Гаплогруппа L0 (мтДНК) - Haplogroup L0 (mtDNA)
| Гаплогруппа L0 | |
|---|---|
| Возможное время происхождения | От 130 до 200 тыс. Лет назад[1][2] |
| Возможное место происхождения | Южная Африка или же Юго-Восточная Африка |
| Предок | L (Митохондриальная Ева ) |
| Потомки | L0a'b'f'k, L0d |
| Определение мутаций | 263 !, 1048, 3516A, 5442, 6185, 9042, 9347, 10589, 12007, 12720[3] |
Гаплогруппа L0 это митохондриальная ДНК человека (мтДНК) гаплогруппа.
Источник

L0 - одна из двух ветвей от самый последний общий предок (MRCA) для общей человеческой материнской линии. Гаплогруппа состоит из пяти основных ветвей (L0a, L0b, L0d, L0f, L0k). Четыре из них изначально были классифицированы на субклады L1, L1a, L1d, L1f и L1k.
В 2014 году древний анализ ДНК скелета 2330-летнего самца-фуражира в г. Южная Африка установили, что образец принадлежал к субкладу мтДНК L0d2c1c. Эта материнская гаплогруппа сегодня наиболее тесно связана с дзю, подгруппой коренных народов. Народ сан, что свидетельствует о преемственности населения в регионе.[4] В 2016 г. Поздний железный век иссушенная мумия из региона Тули на севере Ботсваны также принадлежит к гаплогруппе L0.[5]
| MRCA (мтДНК) |
| ||||||||||||||||||||||||||||||||||||
Распределение

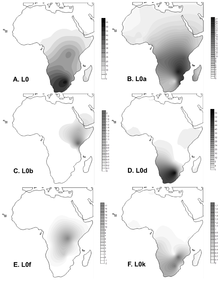
L0 чаще всего встречается в К югу от Сахары. Наивысшей частоты он достигает в Хойсан человек в среднем 73%.[6] Некоторые из самых высоких частот:[7] Намибия (! Сюнь ) 79%, Южная Африка (Кхве /! Сюнь) 83%, и Ботсвана (! Кунг ) 100%.
Гаплогруппа L0d встречается среди койсанских групп Южная Африка ближе к хоидской стороне с (после L0k) более санидской, но в значительной степени ограничивается койсанами в целом.[7][8][9][10] L0d также часто встречается в Цветные население ЮАР и частота колеблется от 60%[11] до 71%.[10] Это иллюстрирует огромный материнский вклад койсанского народа в развитие цветного населения Южной Африки.
Гаплогруппы L0k является второй по распространенности гаплогруппой в койсанских группах, более близких к санидской стороне (после L0d), являющейся более хоидской, но в значительной степени ограниченной койсанцами в целом.[7][8][9][10] Хотя связанная с Хойсаном гаплогруппа L0d была обнаружена с высокой частотой в цветном населении Южной Африки, L0k не наблюдались в двух исследованиях с участием больших групп цветных людей.[10][11]
Гаплогруппа L0f присутствует на относительно небольших частотах в Танзания, Восточная Африка среди Люди сандаве Танзании, которые старше хойсанов.
Гаплогруппа L0a наиболее распространена среди населения Юго-Восточной Африки (25% в Мозамбик ).[6]Среди Гвинейцы, он имеет частоту от 1% до 5%, с Баланта группа показывает повышенную частоту около 11%. Гаплогруппа L0a имеет палеолитическую временную глубину около 33 000 лет и, вероятно, достигла Гвинеи между 10 000 и 4 000 лет назад. Это также часто встречается в Мбути и Биака Пигмеи. L0a встречается с частотой почти 25% в Hadramawt (Йемен ).[12]
Гаплогруппа L0b Эфиопия.
Взаимодействие с лекарствами и болезнями
У пациентов, которым вводят препарат ставудин лечить ВИЧ, Гаплогруппа L0a2 связана с более высокой вероятностью периферическая невропатия как побочный эффект.[13]
Субклады
Дерево

Это филогенетическое дерево субкладов гаплогруппы L0 основано на статье Манниса ван Овена и Манфреда Кайзера. Обновленное комплексное филогенетическое дерево глобальных вариаций митохондриальной ДНК человека[3] и последующие опубликованные исследования.
- Самый последний общий предок (MRCA)
- L0
- L0d
- L0d3
- L0d1'2
- L0d1
- L0d1a
- L0d1b
- L0d1c
- L0d1c1
- L0d2
- L0d2a'b
- L0d2a
- L0d2a1
- L0d2b
- L0d2a
- L0d2c
- L0d2c1c[14]
- L0d2a'b
- L0d1
- L0a'b'f'k
- L0k
- L0k1
- L0k2
- L0a'b'f
- L0f
- L0f1
- L0f2
- L0f2a
- L0f2b
- L0a'b
- L0a
- L0a1
- L0a1a
- L0a1a2
- L0a1b
- L0a1b1
- L0a1b1a
- L0a1b2
- L0a1b1
- L0a1c
- L0a1d
- L0a1a
- L0a2
- L0a2a
- L0a2a1
- L0a2a1a
- L0a2a1a1
- L0a2a1a2
- L0a2a1a
- L0a2a2
- L0a2a2a
- L0a2a1
- L0a2b
- L0a2ba
- L0a2c
- L0a2d
- L0a2a
- L0a3
- L0a4
- L0a1
- L0b
- L0a
- L0f
- L0k
- L0d
- L0
Смотрите также
- Генеалогический ДНК-тест
- Генетическая генеалогия
- Митохондриальная генетика человека
- Популяционная генетика
- Гаплогруппы митохондриальной ДНК человека
Филогенетическое дерево гаплогруппы митохондриальной ДНК человека (мтДНК) | |||||||||||||||||||||||||||||||||||||||
| Митохондриальная Ева (L ) | |||||||||||||||||||||||||||||||||||||||
| L0 | L1–6 | ||||||||||||||||||||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | L6 | ||||||||||||||||||||||||||||||||||
| M | N | ||||||||||||||||||||||||||||||||||||||
| CZ | D | E | грамм | Q | О | А | S | р | я | W | Икс | Y | |||||||||||||||||||||||||||
| C | Z | B | F | R0 | pre-JT | п | U | ||||||||||||||||||||||||||||||||
| HV | JT | K | |||||||||||||||||||||||||||||||||||||
| ЧАС | V | J | Т | ||||||||||||||||||||||||||||||||||||
Рекомендации
- ^ точечная оценка 168,5 тыс. лет (136,3–201,1 тыс. лет) 95% ДИ ) в соответствии с Хайнц, Таня; и другие. (2017). «Обновление африканского дерева митохондриальной ДНК человека: актуальность для судебной медицины и популяционной генетики». Forensic Science International: генетика. 27: 156–159. Дои:10.1016 / j.fsigen.2016.12.016. PMID 28086175. (таблица 2) .150 тыс. лет предлагается в:Соарес, Педро; Эрмини, Лука; Томсон, Ноэль; Мормина, Мару; Рито, Тереза; Рёль, Арне; Салас, Антонио; Оппенгеймер, Стивен; Маколей, Винсент (2009). «Поправка на очищающий отбор: улучшенные митохондриальные молекулярные часы человека». Американский журнал генетики человека. 84 (6): 740–59. Дои:10.1016 / j.ajhg.2009.05.001. ЧВК 2694979. PMID 19500773..
- ^ Оценки возраста (ka, 95% ДИ в угловых скобках): оценка возраста всей мтДНК ML: 128,2 [95% ДИ: 107,9–148,9], оценка возраста цельной мтДНК ρ: 121,3 [99,2; 143,7], оценка возраста синонима ρ ( ka): 131,0 [97,8; 164,2]: Рито Т., Ричардс МБ, Фернандес В., Альшамали Ф., Черни В., Перейра Л., Соареш П., «Первые современные расселения людей по Африке», PLoS One 2013 Nov 13; 8 (11): e80031. DOI: 10.1371 / journal.pone.0080031.
- ^ а б Ван Овен, Маннис; Кайзер, Манфред (2009). «Обновленное комплексное филогенетическое дерево глобальных вариаций митохондриальной ДНК человека». Человеческая мутация. 30 (2): E386–94. Дои:10.1002 / humu.20921. PMID 18853457. S2CID 27566749.
- ^ Алан Дж. Моррис; Аня Хайнце; Ева К.Ф. Чан; Эндрю Б. Смит; Ванесса М. Хейс (2014). «Первый древний митохондриальный геном человека из южноафриканских докотоводов». Геномная биология и эволюция. 6 (10): 2647–53. Дои:10.1093 / gbe / evu202. ЧВК 4224329. PMID 25212860. Получено 17 апреля 2016.
- ^ Франк Дж. Рюли; Марина Стейн; Моронгва Н. Мосотване; Лена Эрстрём; Молебогенг К. Бодиба; Эбигейл Бауман (январь – февраль 2016 г.). «Радиологический и генетический анализ мумии позднего железного века из блока Тули, Ботсвана» (PDF). Южноафриканский научный журнал. 112 (1/2). Архивировано из оригинал (PDF) 21 июня 2016 г.. Получено 26 апреля 2016.
- ^ а б Роза, Александра; Брем, Антонио; Кивисилд, Тоомас; Мецпалу, Эне; Виллемс, Ричард (2004). «Профиль мтДНК жителей Западной Африки в Гвинее: на пути к лучшему пониманию региона Сенегамбия». Анналы генетики человека. 68 (4): 340–52. Дои:10.1046 / j.1529-8817.2004.00100.x. PMID 15225159. S2CID 15391342.
- ^ а б c Тишкофф, С. А .; Gonder, M. K .; Henn, B.M .; Mortensen, H .; Knight, A .; Gignoux, C .; Fernandopulle, N .; Lema, G .; Ньямбо, Т. Б. (2007). "История кликабельных популяций Африки на основе генетической изменчивости мтДНК и Y-хромосомы". Молекулярная биология и эволюция. 24 (10): 2180–95. Дои:10.1093 / молбев / мсм155. PMID 17656633.
- ^ а б Чен Юй-Шэн; Олькерс, Антонель; Schurr, Theodore G .; Когельник, Андреас М .; Хуопонен, Кирси; Уоллес, Дуглас С. (2000). «Вариации мтДНК в южноафриканских кунгах и кхве - и их генетические связи с другими африканскими популяциями». Американский журнал генетики человека. 66 (4): 1362–83. Дои:10.1086/302848. ЧВК 1288201. PMID 10739760.
- ^ а б Рыцарь, Алек; Андерхилл, Питер А .; Мортенсен, Холли М .; Животовский, Лев А .; Лин, Алиса А .; Henn, Brenna M .; Луи, Дороти; Рулен, Мерритт; Гора, Джоанна Л. (2003). «Африканская Y-хромосома и дивергенция мтДНК обеспечивает понимание истории языков клика». Текущая биология. 13 (6): 464–73. Дои:10.1016 / S0960-9822 (03) 00130-1. PMID 12646128. S2CID 52862939.
- ^ а б c d Schlebusch, Carina M .; Найду, Тиессен; Соодьял, Химла (2009). «Минисеквенирование SNaPshot для разрешения митохондриальных макрогаплогрупп, обнаруженных в Африке». Электрофорез. 30 (21): 3657–64. Дои:10.1002 / elps.200900197. PMID 19810027. S2CID 19515426.
- ^ а б Кинтана-Мурси, Луис; Хармант, Кристина; Квач, Элен; Балановский, Олег; Запорожченко, Валерий; Борман, Конни; Ван Хелден, Пол Д .; Hoal, Eileen G .; Бехар, Дорон М. (2010). "Сильный вклад материнской койсанской семьи в развитие цветного населения Южной Африки: случай гендерно-предвзятого смешения". Американский журнал генетики человека. 86 (4): 611–20. Дои:10.1016 / j.ajhg.2010.02.014. ЧВК 2850426. PMID 20346436.
- ^ Ридл, Якуб; Эденс, Кристофер М .; Черный, Виктор (2009). «Структура митохондриальной ДНК йеменского населения: региональные различия и последствия для различных миграционных факторов». Эволюция человеческих популяций в Аравии. Палеобиология и палеоантропология позвоночных. С. 69–78. Дои:10.1007/978-90-481-2719-1_5. ISBN 978-90-481-2718-4.
- ^ Кампира Э., Кумвенда Дж., Ван Остерхаут Дж. Дж., Дандара К. Субгаплогруппы L0a2 и L2a митохондриальной ДНК изменяют предрасположенность к периферической невропатии у взрослых малавийских жителей, получающих ставудин, содержащий высокоактивную антиретровирусную терапию., J Acquir Immune Defic Syndr. 2013 Aug 15; 63 (5): 647-52. DOI: 10.1097 / QAI.0b013e3182968ea5
- ^ Первый древний митохондриальный геном человека от доскотоводства из Южной Африки
внешняя ссылка
- Общий
- Яна Логана Сайт митохондриальной ДНК
- Манниса ван Овена Филотрия
- Гаплогруппа L0
- Роза, Александра; Брем, Антонио; Кивисилд, Тоомас; Мецпалу, Эне; Виллемс, Ричард (2004). «Профиль мтДНК жителей Западной Африки в Гвинее: на пути к лучшему пониманию региона Сенегамбия». Анналы генетики человека. 68 (4): 340–52. Дои:10.1046 / j.1529-8817.2004.00100.x. PMID 15225159. S2CID 15391342.
Основываясь на предыдущих знаниях африканских полных последовательностей, парафилетическая клада L1 делится на две монофилетические единицы L0, захватывающие ранее определенные клоны L1a и L1d, и кладу L1, которая включает клады L1b и L1c…
- Павеси, А. (2005). «Полезность полиомавируса JC в отслеживании модели миграции людей, относящейся к доисторическим временам». Журнал общей вирусологии. 86 (5): 1315–26. Дои:10.1099 / vir.0.80650-0. PMID 15831942.
С другой стороны, первая ось мтДНК помещает предковую гаплогруппу L0 в крайнее левое положение, так как она дала начало одной единственной линии ...
- Мишмар, Дан; Руис-Песини, Эдуардо; Брэндон, Мартин; Уоллес, Дуглас С. (2004). «Митохондриальные ДНК-подобные последовательности в ядре (NUMT): понимание нашего африканского происхождения и механизма интеграции чужеродной ДНК». Человеческая мутация. 23 (2): 125–33. Дои:10.1002 / humu.10304. PMID 14722916. S2CID 25109836.
мтДНК гаплогруппы L0, гаплогруппа, которую мы ранее пришли к выводу, лежит в основе мтДНК человека. дерево на основе филогенетического анализа…
- Рыцарь, Алек; Андерхилл, Питер А .; Мортенсен, Холли М .; Животовский, Лев А .; Лин, Алиса А .; Henn, Brenna M .; Луи, Дороти; Рулен, Мерритт; Гора, Джоанна Л. (2003). «Африканская Y-хромосома и дивергенция мтДНК обеспечивает понимание истории языков клика». Текущая биология. 13 (6): 464–73. Дои:10.1016 / S0960-9822 (03) 00130-1. PMID 12646128. S2CID 52862939.
- Роза, Александра; Брем, Антонио; Кивисилд, Тоомас; Мецпалу, Эне; Виллемс, Ричард (2004). «Профиль мтДНК жителей Западной Африки в Гвинее: на пути к лучшему пониманию региона Сенегамбия». Анналы генетики человека. 68 (4): 340–52. Дои:10.1046 / j.1529-8817.2004.00100.x. PMID 15225159. S2CID 15391342.