Черепаший панцирь - Turtle shell


В черепаший панцирь представляет собой сложный щит для вентральной и дорсальной частей черепахи, черепахи и черепахи (все они классифицируются зоологами как «черепахи»), полностью покрывая все жизненно важные органы черепахи, а в некоторых случаях даже голову.[1] Он состоит из модифицированных костных элементов, таких как ребра, части таза и другие кости, которые встречаются у большинства рептилий. Кость панциря состоит как из скелетной, так и из кожная кость, показывая, что оболочка полностью покрыта оболочкой, вероятно, благодаря включению дермальной брони в грудную клетку.
Панцирь черепахи является важным исследованием не только из-за очевидной защиты, которую он обеспечивает животному, но и как инструмент идентификации, в частности, с окаменелостями, поскольку панцирь является одной из вероятных частей черепахи, которые могут выжить при окаменении. Следовательно, понимание структуры раковины у живых видов дает нам материал, сопоставимый с ископаемыми.
Оболочка черепаха, среди других видов, с древних времен использовался в качестве материала для широкого спектра небольших декоративных и практических предметов, но обычно его называют черепаховый.
Номенклатура оболочки
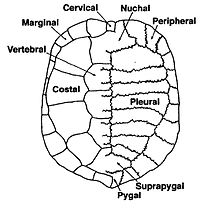
Панцирь черепахи состоит из множества костных элементов, обычно названных в честь схожих костей у других позвоночных, и ряда ороговевший щитки которые также имеют однозначные названия. Некоторые из костей, составляющих верхнюю часть панциря, панцирь, произошли от ветвей лопаток ключиц вместе с дорсальной и поверхностной миграцией клетры.[2] Вентральная поверхность называется пластрон.[3][4] К ним присоединяется зона, называемая мостом. Фактический шов между мостом и пластроном называется передним мостом.[5] В Pleurodires задний таз также является частью панциря, полностью срастаясь с ним. Это не так в Крипто-каталоги которые имеют плавающий таз.[3][4] Передняя стойка моста и задняя стойка моста являются частью пластрона, на панцире есть швы, в которые они вставляются, известные как швы панциря моста.[5]
Кости панциря названы в честь стандартных элементов позвоночных. Таким образом, панцирь состоит из 8 плевральных каналов с каждой стороны, они представляют собой комбинацию ребер и сросшейся дермальной кости. Снаружи в передней части раковины находится единственная затылочная кость, затем с каждой стороны проходит серия из 12 парных периферийных костей. В задней части панциря находится пигальная кость, а перед ней, за восьмыми плевральными отверстиями, расположена супрапигал.[3]

Между каждой плевральной оболочкой находится ряд нервных костей,[6] которые хотя всегда присутствуют, но не всегда видны,[7] у многих видов Pleurodire они погружены ниже плевральной полости.[8] Под нервной костью находится нервная дуга, которая образует верхнюю половину оболочки спинного мозга. Ниже этого остального позвоночника.[4] У некоторых видов черепах есть дополнительные кости, называемые мезопластами, которые расположены между панцирем и пластроном в области моста. Они присутствуют в большинстве Пеломедусид черепахи.[9]
Скелетные элементы пластрона также в основном парные. Спереди две эпипластры, за ними гипопластры. Они охватывают единичный энтопластрон. Они составляют переднюю половину пластрона, а гипопластрон содержит переднюю стойку моста. Задняя половина состоит из двух гипопластов (содержащих задний мостовидный протез), а задняя - из пары ксипипластов.[4][5]
Поверх костяных элементов находится ряд щитков, которые сделаны из кератин и очень похожи на ткань рога или ногтя. В центре панциря находятся 5 позвоночных щитков, а от них - 4 пары реберных щитков. По краю панциря 12 пар краевых щитков. Все эти щитки выровнены так, что швы между костями большей частью находятся посередине щитков выше. В передней части панциря может быть шейный щиток (иногда неправильно называемый затылочным щитком), однако наличие или отсутствие этого щитка сильно варьируется даже внутри вида.[4][9]
На пластроне есть две горловые щитки спереди, затем пара грудных, затем брюшные, бедренные и, наконец, анальные. Особый вариант - плевродиранские черепахи имеют межгулярный щиток между гулярами спереди, что дает им в общей сложности 13 пластральных щитков. По сравнению с 12 у всех криптодиранских черепах.[4][9]
Панцирь

В панцирь это спинной (сзади), выпуклая часть оболочечной конструкции черепаха, состоящий из окостеневших ребер животного, сросшихся с дермальной костью. В позвоночник и расширенные ребра сливаются путем окостенения кожные пластинки под кожей, чтобы сформировать твердую оболочку. Снаружи от кожи раковина покрыта щитки, которые представляют собой роговые пластины из кератин защищающие скорлупу от царапин и ушибов. А киль, гребень, идущий от передней части к задней части животного, присутствует у некоторых видов, они могут быть одиночными, парными или даже трехрядными. У большинства черепах панцирь относительно однороден по структуре, основные различия в форме и цвете составляют разновидности. Тем не менее мягкая оболочка черепахи, свиной нос черепахи и кожистая морская черепаха потеряли щитки и уменьшили окостенение панциря. Это оставляет оболочку покрытой только кожа.[11] Все это очень водные формы.
Эволюция панциря черепахи уникальна из-за того, что панцирь представляет собой преобразованные позвонки и ребра. В то время как у других четвероногих есть лопатка или лопатка Лопатка черепах находится за пределами грудной клетки, а внутри грудной клетки.[12][13] Раковины других четвероногих, таких как броненосцы, не связаны напрямую с позвоночником или грудной клеткой, что позволяет ребрам свободно перемещаться вместе с окружающей межреберной мышцей.[14] Однако анализ переходной окаменелости Eunotosaurus africanus показывает, что ранние предки черепах потеряли межреберные мышцы, обычно находящиеся между ребрами.[15]
Недавние открытия в поисках окаменелостей стволовых черепах вносят вклад в изучение эволюции панциря черепахи. Первой обнаруженной палеонтологической летописью, необходимой для построения модели эволюции и развития, была рептилия позднего триаса возрастом 214 миллионов лет из Германии и Таиланда. Проганохелис, который был отмечен как первая точка полного развития панциря и окостенения панциря у Testudines.[16]
Следующее феноменальное открытие китайской стволовой черепахи возрастом 220 миллионов лет, предшествовавшее Проганончелис к 6 миллионам лет Odontochelys semitestacea Ископаемое в Китае пролило свет на промежуточные стадии эволюции панциря черепахи, продемонстрировав частично сформированный спинной панцирь.[17] Одним из главных открытий было то, что O. semitestacea предоставили документацию о том, что пластрон развился раньше, чем структура панциря.[18] Тщательное изучение частично сформированного панциря показало сходство с полностью сформированным панцирем у кроновых черепах, такое как отсутствие межреберных мышц и ограниченная подвижность ребер. Кроме того, он состоит из модифицированных форм расширенных с боков и расширенных ребер без окостенения, подобных по структуре эмбриону современной черепахи.[16]
Добавление 260-миллионной пермской рептилии из Южной Африки Eunotosaurus africanus, предположительно самая ранняя стволовая черепаха, предшествовавшая O. semitestacea к 40 миллионам лет выявили ранние стадии эволюции панциря. Гистологические данные охарактеризованы E. africanus и Проганохелис ребра как гомологичные структуры, как расширенные, так и Т-образные в поперечном сечении.[19]
Лайсон предполагает, что эта морфология у Eunotosaurus africanus предполагает, что черепахи могут иметь роющий источник. Широкий торс дал начало панцирю черепахи, но во время Пермский период, расширенные ребра могли обеспечить большую устойчивость при рытье. Скелетное строение E. africanus по сравнению с сохранившимися ископаемыми суслик черепаха имеют аналогичные характеристики, адаптированные для противодействия ударам и силе, необходимым при копании. Например, E. africanus демонстрирует плечи и передние конечности, приспособленные к рытью, демонстрируя увеличенные мышцы, указанные в таких структурах, как их бугорок на заднем коракоиде и их большие и широкие концевые фаланги. Более того, фоссориальность могла помочь E. africanus пережить глобальное массовое вымирание, которое привело к исчезновению более девяноста процентов видов в конце пермского периода.[20][21] Эта особенность также обнаружена в стебле кроновых черепах, что предполагает возможность того, что поведение окаменелостей не было подвержено влиянию E. africanus , а скорее сыграл важную роль в ранней эволюции панцирных черепах. [20]
Самая последняя находка среднетриасовой стволовой черепахи предлагает как морфологическое, так и временное промежуточное звено по сравнению с Eunotosaurus africanus и Odontochelys semitestacea, способствуя построению временной шкалы эволюции панциря. 240 миллионов лет назад Паппочелис Ископаемые останки, найденные в Германии, демонстрируют такие же расширенные и Т-образные ребра, которые различаются по форме в зависимости от положения вдоль позвоночника.[22]
Было обнаружено, что гребешок карапаса играет важную роль в развитии панциря черепахи. Эмбриологический анализ показывает, что гребешок карапаса инициирует формирование панциря черепахи.[23] Это вызывает осевой останов, который вызывает дорсализацию ребер, перегруппировку плечевого пояса и его инкапсуляцию в грудную клетку и развитие панциря.[24] Odontochelys semitestacea представляет собой свидетельство остановки оси, наблюдаемой у эмбрионов, но без веерообразных ребер и панциря. Это предполагает, что примитивный карапациальный гребень функционировал по-другому и, должно быть, получил функцию опосредования развития ребер и панциря позже.[17][25] В Pax1 и Соник ежик ген (Шшш) служат ключевыми регуляторами во время развития позвоночника. Шшш экспрессия в нервной трубке важна для поддержания Pax1 экспрессия в вентральном склеротоме и, таким образом, играет ключевую роль в развитии карапациальных ребер. Генетические наблюдения Pax1 и Шшш также обеспечивают понимание экспрессии ключевых генов, которые потенциально могут быть ответственны за изменение морфологии черепах. [26]
Пластрон


Пластрон (множественное число: пластроны или пластра) - это почти плоская часть структуры оболочки черепаха, то, что можно было бы назвать брюшной или брюшной поверхностью раковины. Он также включает в свою структуру переднюю и заднюю стойки моста и перемычку оболочки.[4][5] Пластрон состоит из девяти костей, а две эпипластры на переднем крае пластрона гомологичны ключицам других четвероногих.[27] Остальные пластральные кости гомологичны гастралия других четвероногих. Пластрон был описан как экзоскелет, как остеодермы других рептилий; но в отличие от остеодерм, пластрон также обладает остеобласты, то остеоид, а надкостница.[28]
Эволюция пластрона остается более загадочной, хотя Жорж Кювье, французский естествоиспытатель и зоолог XIX века, писал, что пластрон развился в основном из грудины черепахи.[29] Это хорошо согласуется со знаниями, полученными в ходе эмбриологических исследований, показывающими, что изменения в путях развития ребер часто приводят к уродствам или потере пластрона. Это явление происходит при развитии черепахи, но вместо полной потери грудины черепаший план тела перепрофилирует кость в форму пластрона,[25] хотя другие исследования показывают, что эндохондральный грудина отсутствует и заменена экзоскелетным пластроном. Вентральные ребра фактически отсутствуют, их заменяет пластрон, если только гастралии, из которых развился пластрон, когда-то не были плавающими вентральными ребрами.[28] Во время эволюции черепах, вероятно, существовало разделение труда между ребрами, которые специализировались для стабилизации туловища, и брюшными мышцами, которые специализировались на дыхании, и эти изменения произошли за 50 миллионов лет до того, как панцирь полностью окостенел.[30]
Открытие ископаемых останков черепахи, Pappochelys rosinae, дает дополнительные сведения о том, как образовался пластрон. Паппочелис служит промежуточной формой между двумя ранними черепахами-стеблями, E. africanus и Одонточелис, последний из них обладает полностью сформированным пластроном. Вместо современного пластрона, Паппочелис имеет парную гастралию, как у E. africanus. Паппочелис отличается от своего предка, потому что гастралии имеют признаки того, что когда-то были слиты, на что указывают ископаемые образцы, на которых видны раздвоенные концы. Эти данные показывают постепенный переход от парной гастралии к парной и слитой гастралии и, наконец, к современному пластрону у этих трех образцов.[22]
В некоторых семьях есть петля между грудной и брюшной щитки, позволяющие черепахе почти полностью закрыться. В определенных разновидность пол тестудина можно определить по тому, является ли пластрон вогнутый, мужчина или выпуклый, женский. Это из-за брачного положения; Вогнутый пластрон самца позволяет ему легче садиться на самку во время совокупления.
Пластральные щитки соединяются по центральному шву по середине пластрона. Относительная длина сегментов шва может использоваться для определения вида черепаха. На пластроне шесть симметричных латерально симметричных пар щитков: гулярная, плечевая, грудная, брюшная, бедренная и анальная (идущая от головы к хвосту по шву); брюшной и гулярный швы щитка примерно одинаковой длины, бедренный и грудной швы примерно одинаковой длины.
В горловой щиток или же горная проекция на черепаха - самая передняя часть пластрона, нижняя сторона раковины. У некоторых черепах есть пара гуляр щитки, у других - единый нераздельный щиток. Гулярные щитки могут называться горным выступом, если они выступают как шпатель.


- Гулар (значения), анатомические образования горловины у других видов
Пластральная формула
Пластральная формула используется для сравнения размеров отдельных пластральных щитков (измеренных по среднему шву). Часто выделяют следующие пластральные щитки (с их сокращением):
межгулярный = интергул Gular = гуль плечевой = гул грудной = pect брюшной = abd бедренный = fem анальный = an
Сравнение пластральных формул позволяет различить эти два вида. Например, для восточная коробчатая черепаха, пластральная формула: an> abd> gul> pect> hum>
Пластроны черепах использовались древними китайцами в качестве гадание называется пластромантия. Смотрите также Кости оракула.
Источник
Исследования двух предков черепах, предок черепахи в горошек[32] и черепаха позднего триаса Проганохелис[33] постулировали предковую форму панциря. Было высказано предположение, что панцирь произошел от разбросанных остеодерм, покрывающих не полностью развитую внешнюю оболочку. Разбросанные остеодермы сначала сливались бы друг с другом, а затем сливались бы с нижележащими эндоскелетными позвонками и ребрами в процессе эволюции. Этот образец остеодермы, внедренный в кожу, также показан у многих других рептилий. Постепенное преобразование панциря черепахи от предковых форм к современным было предложено Майклом С. Й. Ли:[34] Серия трансформации панциря сначала начинается с некоторого небронированного парарептиля, затем бронированного парейазавра и, наконец, развивается в современных черепах с полностью развитым панцирем и перемещенной грудной клеткой. Эта трансформационная точка зрения на происхождение панциря черепах согласуется с дарвиновской точкой зрения, которая соглашается с теорией о том, что структуры потомков являются производными от постепенных, ступенчатых модификаций родительских.
Открытие ископаемых останков черепахи Eunotosaurus africanus (Возраст 260 миллионов лет) предоставил больше доказательств диапсидного происхождения черепах. В Eunotosaurus africanus был описан как «диапсидная рептилия в процессе вторичного анапсидирования»,[35] тем самым заполняя решающий пробел в эволюционном превращении черепах из диапсидных рептилий-предков в современных анапсидных черепах. Филогенетическое родство предков черепах можно резюмировать следующим образом: Eunotosaurus помещается в нижней части ствола черепашьего дерева, за которым следует Паппочелис и Одонточелис вдоль стебля черепахи и далее к другим черепахам в короне ".[36]
Щитки

Панцирь черепахи покрыт щитки которые сделаны из кератин. Отдельные щитки, показанные выше, имеют определенные имена и, как правило, одинаковы для разных видов черепах. Наземные черепахи не сбрасывают щитков. Новые щитки растут за счет добавления слоев кератина к основанию каждого щитка. Водный Chelonii сбрасывать отдельные щитки. Щиток эффективно формирует кожа над подлежащими костными структурами; между щитком и скелетом очень тонкий слой подкожной клетчатки. Щитки могут быть ярко окрашены у некоторых видов, но базальная окраска от серого до темно-коричневого цвета сверху; Пластральные щитки часто имеют основной цвет от белого до желтого.[нужна цитата ] Эмбриологическое исследование Moustakas-Verho и Черепанова показывает, что формирование паттерна пластральных щитков не зависит от паттерна панцирных щитков, предполагая, что панцирь и пластрон развивались отдельно.[37]
Появление щитков коррелирует с переходом четвероногих от водного к наземному образу жизни в каменноугольный период (340 млн лет назад).[38] В процессе эволюции от земноводных к наземным амниотам произошел переход в самых разных кожных структурах. Предки черепах, вероятно, отклонились от земноводных, чтобы развить роговой покров у своих ранних земных предков.[39]
Болезни

Септицемическая кожная язвенная болезнь (SCUD) / гниль скорлупы
При сепсисе кожно-язвенной болезни (SCUD) или «гнили скорлупы», первоначально описанной Kaplan (1957),[40] язвы оболочки могут быть поверхностными или глубокими. Язвы можно увидеть как на панцире, так и на ногах.[нужна цитата ] Известно, что заболевание вызывается различными бактерии или же грибы входя через какой-то истирание, в сочетании с бедными животноводство. Заболевание идентифицируется по его прогрессированию, и то, что начинается с язвенного поражения пластрона, приводит к септической инфекции, вызывающей деградацию печени и других органов.[41] Без лечения это приведет к смерти. Заболевание часто связано с бактериями, Citrobacter freundii.[нужна цитата ]
Черепах с язвенными поражениями панциря должны быть осмотрены и обработаны ветеринаром, так как язвы могут инфицироваться и проникнуть через панцирь. Скорлупу необходимо будет ежедневно чистить и удалять мертвые ткани. В случае бактериальных инфекций необходимы местные и / или инъекционные антибиотики. Глубокие язвы могут потребовать хирургического вмешательства с применением акрилового или стекловолоконного материала.
В отдаленных районах, где нет специализированных ветеринаров, были достигнуты некоторые успехи в лечении гниения скорлупы путем тщательного удаления мертвых тканей и дезинфекции участков с помощью пероксид водорода а затем обработать пораженные участки прополис растворяется в спирте в местах поражения скорлупы или применяется прополис мазь в местах поражения мягких тканей.[42][43]
Пирамидинг

Пирамидирование - это деформация раковины, иногда обнаруживаемая у пленников. черепахи, при котором оболочка растет неравномерно, что приводит к пирамида форма, лежащая в основе каждого щитка. Эта деформация может различаться по степени тяжести от едва заметной до опасной для жизни. Индийские звездные черепахи и некоторые другие виды более подвержены этому состоянию, чем другие.
Несколько факторов могут усугубить образование пирамид, однако это состояние тесно связано с доступностью влаги, которая способствует правильному распределению роста кератина, который составляет панцирь черепах. Если черепаха обезвожена или не может получить доступ к условиям, которые достаточно влажны, ороговевшие слои, которые в противном случае образовались бы на краях щитков, вырастают под существующим затвердевшим панцирем, вызывая эффект наложения, который подталкивает рост панциря вверх, а не наружу, и оказывает давление на скелет под оболочкой. В тяжелых случаях это приводит к уродству позвоночника и физическому развитию.
Другие факторы, которые также могут способствовать формированию пирамиды, включают чрезмерное потребление животных или овощей. белок; неадекватный кальций, UVB и / или витамин D3; плохое питание.[44][45][46]Пирамида также может быть видимым признаком метаболическая болезнь костей (MBD) у черепах. Как только возникла пирамида, ее нельзя повернуть вспять, хотя, если основные проблемы будут исправлены, любой последующий рост оболочки будет плавно формироваться.
Сломанные снаряды
Панцирь черепах может сломаться по естественным причинам, в результате несчастного случая или умышленно.[47] Если разрез не слишком велик, оболочку можно собрать, ввернув в нее болты, а затем соединив болты проволокой; в противном случае может потребоваться специальное устройство.[48]
Маски из черепашьего панциря

Житель островов Торресова пролива люди - единственная культура в мире, которая делает маски из панциря черепахи, известные как крар (черепаховый панцирь) на Западных островах и ле-оп (человеческое лицо) на Восточных островах.[49]
Смотрите также
Рекомендации
- ^ Кордеро, Джорджия (2017). "Панцирь черепахи". Текущая биология. 27 (5): R168 – R169. Дои:10.1016 / j.cub.2016.12.040. PMID 28267966.
- ^ Lyson, Tyler R .; Bhullar, Bhart-Anjan S .; Бевер, Гейб С .; Джойс, Уолтер Дж .; де Кейруш, Кевин; Абжанов, Архат; Готье, Жак А. (01.09.2013). «Гомология загадочной затылочной кости раскрывает новую реорганизацию плечевого пояса в эволюции панциря черепахи». Эволюция и развитие. 15 (5): 317–325. Дои:10.1111 / ede.12041. ISSN 1525–142X. PMID 24074278.
- ^ а б c Ромер, А. (1956) Остеология рептилий. Univ. Чикаго Пресс.
- ^ а б c d е ж грамм Зангерл Р. 1969. Панцирь черепахи. В: Gans, C., Bellairs, D.d'A. и Парсонс, Т. (Ред.). Биология рептилий, Том 1, Морфология А. Лондон: Academic Press. стр. 311–340
- ^ а б c d Томсон С., Уайт А. и Жорж А. (1997). "Повторная оценка Emydura lavarackorum: Идентификация живого ископаемого " (PDF). Воспоминания о музее Квинсленда. 42 (1): 327–336. Архивировано из оригинал (PDF) на 2015-06-09.CS1 maint: несколько имен: список авторов (связь)
- ^ Причард, P.C.H. (1988). «Обзор вариаций нервных костей среди современных хелонических видов с функциональной интерпретацией». Acta Zoologica Cracoviensa. 31 (26): 625–686.
- ^ Томсон С. и Жорж А. (1996). «Нервные кости у черепах-хелидов». Челонский заповедник и биология. 2: 82–86.
- ^ Родин, А.Г.Дж. И Миттермайер, Р. (1977). «Нервные кости черепах-челидов из Австралии и Новой Гвинеи» (PDF). Копея. 1977 (2): 370–372. Дои:10.2307/1443917. JSTOR 1443917.
- ^ а б c Pritchard, P.C.H. и P. Trebbau. 1984. Черепахи Венесуэлы. Вклад SSAR в герпетологию 2:.
- ^ Боянус, Л. Х. 1819. Anatome testudinis Europaeae. 178pp, 31 пластина
- ^ Чен, И. Х., У. Ян, М. А. Мейерс (2015). «Кожаный панцирь морской черепахи: прочный и гибкий биологический дизайн». Acta Biomaterialia. 28: 2–12. Дои:10.1016 / j.actbio.2015.09.023. PMID 26391496.CS1 maint: несколько имен: список авторов (связь)
- ^ Nagashima, H .; Sugahara, F .; Takechi, M .; Ericsson, R .; Кавасима-Охя, Й .; Нарита, Й .; Куратани, С. (2009). «Эволюция плана тела черепахи путем складывания и создания новых мышечных связей». Наука. 325 (5937): 193–196. Bibcode:2009Научный ... 325..193N. Дои:10.1126 / science.1173826. PMID 19590000. S2CID 206519888.
- ^ Wang, Z., J. Pascual-Anaya, A. Zadissa, W. Q. Li, Y. Niimura, Z. Y. Huang, C. Y. Li et al. 2013. Проект геномов мягкой черепахи и зеленой морской черепахи дает представление о развитии и эволюции специфического для черепах строения тела. " Природа Генетика 45:701-+.
- ^ Хирасава Т., Х. Нагашима и С. Куратани. 2013. Эндоскелетное происхождение панциря черепахи. Природные коммуникации 4.
- ^ Ли, М. С. Я. (2013). «Палеонтология: черепахи в переходный период». Текущая биология. 23 (12): R513 – R515. Дои:10.1016 / j.cub.2013.05.011. PMID 23787042.
- ^ а б Li, C .; Wu, X.-C .; Rieppel, O .; Wang, L.-T .; Чжао, Л.-Дж. (2008). «Предковая черепаха из позднего триаса на юго-западе Китая». Природа. 456 (7221): 497–501. Bibcode:2008Натура.456..497L. Дои:10.1038 / природа07533. PMID 19037315. S2CID 4405644.
- ^ а б Куратани, S (2011).«Эволюционная перспектива развития происхождения черепах: теория складывания панциря, основанная на характере развития карапациального гребня». Эволюция и развитие. 13 (1): 1–14. Дои:10.1111 / j.1525-142x.2010.00451.x. PMID 21210938.
- ^ Ли, Чун; У, Сяо-Чун; Риппель, Оливье; Ван, Ли-Тин; Чжао Ли-Цзюнь (27.11.2008). «Предковая черепаха из позднего триаса на юго-западе Китая». Природа. 456 (7221): 497–501. Bibcode:2008Натура.456..497L. Дои:10.1038 / природа07533. ISSN 0028-0836. PMID 19037315. S2CID 4405644.
- ^ Lyson, Tyler R .; Бевер, Гейб С .; Scheyer, Torsten M .; Hsiang, Allison Y .; Готье, Жак А. (2013). «Эволюционное происхождение панциря черепахи». Текущая биология. 23 (12): 1113–1119. Дои:10.1016 / j.cub.2013.05.003. PMID 23727095.
- ^ а б Лайсон, Т. Р., Б. С. Рубидж, Т. М. Шайер, К. Де Кейроз, Э. Р. Шахнер, Р. М. Смит, Дж. Бота-Бринк; и другие. (2016). «Ископаемое происхождение панциря черепахи» (PDF). Текущая биология. 26 (14): 1887–1894. Дои:10.1016 / j.cub.2016.05.020. PMID 27426515. S2CID 3935231.CS1 maint: несколько имен: список авторов (связь)
- ^ Chen, Z.-Q., and M. J. Benton. (2012). «Время и образец восстановления биоты после массового вымирания в конце пермского периода». Природа Геонауки. 5 (6): 375–383. Bibcode:2012Натуральная ... 5..375C. Дои:10.1038 / ngeo1475.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Schoch, Rainer R .; Сьюз, Ханс-Дитер (2015). «Среднетриасовая стволовая черепаха и эволюция строения тела черепахи». Природа. 523 (7562): 584–587. Bibcode:2015Натура.523..584S. Дои:10.1038 / природа14472. PMID 26106865. S2CID 205243837.
- ^ Рукес, Герберт (декабрь 1929 г.). «ИССЛЕДОВАНИЯ ПО ХЕЛОНСКОЙ ОСТЕОЛОГИИ, ЧАСТЬ I: ФЕРМЫ И ДУГИ АНАЛОГИИ В ХЕЛОНСКИХ ТЗАХ». Летопись Нью-Йоркской академии наук. 31 (1): 31–80. Дои:10.1111 / j.1749-6632.1929.tb55191.x. ISSN 0077-8923.
- ^ Риппель, Оливье (2017). Черепахи как обнадеживающие монстры: происхождение и эволюция. Издательство Индианского университета. п. 146. ISBN 9780253024756. OCLC 1037017014.
- ^ а б Хирасава, Тацуя; Паскуаль-Анайя, Хуан; Камезаки, Наоки; Танигучи, марийский; Мой, Канако; Куратани, Шигеру (01.05.2015). «Эволюционное происхождение панциря черепахи и его зависимость от осевой остановки эмбриональной грудной клетки». Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития. 324 (3): 194–207. Дои:10.1002 / jez.b.22579. ISSN 1552-5015. PMID 24898540.
- ^ Мустакас-Верхо, Жаклин; Thomas, C.T .; Гилберт, С. Ф. (2017). «Выкройка панциря черепахи». Текущее мнение в области генетики и развития. 45: 124–131. Дои:10.1016 / j.gde.2017.03.016. PMID 28570929 - через Elsevier Science Direct.
- ^ Гилберт, С. Ф .; Лоредо, Г. А .; Брукман, А .; Берк, А. С. (2001). «Морфогенез панциря черепахи: разработка новой структуры в эволюции четвероногих». Эволюция и развитие. 3 (2): 47–58. Дои:10.1046 / j.1525-142x.2001.003002047.x. PMID 11341674.
- ^ а б Рис, ритва; Каллонен, Аки; Себра-Томас, Юдифь; Гилберт, Скотт Ф. (10 мая 2016 г.). «Развитие пластрона черепахи, определяющего порядок строения скелета». Труды Национальной академии наук. 113 (19): 5317–5322. Дои:10.1073 / pnas.1600958113. ISSN 0027-8424. ЧВК 4868452. PMID 27114549.
- ^ МакКорд, Кейт; Канилья, Гвидо; Мустакас-Верхо, Жаклин Э .; Берк, Энн С. (01.05.2015). «Рассвет исследований келона: черепахи между сравнительной анатомией и эмбриологией в 19 веке». Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития. 324 (3): 169–180. Дои:10.1002 / jez.b.22587. HDL:10138/223805. ISSN 1552-5015. PMID 25074288.
- ^ Lyson, Tyler R .; Schachner, Emma R .; Бота-Бринк, Дженнифер; Scheyer, Torsten M .; Ламбертц, Маркус; Бевер, Г. С .; Рубидж, Брюс С .; де Кейруш, Кевин (07.11.2014). «Происхождение уникального вентиляционного аппарата черепах». Nature Communications. 5 (1): 5211. Bibcode:2014NatCo ... 5.5211L. Дои:10.1038 / ncomms6211. ISSN 2041-1723. PMID 25376734.
- ^ C.H. Эрнст, Р.Г.М. Альтенбург и Р.В. Барбур. "Террапен каролина". Нидерландский информационный фонд по биоразнообразию. Архивировано из оригинал 24 июля 2011 г.. Получено 12 февраля 2011.
- ^ Риппель, Оливье (13 марта 2017). Черепахи как обнадеживающие монстры: происхождение и эволюция. Блумингтон, Индиана. п. 157. ISBN 9780253025074. OCLC 962141060.
- ^ Гаффни, Юджин С. (1990). Сравнительная остеология триасовой черепахи Проганохелис. OCLC 263164288.
- ^ Ли, Майкл С. Ю. (февраль 1996 г.). «Взаимосвязь развития и происхождения черепах». Природа. 379 (6568): 812–815. Дои:10.1038 / 379812a0. ISSN 0028-0836. S2CID 29609847.
- ^ Бевер, Г. С .; Lyson, Tyler R .; Филд, Дэниел Дж .; Бхуллар, Бхарт-Аньян С. (сентябрь 2015 г.). «Эволюционное происхождение черепа черепахи». Природа. 525 (7568): 239–242. Дои:10.1038 / природа14900. ISSN 0028-0836. PMID 26331544. S2CID 4401555.
- ^ Риппель, Оливье (2017). Черепахи как обнадеживающие монстры: происхождение и эволюция. Издательство Индианского университета. п. 70. ISBN 9780253024756. OCLC 1037017014.
- ^ Мустакас-Верхо, Дж. Э., Р. Зимм, Дж. Себра-Томас, Н. К. Лемпиайнен, А. Каллонен, К. Л. Митчелл, К. Хамалайнен; и другие. (2014). «Происхождение и потеря периодического рисунка на панцире черепахи» (PDF). Разработка. 141 (15): 3033–3039. Дои:10.1242 / dev.109041. PMID 25053434. S2CID 7737357.CS1 maint: несколько имен: список авторов (связь)
- ^ Zimm, R .; Винекен, Бентли, Дж. (2017). «Экологическая причинность аномалий черепахи». Интегративная и сравнительная биология. 57 (6): 1303–1311. Дои:10.1093 / icb / icx066. PMID 28992039 - через Общество интегративной и сравнительной биологии.
- ^ Черепанов, Г. О. (2015). «Полиморфизм Щита как источник эволюционного развития панциря черепахи». Палеонтологический журнал. 49 (14): 1635–1644. Дои:10.1134 / S003103011514004X. S2CID 88095099 - через СПбГУ.
- ^ Каплан, Х. М. (1957). «Септицемическая, кожно-язвенная болезнь черепах». Proc. Панель по уходу за животными. 7: 273–277.
- ^ Мадер, Д. (2006) Медицина рептилий и хирургия, 2-е изд., Сондерс, ISBN 072169327X.
- ^ Б. Питцер 2018, личное сообщение, 25 апреля.
- ^ R Ilek 2019, личное сообщение, 1 июня).
- ^ Герлах, Дж (2004). «Влияние диеты на систематическую полезность панциря черепахи» (PDF). Биоразнообразие острова. Получено 17 июля 2019.
- ^ Иннис, Чарльз. «ОСНОВНОЕ ПРОИЗВОДСТВО И ПИТАНИЕ ХЕЛОНЦЕВ» (PDF). Cabi.Org.
- ^ "Пирамида в черепахах". www.reptilesmagazine.com.
- ^ Ли Моран (13 июня 2013 г.). «Игроки в гольф-клуб Delbrook в Висконсине забили черепаху до смерти в песчаном бункере - когда она пыталась отложить яйца». New York Daily News.
- ^ Viva Sarah Press (26 июля 2012 г.). «Израильское устройство исправляет сломанный панцирь черепахи».
- ^ «Художественные декорации. Искусство островов Торресова пролива». Художественная галерея Нового Южного Уэльса. Получено 7 января 2020.
внешняя ссылка
- Веб-сайт африканской черепахи, есть дополнительная информация о пирамидинге