Белки Notch - Википедия - Notch proteins
| Notch (LNR) домен | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Идентификаторы | |||||||||
| Символ | Notch | ||||||||
| Pfam | PF00066 | ||||||||
| ИнтерПро | IPR000800 | ||||||||
| УМНАЯ | SM00004 | ||||||||
| PROSITE | PS50258 | ||||||||
| OPM суперсемейство | 462 | ||||||||
| Белок OPM | 5кзо | ||||||||
| Мембранома | 19 | ||||||||
| |||||||||
Notch белки площадь семья типа-1 трансмембранные белки которые составляют ключевой компонент Notch сигнальный путь, который хорошо сохраняется в многоклеточные животные. Нотч внеклеточный домен (NECD) опосредует взаимодействие с семейством DSL лиганды, позволяя ему участвовать в юкстакриновая сигнализация Внутриклеточный домен Notch (NICD) действует как активатор транскрипции в комплексе с семьей CSL факторы транскрипции. Члены этого семейства трансмембранных белков типа 1 имеют несколько общих структур ядра, включая внеклеточный домен, состоящий из нескольких фактор роста эпидермиса (EGF) -подобные повторы и домен активации транскрипции внутриклеточного домена (TAD). Члены семейства Notch действуют в различных тканях и играют роль во множестве процессов развития, контролируя судьба клетки решения. Многое из того, что известно о функции Notch, получено из исследований, проведенных в Caenorhabditis elegans (C.elegans) и Drosophila melanogaster. Человеческие гомологи также были идентифицированы, но детали функции Notch и взаимодействия с его лигандами в этом контексте не известны.
Открытие
Notch был обнаружен в мутант Дрозофила в марте 1913 г. в лаборатории Томас Хант Морган.[2] Этот мутант появился после нескольких поколений скрещивания крылатых мух и мух дикого типа и был впервые охарактеризован Джоном С. Декстером.[3] Наиболее часто наблюдаемый фенотип у мутантных мух Notch - это появление вогнутой зазубрины на самом дальнем конце крыльев, у которой ген назван, сопровождается отсутствием краевых щетинок.[4][5] Этот мутант оказался связанный с полом доминирует на Х хромосома это можно было наблюдать только у гетерозиготных женщин, поскольку это было летальным для мужчин и гомозиготных женщин.[2] Первый Notch аллель была основана в 1917 году К. В. Метцем и К. Б. Бриджесом.[6] В конце 1930-х гг. Исследования мух эмбриогенез выполненный Дональдом Ф. Поулсоном, дал первое указание на роль Нотча в развитии.[7] У мутантных самцов Notch-8 отсутствовала внутренняя ростковые отростки, то энтодерма и мезодерма, что привело к невозможности последующего морфогенез эмбриональная летальность. Позднее учеба в начале Дрозофила нейрогенез предоставили некоторые из первых указаний на роль Нотча в передаче сигналов клетка-клетка, поскольку нервная система in Notch мутанты были разработаны путем принесения в жертву гиподермальных клеток.[8]
Начиная с 1980-х годов исследователи начали глубже понимать функцию Notch с помощью генетических и молекулярных экспериментов. Генетический скрининг, проведенный в Дрозофила привело к идентификации нескольких белков, которые играют центральную роль в передаче сигналов Notch, включая Enhancer of split,[8] Мастерский разум, Дельта,[9] Подавитель безволосого (CSL),[10] и зазубрины.[11] В то же время ген Notch был успешно секвенирован.[12][13] и клонирован,[14][15] понимание молекулярной архитектуры белков Notch и привело к идентификации Notch гомологи в Caenorhabditis elegans (C. elegans)[16][17][18] и в конечном итоге в млекопитающие.
В начале 1990-х годов Notch все чаще использовался как рецептор ранее неизвестного межклеточного сигнального пути.[19][20] в котором NICD транспортируется в ядро где он действует как фактор транскрипции для прямой регуляции генов-мишеней.[21][22][23] Было обнаружено, что высвобождение NICD происходит в результате протеолитического расщепления трансмембранного белка под действием γ-секретаза комплексная каталитическая субъединица Пресенилин. Это было значимое взаимодействие, поскольку пресенилин участвует в развитии болезни Альцгеймера.[24] Это и дальнейшие исследования механизма подписи Notch привели к исследованию, которое еще больше связывает Notch с широким спектром заболеваний человека.
Структура
Дрозофила содержат единственный белок Notch, C. elegans содержат два дублирующих паралога с надрезом, Лин-12[25] и GLP-1,[18][26] а у людей есть четыре варианта Notch, Notch 1-4. Хотя между гомологами существуют вариации, существует набор высококонсервативных структур, обнаруженных во всех белках семейства Notch. Белок может быть разделен на внеклеточный домен Notch (NECD) и внутриклеточный домен Notch (NICD), соединенные вместе посредством однопроходного трансмембранного домена (TM).
NECD содержит 36 повторов EGF в Дрозофила,[13] 28-36 у человека и 13 и 10 у C. elegans Лин-12 и ГПП-1 соответственно.[27] Эти повторы сильно изменены О-гликозилирование[28] и было показано, что добавление определенных O-связанных гликанов необходимо для правильного функционирования. За повторами EGF следуют три цистеин -богатые Lin-12 / Notch Repeats (LNR) и домен гетеродимеризации (HD). Вместе LNR и HD составляют отрицательную регуляторную область, прилегающую к клеточная мембрана и помогают предотвратить передачу сигналов в отсутствие связывания лиганда.
NICD действует как фактор транскрипции, который высвобождается после того, как связывание лиганда запускает его расщепление. Он содержит последовательность ядерной локализации (NLS), которая опосредует его транслокацию в ядро. где он образует транскрипционный комплекс вместе с несколькими другими факторами транскрипции. Попав в ядро, несколько акириновых повторов (ANK) и RAM-домен взаимодействуют между белками NICD и CSL с образованием комплекса активации транскрипции.[29] У людей дополнительный домен PEST играет роль в деградации NICD.[30]
Функция
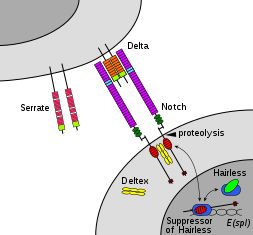
Члены семейства Notch играют роль во множестве процессов развития, контролируя решения о судьбе клеток. Сеть сигнализации Notch - это эволюционно сохраненный межклеточный сигнальный путь который регулирует взаимодействие между физически соседними клетками. В Дрозофила, notch взаимодействие с его клеточно-связанными лигандами (дельта, пильчатый) устанавливает межклеточный сигнальный путь, который играет ключевую роль в развитии. Этот белок функционирует как рецептор для мембраносвязанных лигандов и может играть несколько ролей во время развития.[31]Дефицит может быть связан с двустворчатый аортальный клапан.[32]
Есть данные, что активировали Notch 1 и Вырез 3 способствовать дифференцировке клеток-предшественников в астроглия.[33] Notch 1, активированный до рождения, вызывает радиальная глия дифференциация,[34] но постнатально вызывает дифференциацию в астроциты.[35] Одно исследование показывает, что каскад Notch-1 активируется Reelin неустановленным способом.[36] Reelin и Notch1 сотрудничают в разработке зубчатые извилины, согласно другому.[37]
Лиганд взаимодействия
| Зубчатый / зубчатый белок | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | DSL | ||||||||
| Pfam | PF01414 | ||||||||
| ИнтерПро | IPR026219 | ||||||||
| Мембранома | 76 | ||||||||
| |||||||||
Передача сигналов Notch запускается посредством прямого межклеточного контакта, опосредованного взаимодействиями между белком рецептора Notch в принимающей сигнал клетке и лигандом в соседней передающей сигнал клетке. Эти однопроходные трансмембранные белки типа 1 относятся к семейству белков Delta / Serrate / Lag-2 (DSL), названному в честь трех канонических лигандов Notch.[19] Delta и Serrate встречаются в Дрозофила в то время как Лаг-2 находится в C. elegans. Люди содержат 3 дельта-гомолога, дельта-подобные 1, 3 и 4, а также два зубчатых гомолога, Зубчатый 1 и 2. Белки Notch состоят из относительно короткого внутриклеточного домена и большого внеклеточного домена с одним или несколькими EGF мотивы и N-концевой DSL-связывающий мотив. Было показано, что повторы EGF 11-12 во внеклеточном домене Notch необходимы и достаточны для транс сигнальные взаимодействия между Notch и его лигандами.[38] Кроме того, повторы EGF 24-29 участвуют в ингибировании СНГ взаимодействия между Notch и лигандами коэкспрессируются в одной и той же клетке.[39]
Протеолиз
Для того, чтобы произошло событие передачи сигнала, белок Notch должен расщепляться на нескольких сайтах. У людей Notch сначала расщепляется в домене NRR с помощью Furin, а процессируется в сеть транс-Гольджи до того, как он будет представлен на поверхности клетки в виде гетеродимера.[40][41] Дрозофила Notch не требует этого расщепления для передачи сигналов,[42] и есть некоторые свидетельства того, что LIN-12 и GLP-1 расщепляются по этому сайту в C. elegans.
Высвобождение NICD достигается после дополнительных двух событий расщепления Notch. Связывание Notch с лигандом DSL приводит к конформационному изменению, которое открывает сайт расщепления в NECD. Ферментативный протеолиз в этом сайте осуществляется протеазой семейства A-дезинтегрин и металлопротеазный домен (ADAM). Этот белок называется кузбанским в Дрозопихла,[43][44] sup-17 в C. elegans,[45] и ADAM10 у людей.[46][47] После протеолитического расщепления высвободившийся NECD эндоцитозируется в клетку, передающую сигнал, оставляя после себя только небольшую внеклеточную часть Notch. Этот усеченный белок Notch затем может распознаваться γ-секретазой, которая расщепляет третий сайт, обнаруженный в домене TM.[48]
Человеческие гомологи
Notch-1
Notch-2
Notch-2 (Нейрогенный белок-гомолог notch локуса 2) это белок что у людей кодируется NOTCH2 ген.[49]
NOTCH2 связан с Синдром Алажиля[50] и Синдром Хайду – Чейни.[51]
Notch-3
Notch-4
Смотрите также
Примечания
- ^ Вардар Д., North CL, Санчес-Ирисарри C, Астер JC, Blacklow SC (июнь 2003 г.). «Структура ядерного магнитного резонанса прототипа повторяющегося модуля Lin12-Notch из человеческого Notch1». Биохимия. 42 (23): 7061–7. Дои:10.1021 / bi034156y. PMID 12795601.
- ^ а б Морган TH, Мосты CB (1916). Наследование, сцепленное с полом у дрозофилы. Библиотеки NCSU. Вашингтон, Вашингтонский институт Карнеги.
- ^ Декстер Дж. С. (декабрь 1914 г.). "Анализ случая непрерывной изменчивости у дрозофилы путем изучения ее отношений сцепления". Американский натуралист. 48 (576): 712–758. Дои:10.1086/279446.
- ^ Мор О.Л. (май 1919 г.). «Изменения характера, вызванные мутацией всей области хромосомы у дрозофилы». Генетика. 4 (3): 275–82. ЧВК 1200460. PMID 17245926.
- ^ Линдсли Д.Л., Зимм Г.Г. (2012-12-02). Геном Drosophila Melanogaster. Академическая пресса. ISBN 9780323139847.
- ^ Metz CW, Bridges CB (декабрь 1917 г.). «Несовместимость мутантных рас у дрозофилы». Труды Национальной академии наук Соединенных Штатов Америки. 3 (12): 673–8. Bibcode:1917ПНАС .... 3..673М. Дои:10.1073 / pnas.3.12.673. ЧВК 1091355. PMID 16586764.
- ^ Поулсон Д.Ф. (март 1937 г.). «Хромосомная недостаточность и эмбриональное развитие Drosophila Melanogaster». Труды Национальной академии наук Соединенных Штатов Америки. 23 (3): 133–7. Bibcode:1937ПНАС ... 23..133П. Дои:10.1073 / пнас.23.3.133. ЧВК 1076884. PMID 16588136.
- ^ а б Леманн Р., Хименес Ф., Дитрих Ю., Кампос-Ортега Дж. А. (март 1983 г.). «О фенотипе и развитии мутантов раннего нейрогенеза у Drosophila melanogaster». Архив биологии развития Вильгельма Ру. 192 (2): 62–74. Дои:10.1007 / BF00848482. PMID 28305500. S2CID 25602190.
- ^ Леманн Р., Дитрих Ю., Хименес Ф., Кампос-Ортега Дж. А. (июль 1981 г.). «Мутации раннего нейрогенеза у дрозофилы». Архив биологии развития Вильгельма Ру. 190 (4): 226–229. Дои:10.1007 / BF00848307. PMID 28305572. S2CID 21814447.
- ^ Фортини М.Э., Артаванис-Цаконас С. (октябрь 1994 г.). «Супрессор бесшерстного белка участвует в передаче сигналов рецептора notch». Клетка. 79 (2): 273–82. Дои:10.1016/0092-8674(94)90196-1. PMID 7954795. S2CID 40771329.
- ^ Fleming RJ, Scottgale TN, Diederich RJ, Artavanis-Tsakonas S (декабрь 1990 г.). «Ген Serrate кодирует предполагаемый EGF-подобный трансмембранный белок, необходимый для правильного эктодермального развития у Drosophila melanogaster». Гены и развитие. 4 (12A): 2188–201. Дои:10.1101 / gad.4.12a.2188. PMID 2125287.
- ^ Кидд С., Келли М.Р., Янг М.В. (сентябрь 1986 г.). «Последовательность notch локуса Drosophila melanogaster: взаимосвязь кодируемого белка с факторами свертывания и роста млекопитающих». Молекулярная и клеточная биология. 6 (9): 3094–108. Дои:10.1128 / mcb.6.9.3094. ЧВК 367044. PMID 3097517.
- ^ а б Wharton KA, Johansen KM, Xu T., Artavanis-Tsakonas S (декабрь 1985 г.). «Нуклеотидная последовательность из вырезки нейрогенного локуса подразумевает продукт гена, который имеет гомологию с белками, содержащими EGF-подобные повторы». Клетка. 43 (3, часть 2): 567–81. Дои:10.1016/0092-8674(85)90229-6. PMID 3935325.
- ^ Кидд С., Локетт Т.Дж., Янг М.В. (сентябрь 1983 г.). «Notch locus Drosophila melanogaster». Клетка. 34 (2): 421–33. Дои:10.1016/0092-8674(83)90376-8. PMID 6193889. S2CID 36425372.
- ^ Артаванис-Цаконас С., Мускавич М.А., Едвобник Б. (апрель 1983 г.). «Молекулярное клонирование Notch, локуса, влияющего на нейрогенез у Drosophila melanogaster». Труды Национальной академии наук Соединенных Штатов Америки. 80 (7): 1977–81. Bibcode:1983ПНАС ... 80.1977А. Дои:10.1073 / пнас.80.7.1977. ЧВК 393735. PMID 6403942.
- ^ Гринвальд I (февраль 1987 г.). «Локус lin-12 Caenorhabditis elegans». BioEssays. 6 (2): 70–3. Дои:10.1002 / bies.950060207. PMID 3551950. S2CID 19253030.
- ^ Присс-младший, Шнабель Х, Шнабель Р. (ноябрь 1987 г.). «Локус glp-1 и клеточные взаимодействия в ранних эмбрионах C. elegans». Клетка. 51 (4): 601–11. Дои:10.1016/0092-8674(87)90129-2. PMID 3677169. S2CID 6282210.
- ^ а б Остин Дж., Кимбл Дж. (Ноябрь 1987 г.). «glp-1 необходим в зародышевой линии для регуляции выбора между митозом и мейозом у C. elegans». Клетка. 51 (4): 589–99. Дои:10.1016/0092-8674(87)90128-0. PMID 3677168. S2CID 31484517.
- ^ а б Артаванис-Цаконас С., Мацуно К., Фортини М.Э. (апрель 1995 г.). «Notch signaling». Наука. 268 (5208): 225–32. Bibcode:1995Научный ... 268..225A. Дои:10.1126 / science.7716513. PMID 7716513.
- ^ Гринвальд I, Рубин GM (январь 1992 г.). «Различия: роль межклеточного взаимодействия в установлении отдельных идентичностей для эквивалентных клеток». Клетка. 68 (2): 271–81. Дои:10.1016 / 0092-8674 (92) 90470-в.. PMID 1365402. S2CID 11901508.
- ^ Schroeter EH, Kisslinger JA, Kopan R (май 1998 г.). «Передача сигналов Notch-1 требует протеолитического высвобождения внутриклеточного домена, индуцированного лигандом». Природа. 393 (6683): 382–6. Bibcode:1998Натура.393..382С. Дои:10.1038/30756. PMID 9620803. S2CID 4431882.
- ^ Копан Р., Най Дж. С., Вайнтрауб Х (сентябрь 1994 г.). «Внутриклеточный домен Notch мыши: конститутивно активированный репрессор миогенеза, направленный на основную область спираль-петля-спираль MyoD». Разработка. 120 (9): 2385–96. PMID 7956819.
- ^ Struhl G, Fitzgerald K, Greenwald I (июль 1993 г.). «Внутренняя активность внутриклеточных доменов Lin-12 и Notch in vivo». Клетка. 74 (2): 331–45. Дои:10.1016 / 0092-8674 (93) 90424-о. PMID 8343960. S2CID 27966283.
- ^ Шеррингтон Р., Рогаев Е.И., Лян Ю., Рогаева Е.А., Левеск Дж., Икеда М. и др. (Июнь 1995 г.). «Клонирование гена, несущего миссенс-мутации, при семейной болезни Альцгеймера с ранним началом». Природа. 375 (6534): 754–60. Bibcode:1995Натура.375..754S. Дои:10.1038 / 375754a0. PMID 7596406. S2CID 4308372.
- ^ Гринвальд И.С., Штернберг П.В., Хорвиц HR (сентябрь 1983 г.). «Локус lin-12 определяет судьбы клеток у Caenorhabditis elegans». Клетка. 34 (2): 435–44. Дои:10.1016 / 0092-8674 (83) 90377-х. PMID 6616618. S2CID 40668388.
- ^ Остин Дж, Кимбл Дж (август 1989 г.). «Анализ транскриптов glp-1 и lin-12, гомологичных генов, необходимых для взаимодействия клеток во время развития C. elegans». Клетка. 58 (3): 565–71. Дои:10.1016/0092-8674(89)90437-6. PMID 2758467. S2CID 8514017.
- ^ Гринвальд I (декабрь 1985 г.). "lin-12, гомеотический ген нематод, гомологичен набору белков млекопитающих, который включает эпидермальный фактор роста". Клетка. 43 (3, часть 2): 583–90. Дои:10.1016/0092-8674(85)90230-2. PMID 3000611.
- ^ Shao L, Luo Y, Moloney DJ, Haltiwanger R (ноябрь 2002 г.). «O-гликозилирование повторов EGF: идентификация и начальная характеристика UDP-глюкозы: протеин-O-глюкозилтрансферазы». Гликобиология. 12 (11): 763–70. Дои:10.1093 / glycob / cwf085. PMID 12460944.
- ^ Тамура К., Танигучи Ю., Миногути С., Сакаи Т., Тун Т., Фурукава Т., Хондзё Т. (декабрь 1995 г.). «Физическое взаимодействие между новым доменом рецептора Notch и фактором транскрипции RBP-J kappa / Su (H)». Текущая биология. 5 (12): 1416–23. Дои:10.1016 / S0960-9822 (95) 00279-X. HDL:2433/202204. PMID 8749394. S2CID 18442572.
- ^ Вен А. П., Феррандо А. А., Ли В., Моррис Дж. П., Сильверман Л. Б., Санчес-Иризарри С. и др. (Октябрь 2004 г.). «Активирующие мутации NOTCH1 при остром лимфобластном лейкозе Т-клеток человека». Наука. 306 (5694): 269–71. Bibcode:2004Наука ... 306..269W. Дои:10.1126 / science.1102160. PMID 15472075. S2CID 24049536.
- ^ «Ген Entrez: NOTCH1, гомолог Notch 1, связанный с транслокацией (Drosophila)».
- ^ Маккеллар Ш., Тестер DJ, Ягубян М., Маджумдар Р., Акерман М. Дж., Сундт TM (август 2007 г.). «Новые мутации NOTCH1 у пациентов с заболеванием двустворчатого аортального клапана и аневризмами грудной аорты». Журнал торакальной и сердечно-сосудистой хирургии. 134 (2): 290–6. Дои:10.1016 / j.jtcvs.2007.02.041. PMID 17662764.
- ^ Танигаки К., Ногаки Ф., Такахаши Дж., Таширо К., Куроока Х., Хондзё Т. (январь 2001 г.). «Notch1 и Notch3 инструктивно ограничивают bFGF-чувствительные мультипотентные нейральные клетки-предшественники астроглиальной судьбой». Нейрон. 29 (1): 45–55. Дои:10.1016 / S0896-6273 (01) 00179-9. HDL:2433/150564. PMID 11182080. S2CID 17047028.
- ^ Gaiano N, Nye JS, Fishell G (май 2000 г.). «Идентичность радиальной глии обеспечивается передачей сигналов Notch1 в переднем мозге мышей». Нейрон. 26 (2): 395–404. Дои:10.1016 / S0896-6273 (00) 81172-1. PMID 10839358. S2CID 15861936.
- ^ Chambers CB, Peng Y, Nguyen H, Gaiano N, Fishell G, Nye JS (март 2001 г.). «Пространственно-временная селективность ответа на сигналы Notch1 в предшественниках переднего мозга млекопитающих». Разработка. 128 (5): 689–702. PMID 11171394.
- ^ Кейлани С., Сугая К. (июль 2008 г.). «Рилин индуцирует фенотип радиальной глии в клетках-предшественниках нейронов человека путем активации Notch-1». Биология развития BMC. 8 (1): 69. Дои:10.1186 / 1471-213X-8-69. ЧВК 2447831. PMID 18593473.
- ^ Sibbe M, Förster E, Basak O, Taylor V, Frotscher M (июль 2009 г.). «Reelin и Notch1 сотрудничают в развитии зубчатой извилины». Журнал неврологии. 29 (26): 8578–85. Дои:10.1523 / JNEUROSCI.0958-09.2009. ЧВК 6665659. PMID 19571148.
- ^ Ребай I, Флеминг Р.Дж., Фехон Р.Г., Чербас Л., Чербас П., Артаванис-Цаконас С. (ноябрь 1991 г.). «Специфические EGF-повторы Notch опосредуют взаимодействия с Delta и Serrate: значение Notch как многофункционального рецептора». Клетка. 67 (4): 687–99. Дои:10.1016/0092-8674(91)90064-6. PMID 1657403. S2CID 12643727.
- ^ де Селис Дж. Ф., Брей С. Дж. (март 2000 г.). «Домен Abruptex Notch регулирует негативные взаимодействия между Notch, его лигандами и Fringe». Разработка. 127 (6): 1291–302. PMID 10683181.
- ^ Блаумюллер CM, Ци Х., Загурас П., Артаванис-Цаконас С. (июль 1997 г.). «Внутриклеточное расщепление Notch приводит к гетеродимерному рецептору на плазматической мембране». Клетка. 90 (2): 281–91. Дои:10.1016 / s0092-8674 (00) 80336-0. PMID 9244302. S2CID 16544864.
- ^ Логит Ф., Бессия С., Броу С., ЛеБейл О., Жаррио С., Сейда Н.Г., Израиль А (июль 1998 г.). «Рецептор Notch1 конститутивно расщепляется фурин-подобной конвертазой». Труды Национальной академии наук Соединенных Штатов Америки. 95 (14): 8108–12. Bibcode:1998PNAS ... 95.8108L. Дои:10.1073 / пнас.95.14.8108. ЧВК 20937. PMID 9653148.
- ^ Кидд С., Либер Т. (июль 2002 г.). «Расщепление фурина не является требованием для функции Notch у Drosophila». Механизмы развития. 115 (1–2): 41–51. Дои:10.1016 / s0925-4773 (02) 00120-х. PMID 12049766. S2CID 10249987.
- ^ Рук Дж, Пан Д., Сюй Т., Рубин GM (август 1996). «KUZ, консервативный белок металлопротеаза-дезинтегрин, играющий две роли в нейрогенезе дрозофилы». Наука. 273 (5279): 1227–31. Bibcode:1996Научный ... 273.1227R. Дои:10.1126 / science.273.5279.1227. PMID 8703057. S2CID 10984937.
- ^ Пан Д., Рубин GM (июль 1997 г.). «Kuzbanian контролирует протеолитический процессинг Notch и опосредует латеральное торможение во время нейрогенеза дрозофилы и позвоночных». Клетка. 90 (2): 271–80. Дои:10.1016 / s0092-8674 (00) 80335-9. PMID 9244301. S2CID 18819481.
- ^ Вен Ц., Мецштейн М. М., Гринвальд I (декабрь 1997 г.). «SUP-17, белок ADAM Caenorhabditis elegans, родственный Drosophila KUZBANIAN, и его роль в передаче сигналов LIN-12 / NOTCH». Разработка. 124 (23): 4759–67. PMID 9428412.
- ^ Ховард Л., Глинн П. (1995). «Мембранно-ассоциированная металлопротеиназа, распознаваемая по характерному расщеплению основного белка миелина: анализ и выделение». Методы в энзимологии. 248: 388–95. Дои:10.1016/0076-6879(95)48025-0. PMID 7545777.
- ^ Lunn CA, Fan X, Dalie B, Miller K, Zavodny PJ, Narula SK, Lundell D (январь 1997 г.). «Очистка ADAM 10 из селезенки крупного рогатого скота как TNF-альфа-конвертазы». Письма FEBS. 400 (3): 333–5. Дои:10.1016 / с0014-5793 (96) 01410-х. PMID 9009225. S2CID 83810622.
- ^ Struhl G, Adachi A (сентябрь 2000 г.). «Требования к пресенилин-зависимому расщеплению notch и других трансмембранных белков». Молекулярная клетка. 6 (3): 625–36. Дои:10.1016 / с1097-2765 (00) 00061-7. PMID 11030342.
- ^ Ларссон С, Ларделли М, Уайт I, Лендаль У. (ноябрь 1994 г.). «Гены NOTCH1, 2 и 3 человека расположены в положениях хромосом 9q34, 1p13-p11 и 19p13.2-p13.1 в областях транслокации, связанной с неоплазией». Геномика. 24 (2): 253–8. Дои:10.1006 / geno.1994.1613. PMID 7698746.
- ^ Самедзима Х, Тории С., Косаки Р., Куросава К., Ёсихаши Х, Муроя К., Окамото Н., Ватанабе И., Кошо Т., Кубота М., Мацуда О, Гото М., Изуми К., Такахаши Т., Косаки К. (2007). «Скрининг мутаций синдрома Алажилля в генах JAG1 и NOTCH2 с использованием денатурирующей высокоэффективной жидкостной хроматографии». Генетическое тестирование. 11 (3): 216–27. Дои:10.1089 / gte.2006.0519. PMID 17949281.
- ^ Симпсон М.А., Ирвинг М.Д., Асилмаз Э., Грей М.Дж., Дафу Д., Элмсли Ф.В., Мансур С., Холдер С.Е., Мозг CE, Бертон Б.К., Ким К.Х., Паули Р.М., Афтимос С., Стюарт Х., Ким КА, Холдер-Эспинасс М. Робертсон SP, Дрейк WM, Trembath RC (март 2011 г.). «Мутации в NOTCH2 вызывают синдром Хайду-Чейни, нарушение тяжелой и прогрессирующей потери костной массы». Природа Генетика. 43 (4): 303–5. Дои:10,1038 / нг.779. PMID 21378985. S2CID 205357391.
Рекомендации
- Tax FE, Yeargers JJ, Thomas JH (март 1994). «Последовательность C. elegans lag-2 выявляет клеточный сигнальный домен, общий с Delta и Serrate из Drosophila». Природа. 368 (6467): 150–4. Bibcode:1994Натура.368..150Т. Дои:10.1038 / 368150a0. PMID 8139658. S2CID 4345508.
- Линдселл К.Э., Шаубер С.Дж., Бултер Дж., Вайнмастер Дж. (Март 1995 г.). «Jagged: лиганд млекопитающих, который активирует Notch1». Клетка. 80 (6): 909–17. Дои:10.1016/0092-8674(95)90294-5. PMID 7697721. S2CID 11720367.
- Kelley MR, Kidd S, Deutsch WA, Young MW (ноябрь 1987 г.). «Мутации, изменяющие структуру кодирующих последовательностей, подобных эпидермальному фактору роста, в локусе Notch Drosophila». Клетка. 51 (4): 539–48. Дои:10.1016/0092-8674(87)90123-1. PMID 3119223. S2CID 34338340.