ДМСО редуктаза - DMSO reductase
| Диметилсульфоксид редуктаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Номер ЕС | 1.8.5.3 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
ДМСО редуктаза это молибден -содержащий фермент, катализирующий восстановление диметилсульфоксид (ДМСО) в диметилсульфид (DMS). Этот фермент служит конечным редуктаза в анаэробных условиях у некоторых бактерий, где ДМСО является концевым акцептором электронов. В ходе реакции атом кислорода в ДМСО переводится в молибден, а затем восстанавливается до воды.
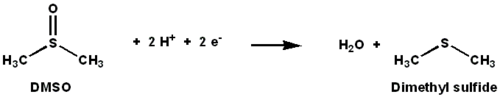
ДМСО-редуктаза (ДМСОР) и другие члены семейства ДМСО-редуктаз уникальны для бактерий и археи. Ферменты этого семейства в анаэробных окислительного фосфорилирования и на основе неорганических доноров литотрофный дыхание. Эти ферменты были разработаны для разложения оксоанионов.[1][2][3]DMSOR катализирует перенос двух электронов и одного атома кислорода в реакции: активный центр DMSOR содержит молибден, который в других случаях редко встречается в биологии.[2]
Третичная структура и активный сайт
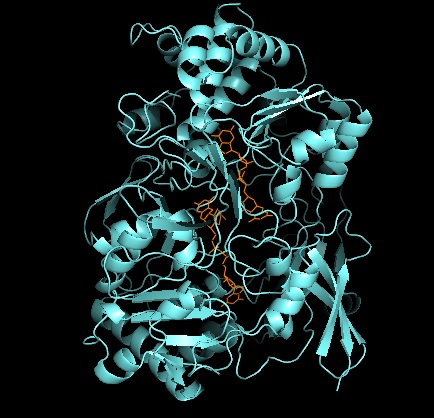
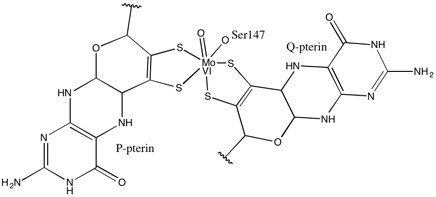
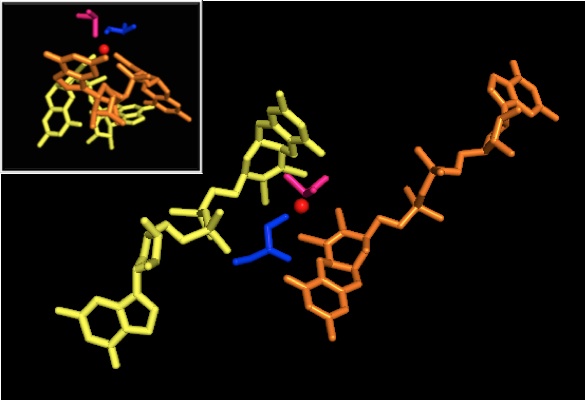
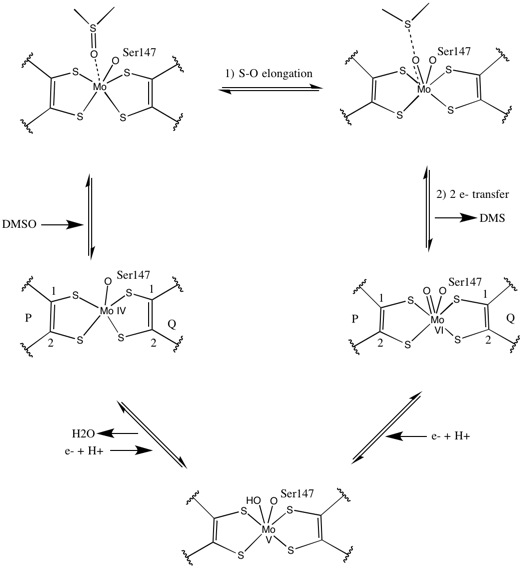
Что касается других членов семейства DMSO редуктаз, третичная структура DMSOR состоит из Mo-окружающих доменов I-IV, причем домен IV сильно взаимодействует с пираноптериндитиоленом. Мо-кофакторы (P- и Q-птерин) активного центра.[2][3] Члены семейства ДМСО-редуктазы различаются по активным сайтам.[3] В случае DMSOR центр Mo находится в двух дитиолен обеспечивается двумя кофакторами пираноптерина. Эти органические кофакторы, называемые молибдоптерины, связаны с GMP для создания динуклеотидной формы. Дополнительным пятым кэп-подобным лигандом является боковая цепь О остатка серина-147, дополнительно классифицируя фермент как ДМСО редуктазу III типа. Серин InType I и II заменен остатками цистеина и аспартата соответственно. В зависимости от окислительно-восстановительного состояния Мо, которое колеблется между IV, V или VI по мере развития реакции, ядро Мо активного центра также может быть лигировано с атомом кислорода аква-, гидроксо- или оксогруппы, соответственно. . Исследования показали, что конкретная идентичность аминокислоты, используемой для координации ядра Мо, сильно влияет на средний окислительно-восстановительный потенциал Мо и состояние протонирования лигирования кислородных групп, которые являются ключевыми детерминантами в механизме катализа фермента.[2]
Механизм
Исходный изотопный ДМСО18 исследования установили механизм двойной оксотрансферазы для DMSOR Р. sphaeroides. В этом механизме обозначенные O18 переносится с субстрата на Mo, который затем переносит O18 к 1,3,5-триаза-7-фосфаадамантан (PTA) для получения PTAO18.[6] По аналогичному механизму ДМСО переводит O в Mo, и образующийся центр Mo (VI) O восстанавливается, давая воду.[7]
Исследования синтетического Mo-бис-дитиолен комплексы предполагают перенос кислорода, перенос электронов. Используя XAS и DFT с S K-краем, эти модельные исследования указывают на согласованный разрыв S-O и перенос электронов. Скорость пропорциональна уменьшению прочности связи X-O субстрата и увеличению протонного сродства субстрата.[8]
Рентгеновская кристаллография Установлено, что общая третичная структура фермента остается постоянной в ходе реакции. Однако несколько различных экспериментов, проведенных на DMSOR Р. sphaeroides сообщили о различных результатах для координационной активности четырех потенциальных дитиоленовых лигандов. В то время как одно исследование рентгеновской кристаллографии пришло к выводу об эквидистантной координации всех четырех лигандов Mo-S в окисленной форме, что подтверждается многочисленными данными. рентгеновская абсорбционная спектроскопия (XAS), другое исследование характеризовало асимметричные расстояния Mo-S. Оба исследования, а также электронный парамагнитный резонанс (EPR) исследования предсказали, что активный сайт Мо очень гибкий с точки зрения положения и степени потенциальной координации лигандов.[7][9]
Данные, которые предполагают наличие двух существенно асимметричных кофакторов пираноптерина, были использованы для предположения механизма реакции. В полностью окисленной форме активного центра Mo VI оксогруппа и сериновые лиганды координировались на расстоянии 1,7 A от центра Mo. S1 и S2 P-птерина и S1 Q-птерина были расположены на расстоянии 2,4 А от Мо, а S2 Q-птерина - на расстоянии 3,1 А. Эта асимметрия птерина может быть результатом транс-эффекта оксогруппы, ослабляющего связь S2-Mo, которая расположена прямо напротив оксогруппы.[7]
Напротив, структура полностью восстановленной формы активного центра Mo IV показала, что S1 и S2 P-птерин и S1 Q-птерин сохраняли полную координацию, однако S2 Q-птерина сдвигался от металлического центра, что указывает на снижение координации. . Этот сдвиг длины связи лиганд-Мо согласуется с предполагаемым механизмом прямого переноса кислорода от субстрата ДМСО к Мо. Более слабая координация дитиолен в восстановленной форме фермента может способствовать прямому связыванию S = O. При восстановлении Мо и протонировании оксогруппы предполагается, что источник электронов цитохрома может связываться с углублением над активным центром и напрямую восстанавливать центр Мо, или же этот цитохром может связываться с хорошо сольватированной полипептидной петлей. в непосредственной близости от Q-птерина, и Q-птерин может опосредовать этот перенос электрона.[7]
Сотовая связь и регулирование
В Р. sphaeroides, DMSOR представляет собой водорастворимый белок с одной субъединицей, который не требует дополнительных кофакторов, кроме птерина. В Кишечная палочка, DMSOR встроен в мембрану и имеет три уникальные субъединицы, одна из которых включает характерный кофактор птерина, а другая - четыре 4Fe: кластеры 4S и последняя трансмембранная субъединица, которая связывает и окисляет менахинол. Перенос е- от менахинола к кластерам 4Fe: 4S и, наконец, к активному центру птерина-Мо генерирует протонный градиент, используемый для генерации АТФ.[7]
DMSOR регулируется преимущественно на уровне транскрипции. Он кодируется геном dor и экспрессируется при активации сигнальным каскадом, который регулируется белками DorS, DorR и DorC. Исследование lacZ слияния (репортерные гены) к соответствующим промоторам dorS, dorR и dorC пришли к выводу, что экспрессия DorR и DorC увеличивается в условиях пониженного содержания кислорода, но на экспрессию DorS не влияет концентрация кислорода. Экспрессия DorC также увеличивалась с увеличением концентрации ДМСО.[10]
Воздействие на окружающую среду
DMS, продукт DMSOR, является компонентом цикл серы. ДМС окисляется до Метансульфонаты, зарождающие конденсацию облаков над открытыми океанами, где альтернативный источник зародышеобразования - пыль - отсутствует. Образование облаков - ключевой компонент в увеличении альбедо Земли и регулировании температуры атмосферы, поэтому этот фермент и реакция, которую он катализирует, могут оказаться полезными на границе контроля климата.[11]
Рекомендации
- ^ Капплер У., Шефер Х (2014). "Глава 11. Превращения диметилсульфида.". В Kroneck PM, Torres ME (ред.). Металлическая биогеохимия газообразных соединений окружающей среды. Ионы металлов в науках о жизни. 14. Springer. С. 279–313. Дои:10.1007/978-94-017-9269-1_11. ISBN 978-94-017-9268-4. PMID 25416398.
- ^ а б c d МакЭван А.Г., Капплер У. (2004). "Семейство ДМСО-редуктазы микробных ферментов молибдена" (PDF). Австралийский биохимик. 35 (3): 17–20. Архивировано из оригинал (PDF) на 2014-03-07. Получено 2014-02-27.
- ^ а б c d е McEwan AG, Ridge JP, McDevitt CA, Hugenholtz P (2002). «Семейство ДМСО редуктазы микробных ферментов молибдена; молекулярные свойства и роль в диссимиляционном восстановлении токсичных элементов». Журнал геомикробиологии. 19 (1): 3–21. Дои:10.1080/014904502317246138.
- ^ PDB: 1DMS; Шнайдер Ф., Лёве Дж., Хубер Р., Шинделин Х., Кискер С., Кнеблейн Дж. (Октябрь 1996 г.). «Кристаллическая структура диметилсульфоксидредуктазы из Rhodobacter capsulatus при разрешении 1,88 A». Журнал молекулярной биологии. 263 (1): 53–69. Дои:10.1006 / jmbi.1996.0555. PMID 8890912.
- ^ PDB: 4ДМР; McAlpine AS, McEwan AG, Bailey S (январь 1998 г.). «Кристаллическая структура высокого разрешения ДМСО редуктазы в комплексе с ДМСО». Журнал молекулярной биологии. 275 (4): 613–23. Дои:10.1006 / jmbi.1997.1513. PMID 9466935.
- ^ Schultz BE, Hille R, Holm RH (1995), "Прямой перенос атома кислорода в механизме действия диметилсульфоксидредуктазы Rhodobacter sphaeroides", Журнал Американского химического общества, 117 (2): 827–828, Дои:10.1021 / ja00107a031, ISSN 0002-7863
- ^ а б c d е Кискер С, Шинделин Х, Рис, округ Колумбия (1997). «Ферменты, содержащие кофактор молибдена: структура и механизм» (PDF). Ежегодный обзор биохимии. 66: 233–67. Дои:10.1146 / annurev.biochem.66.1.233. PMID 9242907.
- ^ Тендерхольт А.Л., Ван Дж.Дж., Силагьи Р.К., Холм Р.Х., Ходжсон К.О., Хедман Б., Соломон Е.И. (июнь 2010 г.). «Рентгеновская абсорбционная спектроскопия с K-краем серы и расчеты функционала плотности для Mo (IV) и Mo (VI) = O бис-дитиоленов: понимание механизма переноса оксо в ДМСО редуктазе и родственных функциональных аналогах». Журнал Американского химического общества. 132 (24): 8359–71. Дои:10.1021 / ja910369c. ЧВК 2907113. PMID 20499905.
- ^ Макалпайн А.С., МакЭван А.Г., Шоу А.Л., Бейли С. (1997). «Молибденовый активный центр ДМСО редуктазы из Rhodobacter capsulatus: кристаллическая структура окисленного фермента с разрешением 1,82 А и фермента, восстановленного дитионитом, с разрешением 2,8 А». JBIC. 2 (6): 690–701. Дои:10.1007 / s007750050185.
- ^ Gunsalus RP (ноябрь 1992 г.). «Контроль потока электронов в Escherichia coli: скоординированная транскрипция генов дыхательного пути». Журнал бактериологии. 174 (22): 7069–74. ЧВК 207394. PMID 1331024.
- ^ Саркар Б. (21 марта 2002 г.). Тяжелые металлы в окружающей среде. CRC Press. п. 456. ISBN 978-0-8247-4475-5.