ATOX1 - ATOX1





ATOX1 это медь металло-шаперон белок который закодирован ATOX1 ген в людях.[5][6] В млекопитающие, ATOX1 играет ключевую роль в меди гомеостаз поскольку он доставляет медь из цитозоль перевозчикам ATP7A и ATP7B.[7][8][9] Гомологичный белки содержатся в самых разных эукариоты, в том числе Saccharomyces cerevisiae как ATX1, и все содержат консервированный металл-связывающий домен.[7][10]
Функция
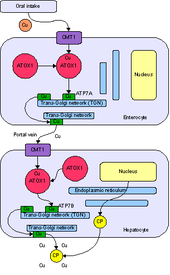
ATOX1 - это аббревиатура от полного названия Antioxidant Protein 1. номенклатура происходит из первоначальной характеристики, которая показала, что ATOX1 защищает клетки от активных форм кислорода. С тех пор была установлена основная роль ATOX1 как белка металло-шаперона меди, обнаруженного в цитоплазма эукариот.[7] Металлохаперон - важный белок, который играет роль в транспортировке и секвестрации металлов. Как белок секвестрации металлов, ATOX1 способен связывать свободные металлы. in vivo, чтобы защитить клетки от образования активные формы кислорода и неправильная металлизация металлопротеины. Как белок транспортировки металлов, ATOX1 отвечает за транспортировку меди из цитозоль к транспортерам АТФазы ATP7A и ATP7B, которые перемещают медь в сеть транс-Гольджи или секреторные пузырьки.[7][8][9] В Saccharomyces cerevisiae, Atx1 доставляет Cu (I) к гомологичному переносчику Ccc2. Доставка меди к транспортерам АТФазы жизненно важна для последующего внедрения меди в церулоплазмин, ферроксидаза, необходимая для метаболизма железа в аппарате Гольджи.[7]В дополнение к функции металло-шаперона в недавних сообщениях ATOX1 охарактеризован как циклин D1 фактор транскрипции.[8]
Координация конструкции и металла

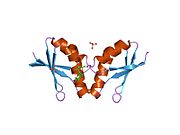
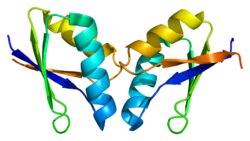
ATOX1 имеет ферродоксин -подобный βαββαβ складывается и координируется с Cu (I) через связывание MXCXXC мотив расположен между первым β-листом и α-спиралью.[7][9] Мотив связывания металла в значительной степени экспонируется растворителем в Апо -ATOX1 и a конформационный изменение индуцируется при координации с Cu (I).[9][10] Cu (I) скоординирован в искаженной линейной геометрии с серами цистин сформировать угол связи 120 °.[9] Общий заряд -1 первичной сфера координации стабилизируется через вторичная координационная сфера который содержит проксимальный положительно заряженный лизин.[9][10] ATOX1 также связывает Hg (II), Cd (II), Ag (I) и цисплатин через этот мотив, но физиологическая роль, если таковая имеется, еще не известна.[9]
Перенос металла

ATOX1 передает Cu (I) транспортерам ATP7A и ATP7B.[7][8][9] Перевод происходит через обмен лиганда механизм, при котором Cu (I) временно принимает 3-координатную геометрию с цистеиновыми лигандами из ATOX1 и ассоциированного переносчика.[9] Механизм обмена лиганда позволяет осуществлять обмен быстрее, чем распространение механизм и придает специфичность как для металла, так и для транспортера.[11] Поскольку обмен лигандом ускоряет этот перенос, а реакция имеет неглубокий термодинамический градиент, говорят, что она проходит ниже кинетический контроль, а не термодинамический контроль.[9][11]
Клиническое значение
Хотя в настоящее время нет известных болезни непосредственно связанных с неисправностью ATOX1, в настоящее время ведутся активные исследования в нескольких областях:
- Существует связь между уровнями ATOX1 и чувствительностью клеток к препаратам на основе Pt, таким как цисплатин.[9]
- Механизм тетратиомолибдат аммония [NH4]2MoS4 лечение болезни Вильсона пересматривается. Поскольку ATOX1 образует стабильный сложный тетратиомолибдат, он изучается как потенциальная терапевтическая мишень.[12][13]
использованная литература
- ^ а б c ГРЧ38: Ансамбль выпуск 89: ENSG00000177556 - Ансамбль, Май 2017
- ^ а б c GRCm38: выпуск Ensembl 89: ENSMUSG00000018585 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ "Ссылка на Mouse PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ Klomp LW, Lin SJ, Yuan DS, Klausner RD, Culotta VC, Gitlin JD (май 1997 г.). «Идентификация и функциональная экспрессия HAH1, нового человеческого гена, участвующего в гомеостазе меди». J Biol Chem. 272 (14): 9221–6. Дои:10.1074 / jbc.272.14.9221. PMID 9083055.
- ^ «Ген Entrez: гомолог антиоксидантного белка 1 ATOX1 ATX1 (дрожжи)».
- ^ а б c d е ж г Бертини И., Грей Х. Б., Штейфель Е. И., Валентин Дж. С. (2006). Биологическая неорганическая химия, структура и реакционная способность. Книги университетских наук. ISBN 978-1891389436.
- ^ а б c d Banci L (2013). Металломика и клетка. Дордрехт: Спрингер. ISBN 978-94-007-5561-1.
- ^ а б c d е ж г час я j k Maret W, Wedd A (2014). Связывание, транспорт и хранение ионов металлов в биологических клетках. [S.l.]: Королевское химическое общество. ISBN 978-1-84973-599-5.
- ^ а б c Boal AK, Rosenzweig AC (октябрь 2009 г.). «Структурная биология торговли меди». Химические обзоры. 109 (10): 4760–4779. Дои:10.1021 / cr900104z. ЧВК 2768115. PMID 19824702.
- ^ а б Робинсон, штат Нью-Джерси, Winge DR (7 июня 2010 г.). «Металло-шапероны меди». Ежегодный обзор биохимии. 79 (1): 537–562. Дои:10.1146 / annurev-biochem-030409-143539. ЧВК 3986808. PMID 20205585.
- ^ Альварес HM, Xue Y, Робинсон CD, Canalizo-Hernández MA, Marvin RG, Kelly RA, Mondragón A, Пеннер-Хан Дж., O'Halloran TV (январь 2010 г.). «Тетратиомолибдат подавляет белки транспортировки меди за счет образования металлических кластеров». Наука. 327 (5963): 331–334. Bibcode:2010Sci ... 327..331A. Дои:10.1126 / science.1179907. ЧВК 3658115. PMID 19965379.
- ^ Mjos KD, Orvig C (апрель 2014 г.). «Металлопрепараты в медицинской неорганической химии». Химические обзоры. 114 (8): 4540–4563. Дои:10.1021 / cr400460s. PMID 24456146.
внешние ссылки
- Человек ATOX1 расположение генома и ATOX1 страница сведений о генах в Браузер генома UCSC.
дальнейшее чтение
- Хунг И.Х., Касарено Р.Л., Лабесс Дж., Мэтьюз Ф.С., Гитлин Д.Д. (1998). «HAH1 - это медьсвязывающий белок с отдельными аминокислотными остатками, обеспечивающий гомеостаз меди и антиоксидантную защиту». J. Biol. Chem. 273 (3): 1749–54. Дои:10.1074 / jbc.273.3.1749. PMID 9430722.
- Ларин Д., Мекиос С., Дас К., Росс Б., Ян А.С., Гиллиам Т.С. (1999). «Характеристика взаимодействия между белками болезни Вильсона и Менкеса и цитоплазматическим медным шапероном, HAH1p». J. Biol. Chem. 274 (40): 28497–504. Дои:10.1074 / jbc.274.40.28497. PMID 10497213.
- Хамза И., Шефер М., Кломп Л.В., Гитлин Д.Д. (1999). «Взаимодействие шаперона меди HAH1 с белком болезни Вильсона необходимо для гомеостаза меди». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 96 (23): 13363–8. Bibcode:1999PNAS ... 9613363H. Дои:10.1073 / пнас.96.23.13363. ЧВК 23953. PMID 10557326.
- Вернимонт А.К., Хаффман Д.Л., Лэмб А.Л., О'Халлоран Т.В., Розенцвейг А.С. (2000). «Структурная основа переноса меди с помощью металлохаперона для белков болезни Менкеса / Вильсона». Nat. Struct. Биол. 7 (9): 766–71. Дои:10.1038/78999. PMID 10966647. S2CID 30817425.
- Боултвуд Дж, Стриксон А.Дж., Джабс Э.В., Ченг Дж.Ф., Фидлер С., Уэйнскоат Дж.С. (2000). «Физическое картирование человеческого гомолога ATX1 (HAH1) в критическую область 5q-синдрома в пределах 5q32 и непосредственно рядом с геном SPARC». Гм. Genet. 106 (1): 127–9. Дои:10.1007 / s004390051020. PMID 10982193.
- Уокер JM, Цивковский R, Луценко S (2002). «Металлохаперон Atox1 переносит медь в NH2-концевой домен белка болезни Вильсона и регулирует его каталитическую активность». J. Biol. Chem. 277 (31): 27953–9. Дои:10.1074 / jbc.M203845200. PMID 12029094.
- Мур С.Д., Хелмле К.Э., Прат Л.М., Кокс Д.В. (2003). «Тканевая локализация медного шаперона ATOX1 и его потенциальная роль в развитии болезни». Мамм. Геном. 13 (10): 563–8. Дои:10.1007 / s00335-002-2172-9. PMID 12420134. S2CID 19978302.
- Лю П.К., Келлер Д.М., Калер С.Г. (2004). «Геномная организация ATOX1, человеческого шаперона меди». BMC Genet. 4: 4. Дои:10.1186/1471-2156-4-4. ЧВК 150598. PMID 12594858.
- Strausak D, Howie MK, Firth SD, Schlicksupp A, Pipkorn R, Multhaup G, Mercer JF (2003). «Кинетический анализ взаимодействия шаперона меди Atox1 с сайтами связывания металлов белка Менкеса». J. Biol. Chem. 278 (23): 20821–7. Дои:10.1074 / jbc.M212437200. PMID 12679332.
- Ралле М, Луценко С, Блэкберн Нью-Джерси (2003). «Рентгеновская абсорбционная спектроскопия шаперона меди HAH1 выявила линейный двухкоординатный центр Cu (I), способный к образованию аддукта с экзогенными тиолами и фосфинами». J. Biol. Chem. 278 (25): 23163–70. Дои:10.1074 / jbc.M303474200. PMID 12686548.
- Луценко С., Цивковский Р., Уокер Дж. М. (2003). «Функциональные свойства человеческой медь-транспортной АТФазы ATP7B (белок болезни Вильсона) и регуляция металлохапероном Atox1». Анна. Акад. Наука. 986 (1): 204–11. Bibcode:2003НЯСА.986..204Л. Дои:10.1111 / j.1749-6632.2003.tb07161.x. PMID 12763797. S2CID 39325916.
- Вернимонт А.К., Яцуник Л.А., Розенцвейг А.С. (2004). «Связывание меди (I) белком болезни Вильсона и его медным шапероном». J. Biol. Chem. 279 (13): 12269–76. Дои:10.1074 / jbc.M311213200. PMID 14709553.
- Бранденбергер Р., Вей Х, Чжан С., Лей С., Мурадж Дж., Фиск Дж. Дж., Ли И, Сюй С., Фанг Р., Гуглер К., Рао М. С., Мандалам Р., Лебковски Дж., Стэнтон Л. В. (2005). «Характеристика транскриптома проясняет сигнальные сети, которые контролируют рост и дифференцировку ES-клеток человека». Nat. Биотехнология. 22 (6): 707–16. Дои:10.1038 / nbt971. PMID 15146197. S2CID 27764390.
- Anastassopoulou I, Banci L, Bertini I, Cantini F, Katsari E, Rosato A (2004). «Структура раствора апо и медь (I)-загруженного металло-шаперона человека HAH1». Биохимия. 43 (41): 13046–53. Дои:10.1021 / bi0487591. PMID 15476398.
- Banci L, Bertini I, Ciofi-Baffoni S, Chasapis CT, Hadjiliadis N, Rosato A (2005). «ЯМР-исследование взаимодействия между шапероном меди (I) человека и вторым и пятым металлсвязывающими доменами белка Менкеса». FEBS J. 272 (3): 865–71. Дои:10.1111 / j.1742-4658.2004.04526.x. PMID 15670166. S2CID 1130281.
- Джени В., Ито С., Вендт М., Градек К., Ушио-Фукаи М., Харрисон Д.Г., Фукаи Т. (2005). «Роль антиоксиданта-1 во внеклеточной функции и экспрессии супероксиддисмутазы». Circ. Res. 96 (7): 723–9. Дои:10.1161 / 01.RES.0000162001.57896.66. PMID 15761197.
Галерея PDB | |
|---|---|
|