше углекислого газа, чем в тропических лесах, и может улавливаться около 27,4 миллиона тонн CO.2 ежегодно.[51] Хранение углерода является важным экосистемная услуга по мере того, как мы вступаем в период повышенного уровня углерода в атмосфере. Однако некоторые модели изменения климата предполагают, что некоторые водоросли вымрут - Посидония океаническая ожидается, что к 2050 году оно исчезнет или почти исчезнет.
Луга с водорослями обеспечивают пищу многим морским травоядным животным. Морские черепахи, ламантины, рыбы-попугаи, хирурги, морские ежи и сосны питаются морской травой. Многие другие более мелкие животные питаются эпифитами и беспозвоночными, которые живут на стеблях водорослей и среди них.[52] Луга с водорослями также обеспечивают физическую среду обитания в областях, которые в противном случае были бы лишены какой-либо растительности. Из-за этой трехмерной структуры в водной толще многие виды занимают места обитания морских водорослей для укрытия и кормления. Подсчитано, что 17 видов рыб коралловых рифов проводят всю свою молодую стадию жизни исключительно на равнинах с водорослями.[53] Эти места обитания также служат рассадником коммерчески и рекреационных видов рыбного промысла, в том числе морского окуня (Mycteroperca microlepis ), красный барабан, обыкновенный снук, и много других.[54][55] Некоторые виды рыб используют луга из морских водорослей и используют различные стадии жизненного цикла. В недавней публикации д-р Росс Боучек и его коллеги обнаружили, что две популярные плоские рыбы: снук и пятнистая морская форель обеспечивают необходимую среду обитания для кормления во время размножения.[56] Половое размножение чрезвычайно затратно с точки зрения энергии, чтобы его завершить за счет накопленной энергии; поэтому для полного воспроизводства им требуются луга морских водорослей в непосредственной близости.[56] Кроме того, многие коммерчески важные беспозвоночные также обитают в местах обитания морских водорослей, включая гребешки (Argopecten irradians ), подковообразные крабы, и креветка. Харизматичную фауну можно увидеть и в местах обитания морских водорослей. Эти виды включают Вест-индийский ламантин, зеленые морские черепахи, и различные виды акул. Большое разнообразие морских организмов, встречающихся в местах обитания водорослей, делает их туристической достопримечательностью и значительным источником дохода для многих прибрежных стран вдоль Мексиканского залива и в Карибском бассейне.
Отношение к людям
Исторически морские травы собирали как удобрение для песчаной почвы. Это было важное использование в Лагуна Авейру, Португалия, где собранные растения были известны как Moliço.
В начале 20 века во Франции и, в меньшей степени, в Нормандские острова, сушеные морские травы использовались в качестве матраса (Paillasse) наполнение - такие матрасы пользовались большим спросом у французских войск во время Первая Мировая Война. Его также использовали для перевязок и других целей.
В феврале 2017 года исследователи обнаружили, что луга с водорослями могут выводить из морской воды различные патогены. На небольших островах без очистных сооружений в центральной Индонезии уровни патогенных морские бактерии - Такие как Энтерококк - которые влияют на людей, рыбу и беспозвоночные были сокращены на 50 процентов при наличии лугов с водорослями по сравнению с парными участками без водорослей,[57] хотя это могло нанести ущерб их выживанию.[58]
Нарушения и угрозы
Естественные нарушения, такие как выпас, штормы, очистка льда и высыхание, являются неотъемлемой частью морских водорослей экосистема динамика. Морские травы демонстрируют высокую степень фенотипическая пластичность, быстро приспосабливаясь к изменяющимся условиям окружающей среды.
Морские травы находятся в упадке, их протяженность составляет около 30 000 км.2 (12 000 квадратных миль), потерянных за последние десятилетия. Основная причина - человеческое беспокойство, прежде всего эвтрофикация, механическое разрушение среды обитания, и перелов. Чрезмерное поступление питательных веществ (азот, фосфор ) напрямую токсичен для морских трав, но, что наиболее важно, он стимулирует рост эпифитных и свободно плавающих макрос - и микро -водоросли. Это ослабляет Солнечный свет, уменьшая фотосинтез питает водоросли и основное производство полученные результаты.
Разлагающиеся листья водорослей и водоросли увеличиваются цветение водорослей, что привело к положительному Обратная связь. Это может вызвать полное смена режима от водорослей до преобладания водорослей. Накапливающиеся данные также предполагают, что перелов верхних хищники (крупная хищная рыба) может косвенно увеличить рост водорослей за счет снижения контроля над выпасом, осуществляемого мезографы, Такие как ракообразные и брюхоногие моллюски через трофический каскад.
Цветение макроводорослей вызывают упадок и искоренение морских трав. Известные как вредные виды, макроводоросли растут в нитевидных и листовых формах и образуют толстые неприкрепленные маты над водорослями, появляясь в виде эпифитов на листьях водорослей. Эвтрофикация приводит к образованию цветков, вызывая ослабление света в толще воды, что в конечном итоге приводит к бескислородным условиям для морских водорослей и организмов, живущих в / вокруг растений. В дополнение к прямой блокировке света для растений, бентосные макроводоросли имеют низкое содержание углерода / азота, что приводит к их разложению и стимулированию бактериальной активности, что приводит к повторному суспендированию отложений, увеличению мутности воды и дальнейшему ослаблению света.[59][60]
Когда люди водят моторные лодки по мелководным участкам водорослей, иногда лопасти гребного винта могут повредить водоросли.
Наиболее часто используемые методы защиты и восстановления лугов с водорослями включают использование питательных веществ и загрязнение снижение, морские охраняемые территории и реставрация с использованием водорослей пересадка. Морские водоросли не считаются устойчивыми к воздействиям будущих изменений окружающей среды.[61]
Реставрация
В разных местах сообщества пытаются восстановить заросли водорослей, утраченные в результате действий человека, в том числе в штатах Вирджиния США,[62] Флорида[63] и Гавайи,[64] а также Соединенное Королевство.[65] Было показано, что такие реинтродукции улучшают экосистемные услуги.[66]
По состоянию на 2019 год[Обновить] Центр исследования прибрежных морских экосистем Центральный университет Квинсленда выращивает водоросли шесть лет и производит семена водорослей. Были проведены испытания техники проращивания и посева.[67]
Смотрите также
- Двулистные
- Синий углерод
- Болото, периодически затопляемое морской водой
- Мангровые заросли
- Средство просмотра океанографических данных: содержит набор данных о глобальном распространении морских водорослей
Рекомендации
- ^ Томлинсон и Варго (1966). «О морфологии и анатомии черепаховой травы Thalassia testudinum (Hydrocharitaceae). I. Вегетативная морфология». Бюллетень морской науки. 16: 748–761.
- ^ Кнаут, Л. Пол; Кеннеди, Мартин Дж. (2009). «Позднедокембрийское озеленение Земли». Природа. 460 (7256): 728–732. Bibcode:2009Натура.460..728K. Дои:10.1038 / природа08213. PMID 19587681. S2CID 4398942.
- ^ а б c Орт; и другие. (2006). «Глобальный кризис экосистемы морских водорослей». Бионаука. 56 (12): 987–996. Дои:10.1641 / 0006-3568 (2006) 56 [987: AGCFSE] 2.0.CO; 2. HDL:10261/88476.
- ^ а б c Папенброк, Дж (2012). «Основные моменты филогении, физиологии и метаболизма морских водорослей: что делает их такими видами?». Международная сеть научных исследований: 1–15.
- ^ Лес, Д.Х., Клеланд, М.А. и Уэйкотт, М. (1997) "Филогенетические исследования Alismatidae, II: эволюция морских покрытосеменных (морских трав) и гидрофилия". Систематическая ботаника 22(3): 443–463.
- ^ Орт, Роберт Дж .; Carruthers, TIM J. B .; Деннисон, Уильям С .; Дуарте, Карлос М .; Fourqurean, Джеймс У .; Черт возьми, Кеннет Л .; Хьюз, А. Рэндалл; Кендрик, Гэри А .; Кенуорти, У. Джадсон; Олярник, Сюзанна; Коротко, Фредерик Т .; Уэйкотт, Мишель; Уильямс, Сьюзан Л. (2006). «Глобальный кризис экосистем водорослей». Бионаука. 56 (12): 987. Дои:10.1641 / 0006-3568 (2006) 56 [987: AGCFSE] 2.0.CO; 2. ISSN 0006-3568.
- ^ Christenhusz, Maarten J.M .; Бинг, Джеймс У. (20 мая 2016 г.). «Количество известных видов растений в мире и его ежегодный прирост». Фитотакса. 261 (3): 201. Дои:10.11646 / phytotaxa.261.3.1. ISSN 1179-3163.
- ^ Уэйкотт, Мишель; МакМахон, Кэтрин; Лавери, Пол (2014). Путеводитель по южным водорослям умеренного пояса. CSIRO Publishing. ISBN 9781486300150.
- ^ Парк, Санг Рул; Ким, Сангил; Ким, Ён Кюн; Канг, Чанг-Гын; Ли, Кун-Сеоп (2016). «Фотоаклиматические реакции Zostera marina в приливной и сублиторальной зонах». PLOS ONE. 11 (5): e0156214. Bibcode:2016PLoSO..1156214P. Дои:10.1371 / journal.pone.0156214. ЧВК 4881947. PMID 27227327.
 Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0. - ^ Silva, J .; Сантос, Р. (2003). «Суточные вариации фотосинтеза водорослей по вертикальному градиенту». Серия "Прогресс морской экологии". 257: 37–44. Bibcode:2003MEPS..257 ... 37S. Дои:10.3354 / meps257037.
- ^ а б Boese, Bruce L .; Роббинс, Брэдли Д.; Терзби, Глен (2005). «Осушение является ограничивающим фактором для распространения угря (Zostera marina L.) в приливной зоне эстуария северо-востока Тихого океана (США)». Ботаника Марина. 48 (4). Дои:10.1515 / BOT.2005.037. S2CID 85105171.
- ^ Дурако, М. Дж .; Kunzelman, J. I .; Kenworthy, W. J .; Хаммерстром, К. К. (2003). «Связанная с глубиной изменчивость в фотобиологии двух популяций Halophila johnsonii и Halophila decipiens». Морская биология. 142 (6): 1219–1228. Дои:10.1007 / s00227-003-1038-3. S2CID 85627116.
- ^ а б Olivé, I .; Vergara, J. J .; Перес-Льоренс, Дж. Л. (2013). «Фотосинтетическая и морфологическая фотоакклимация водорослей Cymodocea nodosa в зависимости от сезона, глубины и положения листьев». Морская биология. 160 (2): 285–297. Дои:10.1007 / s00227-012-2087-2. S2CID 86386210.
- ^ Хемминга М. А. и Дурате К. М. (2000) Экология водорослей. Издательство Кембриджского университета.
- ^ Seddon, S .; Чешир, AC (2001). «Фотосинтетический ответ Amphibolis antarctica и Posidonia australis на температуру и высыхание с использованием флуоресценции хлорофилла». Серия "Прогресс морской экологии". 220: 119–130. Bibcode:2001MEPS..220..119S. Дои:10.3354 / meps220119.
- ^ Херст А., Болл Д., Хейслерс С., Янг П., Блейк С., Кутс А. Программа мониторинга морских водорослей Бэйуэйда, Milestone Report No. 2 (2008). Технический отчет Fisheries Victoria No. 29, январь 2009 г.
- ^ Кох, Эвамария В. (2001). «За пределами света: физические, геологические и геохимические параметры как возможные требования к среде обитания подводной водной растительности». Эстуарии. 24 (1): 1–17. Дои:10.2307/1352808. JSTOR 1352808. S2CID 85287808.
- ^ Tanaka, Y .; Накаока, М. (2004). «Экстресс и морфологические ограничения влияют на распространение видов и рост субтропических приливных водорослей». Серия "Прогресс морской экологии". 284: 117–131. Bibcode:2004MEPS..284..117T. Дои:10,3354 / meps284117.
- ^ Seddon, S .; Чешир, AC (2001). «Фотосинтетический ответ Amphibolis antarctica и Posidonia australis на температуру и высыхание с использованием флуоресценции хлорофилла». Серия "Прогресс морской экологии". 220: 119–130. Bibcode:2001MEPS..220..119S. Дои:10.3354 / meps220119.
- ^ Björk, M .; Uku, J .; Weil, A .; Бир, С. (1999). «Фотосинтетическая устойчивость к высыханию тропических приливных водорослей». Серия "Прогресс морской экологии". 191: 121–126. Bibcode:1999MEPS..191..121B. Дои:10,3354 / meps191121.
- ^ Петру, К .; Jimenez-Denness, I .; Chartrand, K .; McCormack, C .; Рашид, М .; Ральф, П.Дж. (2013). «Сезонная неоднородность фотофизиологической реакции на воздействие воздуха у двух тропических литоральных видов морских водорослей» (PDF). Серия "Прогресс морской экологии". 482: 93–106. Bibcode:2013MEPS..482 ... 93P. Дои:10,3354 / meps10229.
- ^ Сюй, Шаочунь; Чжоу, Йи; Ван, Пэнмэй; Ван, Фэн; Чжан, Сяомэй; Гу, Рутинг (2016). «Засоление и температура существенно влияют на прорастание семян, укоренение рассады и рост рассады угря Зостера маринаЛ ". PeerJ. 4: e2697. Дои:10.7717 / peerj.2697. ЧВК 5119234. PMID 27896031. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ Schwarz, A.-M .; Björk, M .; Булуда, Т .; Мтолера, М .; Бир, С. (2000). «Фотосинтетическое использование углерода и света двумя видами тропических водорослей, измеренное на месте». Морская биология. 137 (5–6): 755–761. Дои:10.1007 / s002270000433. S2CID 86384408.
- ^ Кэмпбелл, Стюарт Дж .; Маккензи, Лен Дж .; Kerville, Simon P .; Бите, Хуанита С. (2007). «Закономерности фотосинтеза тропических водорослей в зависимости от света, глубины и среды обитания». Эстуарии, прибрежные зоны и шельфовые науки. 73 (3–4): 551–562. Bibcode:2007 ECSS ... 73..551C. Дои:10.1016 / j.ecss.2007.02.014.
- ^ Ли, Кун-Сеоп; Дантон, Кеннет Х. (1997). «Влияние уменьшения освещенности на месте на поддержание, рост и распределение углеродных ресурсов в банках Thalassia testudinum ex König». Журнал экспериментальной морской биологии и экологии. 210: 53–73. Дои:10.1016 / S0022-0981 (96) 02720-7.
- ^ Лонгстафф, Б.Дж .; Деннисон, W.C (1999). «Выживание морских водорослей во время событий импульсного помутнения: влияние отсутствия света на водоросли Halodule pinifolia и Halophila ovalis». Водная ботаника. 65 (1–4): 105–121. Дои:10.1016 / S0304-3770 (99) 00035-2.
- ^ Кольер, CJ; Лавери, PS; Ральф, П.Дж.; Масини, RJ (2008). «Физиологические характеристики водорослей Posidonia sinuosa в зависимости от глубины градиента доступности света». Серия "Прогресс морской экологии". 353: 65–79. Bibcode:2008MEPS..353 ... 65C. Дои:10.3354 / meps07171.
- ^ Ли, Кун-Сеоп; Парк, Санг Рул; Ким, Ён Кюн (2007). «Влияние освещенности, температуры и питательных веществ на динамику роста морских трав: обзор». Журнал экспериментальной морской биологии и экологии. 350 (1–2): 144–175. Дои:10.1016 / j.jembe.2007.06.016.
- ^ Nayar, S .; Collings, G.J .; Миллер, Д.Дж .; Bryars, S .; Чешир, A.C. (2009). «Поглощение и распределение ресурсов неорганического углерода морскими травами умеренного пояса Посидония и Амфиболис». Журнал экспериментальной морской биологии и экологии. 373 (2): 87–95. Дои:10.1016 / j.jembe.2009.03.010.
- ^ Пиво, Свен (1989). «Фотосинтез и фотодыхание морских покрытосеменных». Водная ботаника. 34 (1–3): 153–166. Дои:10.1016/0304-3770(89)90054-5.
- ^ Ларкум AWD, Джеймс П.Л. К модели поглощения неорганического углерода водорослями с участием карбоангидразы. В Kuo J, Phillips RC, Walker DI, Kirkman H, редакторы. Биология морских водорослей: материалы международного семинара. Недлендс: Университет Западной Австралии; 1996. С. 191–196.
- ^ Пиво, Свен; Ренберг, Джон (1997). «Приобретение неорганического углерода водорослями Zostera marina». Водная ботаника. 56 (3–4): 277–283. Дои:10.1016 / S0304-3770 (96) 01109-6.
- ^ Сильва, Жоао; Сантос, Руи; Calleja, Maria Ll .; Дуарте, Карлос М. (2005). «Затопленная продуктивность по сравнению с продуктивностью приливных макрофитов при воздействии воздуха: от физиологических оценок до оценок на уровне сообществ». Журнал экспериментальной морской биологии и экологии. 317: 87–95. Дои:10.1016 / j.jembe.2004.11.010.
- ^ О'Лири, Мэрион Х. (1988). «Изотопы углерода в фотосинтезе». Бионаука. 38 (5): 328–336. Дои:10.2307/1310735. JSTOR 1310735.
- ^ Рэйвен, Джон А .; Джонстон, Эндрю М .; Kübler, Janet E .; Корб, Ребекка; Макинрой, Шона Дж .; Handley, Linda L .; Скримджер, Чарли М .; Уокер, Диана I .; Бирдалл, Джон; Вандерклифт, Мэтью; Фредриксен, Штейн; Дантон, Кеннет Х. (2002). «Механистическая интерпретация дискриминации изотопов углерода морскими макроводорослями и водорослями». Функциональная биология растений. 29 (3): 355–378. Дои:10.1071 / PP01201. PMID 32689482.
- ^ а б Угарелли К., Чакрабарти С., Лаас П. и Стингл У. (2017) «Холобионт из морских водорослей и его микробиом». Микроорганизмы, 5(4): 81. Дои:10.3390 / микроорганизмы 5040081. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ Tarquinio, F., Hyndes, G.A., Laverock, B., Koenders, A. и Säwström, C. (2019) «Холобионт из морских водорослей: понимание взаимодействия морских водорослей и бактерий и их роли в функционировании экосистемы морских водорослей». Письма по микробиологии FEMS, 366(6): fnz057. Дои:10.1093 / femsle / fnz057.
- ^ Маргулис, Линн (1991) «Симбиогенез и симбионтизм». В: Симбиоз как источник эволюционных инноваций; Маргулис, Л., Фестер, Р. (ред.), Cambridge MIT Press. ISBN 9780262132695.
- ^ Дуглас, A.E .; Веррен, Дж. (2016) «Дыры в гологеноме: почему симбиозы микробов-хозяев не являются голобионтами». мБио, 7: e02099-15. Дои:10.1128 / mBio.02099-15.
- ^ Theis, K.R .; Dheilly, N.M .; Klassen, J.L .; Brucker, R.M .; Baines, J.F .; Bosch, T.C.G .; Cryan, J.F .; Gilbert, S.F .; Спокойной ночи, Си-Джей; Lloyd, E.A .; и другие. Правильная концепция гологенома: эко-эволюционная структура для хозяев и их микробиомов. mSystems 2016, 1, e00028-16. Дои:10.1128 / м Системы.00028-16.
- ^ Розенберг, Э. и Зильбер-Розенберг, И.(2016) «Микробы двигают эволюцию животных и растений: концепция гологенома». MBio, 7(2). Дои:10,1128 / мБио.01395-15.
- ^ Зильбер-Розенберг, И. и Розенберг, Э. (2008) "Роль микроорганизмов в эволюции животных и растений: теория эволюции гологенома". Обзор микробиологии FEMS, 32(5): 723–735. Дои:10.1111 / j.1574-6976.2008.00123.x.
- ^ Ванденкоорнхейз П., Квайзер А., Дюамель М., Ле Ван А. и Дюфрен А. (2015) «Важность микробиома холобионта растения». Новый Фитолог, 206(4): 1196-1206. Дои:10.1111 / nph.13312.
- ^ Санчес-Каньисарес, К., Хоррин, Б., Пул, П.С. и Ткач А. (2017) «Понимание холобионта: взаимозависимость растений и их микробиома». Текущее мнение в микробиологии, 38: 188–196. Дои:10.1016 / j.mib.2017.07.001.
- ^ Seagrass-Watch: Что такое водоросли? Проверено 16 ноября 2012.
- ^ Нордлунд, Лина; Koch, Evamaria W .; Барбье, Эдвард Б .; Крид, Джоэл С. (12 октября 2016 г.). Рейнхарт, Курт О. (ред.). «Экосистемные услуги морских водорослей и их изменчивость в разных родах и географических регионах». PLOS ONE. 11 (10): e0163091. Bibcode:2016PLoSO..1163091M. Дои:10.1371 / journal.pone.0163091. ISSN 1932-6203. ЧВК 5061329. PMID 27732600.
- ^ Программа ООН по окружающей среде (2020). Совершенно неожиданно: ценность морской травы для окружающей среды и людей. ЮНЕП, Найроби. https://www.unenvironment.org/resources/report/out-blue-value-seagrasses-environment-and-people
- ^ а б Джонс, Клайв Дж .; Лоутон, Джон Х .; Шахак, Моше (1994). «Организмы как экосистемные инженеры». Ойкос. 69 (3): 373–386. Дои:10.2307/3545850. JSTOR 3545850.
- ^ Грей, Уильям; Моффлер, Марк (1987). «Цветение морской травы Thalassia testudinum (Hydrocharitacea) в заливе Тампа, Флорида». Водная ботаника. 5: 251–259. Дои:10.1016/0304-3770(78)90068-2.
- ^ Дарнелл, Келли; Дантон, Кеннет (2016). «Репродуктивная фенология субтропических морских водорослей Thalassia testudinum (черепаховая трава) и Halodule wrightii (мелководная трава) на северо-западе Мексиканского залива». Ботаника Марина. 59 (6): 473–483. Дои:10.1515 / бот-2016-0080. S2CID 88685282.
- ^ Macreadie, P. I .; Baird, M.E .; Trevathan-Tackett, S.M .; Larkum, A. W. D .; Ральф, П. Дж. (2013). «Количественная оценка и моделирование способности водорослей улавливать углерод». Бюллетень загрязнения морской среды. 83 (2): 430–439. Дои:10.1016 / j.marpolbul.2013.07.038. PMID 23948090.
- ^ https://myfwc.com/research/habitat/seagrasses/information/faq/#:~:text=Expand%2FCollapse%20What%20animals%20eat,on%20and%20among%20seagrass%20blades.
- ^ Nagelkerken, I .; Roberts, C.M .; van der Velde, G .; Dorenbosch, M .; van Riel, M.C .; Cocheret de la Morinière, E .; Ниенхейс, П. Х. (2002). «Насколько важны мангровые заросли и заросли водорослей для рыб коралловых рифов? Гипотеза питомников проверена в масштабе острова». Серия "Прогресс морской экологии". 244: 299–305. Bibcode:2002MEPS..244..299N. Дои:10,3354 / meps244299.
- ^ Nordlund, L.M .; Ансуорт, Р. К. Ф .; Gullstrom, M .; Каллен-Ансуорт, Л. С. (2018). «Глобальное значение промысла морских водорослей». Рыба и рыболовство. 19 (3): 399–412. Дои:10.1111 / faf.12259.
- ^ Ансуорт, Р. К. Ф .; Nordlund, L.M .; Каллен-Ансуорт, Л. С. (2019). «Луга водорослей поддерживают мировое рыболовство». Conserv Lett. e12566: e12566. Дои:10.1111 / conl.12566.
- ^ а б Boucek, R.E .; Leone, E .; Bickford, J .; Walters-Burnsed, S .; Лоуэрре-Барбьери, С. (2017). «Больше, чем просто место нереста: изучение мелкомасштабного использования двух устьевых видов рыб в месте скопления нереста». Границы морских наук (4): 1–9.
- ^ Байингтон, Кара (17 февраля 2017 г.). «Новая наука показывает, что луга водорослей подавляют патогены». Nature.org. Стипендиаты NatureNet за классную зеленую науку. Получено 17 февраля 2017.
- ^ Джонс, Би Джей; Каллен-Ансуорт, L.C .; Ансуорт, Р. К. Ф. (2018). «Отслеживание источника азота с использованием δ15N выявляет человеческие и сельскохозяйственные факторы, вызывающие деградацию водорослей на Британских островах». Границы науки о растениях. 9: 133. Дои:10.3389 / fpls.2018.00133. ЧВК 5808166. PMID 29467789.
- ^ МакГлэри, KJ (2001). «Цветение макроводорослей способствует исчезновению водорослей в прибрежных водах, обогащенных питательными веществами» (PDF). Журнал психологии. 37 (4): 453–456. Дои:10.1046 / j.1529-8817.2001.037004453.x. S2CID 38983997.
- ^ Fox SE, YS Olsen и AC Spivak (2010) «Воздействие восходящего и нисходящего контроля и изменения климата на сообщества эстуарных макрофитов и экосистемные услуги, которые они предоставляют» В: PF Kemp (Ed) Материалы симпозиума Eco-DAS, ТАКЖЕ, Глава 8: 129–145.
- ^ Ансуорт, Ричард К.Ф .; Кольер, Кэтрин Дж .; Уэйкотт, Мишель; Маккензи, Лен Дж .; Каллен-Ансуорт, Линн С. (2015). «Основа устойчивости экосистем водорослей». Бюллетень загрязнения морской среды. 100 (1): 34–46. Дои:10.1016 / j.marpolbul.2015.08.016. PMID 26342389.
- ^ "Реставрация Eelgrass | Охрана природы в Вирджинии". www.nature.org. Получено 2018-08-06.
- ^ «Реставрация водорослей». myfwc.com. Получено 2018-08-06.
- ^ «Инициатива по восстановлению водорослей - Малама Мауналуа». www.malamamaunalua.org. Получено 2018-08-06.
- ^ Ансуорт, Ричард К. Ф .; Маккензи, Лен Дж .; Кольер, Кэтрин Дж .; Каллен-Ансуорт, Линн С.; Дуарте, Карлос М .; Eklöf, Johan S .; Джарвис, Джесси С.; Джонс, Бенджамин Л .; Нордлунд, Лина М. (01.08.2019). «Глобальные проблемы сохранения водорослей». Ambio. 48 (8): 801–815. Дои:10.1007 / s13280-018-1115-y. ISSN 1654-7209. ЧВК 6541581. PMID 30456457.
- ^ van Katwijk, Marieke M .; Торхауг, Анитра; Марба, Нурия; Орт, Роберт Дж .; Дуарте, Карлос М .; Кендрик, Гэри А .; Althuizen, Inge H.J .; Балестри, Елена; Бернар, Гийом (25 ноября 2015 г.). «Глобальный анализ восстановления водорослей: важность крупномасштабных посадок». Журнал прикладной экологии. 53 (2): 567–578. Дои:10.1111/1365-2664.12562. ISSN 0021-8901.
- ^ Маккей, Джеки; Штюнцнер, Инга (24 октября 2019 г.). «Питомник морских водорослей в центральном Квинсленде может компенсировать выбросы углерода». ABC News. Австралийская радиовещательная корпорация. Получено 24 октября 2019.
Один цветок может дать 15 семян, а одно семя, посаженное в правильных условиях, может дать гектар морских водорослей.
Дальнейшие ссылки
- ден Хартог, К. 1970. Морские травы мира. Verhandl. der Koninklijke Nederlandse Akademie van Wetenschappen, Afd. Natuurkunde№ 59 (1).
- Дуарте, Карлос М. и Карина Л. Чискано «Биомасса и производство морских водорослей: переоценка» Aquatic Botany, том 65, выпуски 1–4, ноябрь 1999 г., страницы 159–174.
- Грин, Э. & Шорт, F.T. (ред.). 2003 г. Мировой атлас морских водорослей. Калифорнийский университет Press, Беркли, Калифорния. 298 стр.
- Хемминга, М.А. и Дуарте, К. 2000. Экология морских водорослей. Издательство Кембриджского университета, Кембридж. 298 стр.
- Хогарт, Питер Биология мангровых зарослей и морских водорослей (Издательство Оксфордского университета, 2007 г.)
- Ларкум, Энтони В.Д., Роберт Дж. Орт и Карлос М. Дуарте (редакторы) Морские травы: биология, экология и сохранение (Спрингер, 2006 г.)
- Орт, Роберт Дж. И др. «Глобальный кризис для экосистем водорослей» Бионаука Декабрь 2006 г. / Том. 56 № 12, страницы 987–996.
- Шорт, F.T. & Coles, R.G. (ред.). 2001 г. Глобальные методы исследования водорослей. Elsevier Science, Амстердам. 473 стр.
- A.W.D. Ларкум, Р.Дж. Орт и К. Дуарте (ред.). Биология водорослей: трактат. CRC Press, Бока-Ратон, Флорида, в печати.
- А. Шварц; М. Моррисон; И. Хоуз; Дж. Холлидей. 2006. Физические и биологические характеристики редкой морской среды обитания: приливных водорослей прибрежных островов. Наука в интересах сохранения 269. 39 стр. [1]
- Уэйкотт, М., МакМахон, К., и Лавери, П., 2014, Путеводитель по южным водорослям умеренного пояса, CSIRO Publishing, Мельбурн
внешняя ссылка
- Каллен-Ансуорт, Линн С.; Ансуорт, Ричард (2018-08-03). «Призыв к защите морских водорослей». Наука. 361 (6401): 446–448. Bibcode:2018Научный ... 361..446C. Дои:10.1126 / science.aat7318 (неактивно 30.11.2020). ISSN 0036-8075. PMID 30072524.CS1 maint: DOI неактивен по состоянию на ноябрь 2020 г. (связь)
- Project Seagrass - Благотворительная организация, содействующая сохранению морских водорослей посредством образования, влияния, исследований и действий.
- SeagrassSpotter - проект Citizen Science, направленный на повышение осведомленности о луговых водорослях и картографирование их местоположений.
- Кровать из водорослей и водорослей обзор со Смитсоновского океанского портала
- Статья в Nature Geoscience с описанием расположения лугов морских водорослей по всему миру.
- Seagrass-Watch - крупнейшая в мире научная программа неразрушающего контроля и мониторинга водорослей.
- Группа исследований экосистемы морских водорослей в Университете Суонси - междисциплинарные морские исследования в целях сохранения
- Restore-A-Scar - некоммерческая кампания по восстановлению лугов с водорослями, поврежденных лодочными подпорками.
- SeagrassNet - глобальная программа мониторинга водорослей
- Фонд морских водорослей в The Ocean Foundation
- Таксономия морских трав
- Всемирная ассоциация морских водорослей
- ВодорослиLI
- Наука и управление водорослями в Южно-Китайском море и Сиамском заливе
- Морская экология (Декабрь 2006 г.) - специальный выпуск о морских травах
- Камбоджийские морские травы
- Seagrass Productivity - COST Action ES0906
- Рыболовство Западной Австралии - Информационный бюллетень по водорослям

