Дорсолатеральная префронтальная кора - Dorsolateral prefrontal cortex
| Дорсолатеральная префронтальная кора | |
|---|---|
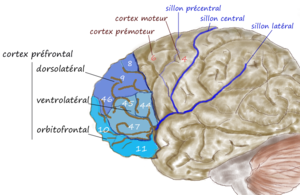 Иллюстрация префронтальной области мозга | |
| Подробности | |
| Идентификаторы | |
| латинский | Cortex praefrontalis dorsolateralis |
| FMA | 276189 |
| Анатомические термины нейроанатомии | |
В дорсолатеральная префронтальная кора (DLPFC или же DL-PFC) - это область в префронтальная кора мозга человека и других приматов. Это одна из самых недавно полученных частей человеческого мозга. Он проходит длительный период созревания, который длится до зрелого возраста.[1] DLPFC - это не анатомическая структура, а скорее функциональная. Он расположен в средней лобной извилине человека (т. Е. В боковой части Площадь Бродмана (БА) 9 и 46[2]). У макак он находится вокруг основной борозды (т. Е. В зоне Бродмана 46[3][4][5]). Другие источники считают, что DLPFC анатомически приписывают BA 9 и 46.[6] и BA 8, 9 и 10.[1]
DLPFC имеет связь с орбитофронтальная кора, так же хорошо как таламус, части базальный ганглий (в частности, спинной хвостатое ядро ), гиппокамп, а также первичные и вторичные зоны ассоциации неокортекс (включая заднюю височную, теменную и затылочную области).[7] DLPFC также является конечной точкой для дорсальный путь (ручей),[8] который связан с тем, как взаимодействовать со стимулами.
Важной функцией DLPFC является исполнительные функции, Такие как рабочая память, когнитивная гибкость,[9] планирование, сдерживание и абстрактные рассуждения.[10] Однако DLPFC не несет исключительной ответственности за исполнительные функции. Вся сложная умственная деятельность требует дополнительных корковых и подкорковых цепей, с которыми связан DLPFC.[11] DLPFC также является высшей корковой областью, которая участвует в двигательное планирование, организация и регулирование.[11]
Структура
Поскольку DLPFC состоит из пространственно-селективных нейроны, он имеет нейронную схему, которая охватывает весь спектр подфункций, необходимых для выполнения интегрированного ответа, таких как: сенсорный ввод, удержание в краткосрочная память, и моторную сигнализацию.[12] Исторически DLPFC определялась его связью с: височная кора, то задняя теменная кора, то передняя и задняя поясная извилина, то премоторная кора, то ретроспленальный кора головного мозга, а неоцеребеллум.[1] Эти соединения позволяют DLPFC регулировать деятельность этих регионов, а также получать информацию из этих регионов и регулироваться ими.[1]
Функция
Основные функции
DLPFC известен своим участием в исполнительных функциях, что является общим термином для управления когнитивные процессы,[13] включая рабочая память, когнитивная гибкость,[14] и планирование.[15] Несколько задач были очень важны в исследованиях DLPFC, таких как Задача A-not-B, то задача с отложенным ответом и задачи поиска объектов.[1] Поведенческая задача, которая наиболее сильно связана с DLPFC, - это комбинированная задача A-not-B / отложенного ответа, в которой субъект должен найти скрытый объект после определенной задержки. Эта задача требует хранения информации (рабочая память ), что считается одной из функций DLPFC.[1] Важность DLPFC для рабочей памяти была подтверждена исследованиями на взрослых макаках. Повреждения, которые разрушили DLPFC, нарушили выполнение макаками задачи A-not-B / отложенного ответа, тогда как поражения других частей мозга не ухудшили их выполнение этой задачи.[1]
DLPFC не требуется для памяти отдельного элемента. Таким образом, повреждение дорсолатеральной префронтальной коры не нарушает память распознавания.[16] Тем не менее, если нужно сравнить два элемента по памяти, потребуется задействовать DLPFC. Люди с поврежденным DLPFC не могут идентифицировать изображение, которое они видели, через некоторое время, когда им предоставляется возможность выбрать одно из двух изображений.[16] Более того, эти предметы тоже не смогли Тест сортировки карточек в Висконсине так как они теряют отслеживать в настоящее время верного правила и настойчиво организовывать свои карты в ранее правильном правиле.[17] Кроме того, поскольку DLPFC занимается тестированием мыслей и реальности наяву, он не активен, когда человек спит.[17] Точно так же DLPFC чаще всего связана с дисфункцией влечения, внимания и мотивации.[18] Пациенты с незначительным повреждением DLPFC не проявляют интереса к своему окружению и лишены спонтанности в речи, а также в поведении.[18] Пациенты также могут быть менее внимательны, чем обычно, к людям и событиям, которых они знают.[18] Повреждение этой области у человека также приводит к отсутствию мотивации делать что-то для себя и / или для других.[18]
Принимать решение
DLPFC занимается как рискованными, так и моральными принимать решение; когда людям приходится принимать моральные решения, например, как распределять ограниченные ресурсы, активируется DLPFC.[19] Этот регион также активен, когда интересуют затраты и выгоды альтернативных вариантов.[20] Точно так же, когда присутствуют варианты выбора альтернатив, DLPFC вызывает предпочтение наиболее справедливого варианта и подавляет искушение максимизировать личную выгоду.[21]
Рабочая память
Рабочая память - это система, которая активно хранит в уме несколько фрагментов преходящей информации, где ими можно манипулировать. DLPFC важен для рабочей памяти;[22] снижение активности в этой области коррелирует с низкой производительностью при выполнении задач с рабочей памятью.[23] Однако в рабочую память вовлечены и другие области мозга.[24]
В настоящее время ведутся дискуссии о том, специализируется ли DLPFC на определенных тип рабочей памяти, а именно вычислительных механизмов для мониторинга и манипулирования элементами, или если она имеет определенный содержание, а именно зрительно-пространственная информация, которая позволяет мысленно представить координаты в пространственной области.[22]
Также были некоторые предположения, что функция DLPFC в вербальной и пространственной рабочей памяти передается в левое и правое полушарие соответственно. Смит, Йонидес и Коппе (1996)[25] наблюдали латерализацию активаций DLPFC во время вербальной и зрительной рабочей памяти. Задачи вербальной рабочей памяти в основном активировали левую DLPFC, а задачи визуальной рабочей памяти в основном активировали правую DLPFC. Мерфи и др. (1998)[26] также обнаружили, что задачи вербальной рабочей памяти активируют правую и левую DLPFC, тогда как задачи пространственной рабочей памяти преимущественно активируют левую DLPFC. Reuter-Lorenz et al. (2000)[27] обнаружили, что активация DLPFC показала заметную латерализацию вербальной и пространственной рабочей памяти у молодых людей, тогда как у пожилых людей эта латерализация была менее заметной. Было высказано предположение, что это уменьшение латерализации могло быть связано с привлечением нейронов из противоположного полушария, чтобы компенсировать снижение нейронов с возрастом.
Вторичные функции
DLPFC также может быть причастен к обману и лжи,[28] который, как считается, препятствует нормальной тенденции говорить правду. Исследования также показывают, что использование ТМС на DLPFC может помешать человеку лгать или говорить правду.[29]
Кроме того, подтверждающие данные свидетельствуют о том, что DLPFC может также играть роль в корректировке поведения, вызванной конфликтом, например, когда человек решает, что делать, когда сталкивается с конфликтующими правилами.[30] Один из способов, которым это было проверено, - это Струп тест,[31] в котором испытуемым показывают название цвета, напечатанное цветными чернилами, а затем просят как можно быстрее назвать цвет чернил. Конфликт возникает, когда цвет чернил не совпадает с названием напечатанного цвета. Во время этого эксперимента отслеживание активности мозга субъектов показало заметную активность в DLPFC.[31] Активация DLPFC коррелировала с поведенческими характеристиками, что свидетельствует о том, что эта область поддерживает высокие требования задачи по разрешению конфликта и, таким образом, теоретически играет роль в принятии контроля.[31]
DLPFC также может быть связан с человеческим интеллектом. Однако даже когда обнаруживается корреляция между DLPFC и человеческим интеллектом, это не означает, что весь человеческий интеллект является функцией DLPFC. Другими словами, этот регион можно отнести к общему интеллекту в более широком масштабе, а также к очень специфическим ролям, но не ко всем ролям. Например, используя исследования изображений, такие как ДОМАШНИЙ ПИТОМЕЦ и фМРТ указать на участие DLPFC в дедуктивный, силлогистический рассуждения.[32] В частности, когда вы участвуете в деятельности, которая требует силлогистический рассуждая, левые области DLPFC особенно и постоянно активны.[32]
DLPFC также может быть вовлечен в вызванную угрозой тревогу.[33] В одном эксперименте участников попросили оценить себя как заторможенное или нет. Более того, те, кто оценил себя как заторможенный в поведении, продемонстрировали большую тоническую (покоящуюся) активность в правой задней части DLPFC.[33] Такую деятельность можно увидеть насквозь Электроэнцефалограмма (ЭЭГ) записи. Лица с заторможенным поведением с большей вероятностью будут испытывать чувство стресса и беспокойства, когда сталкиваются с особенно опасной ситуацией.[33] Согласно одной из теорий, восприимчивость к тревоге может увеличиваться в результате присутствующей бдительности. Доказательства этой теории включают: нейровизуализация исследования, демонстрирующие активность DLPFC, когда человек проявляет бдительность.[33] В частности, предполагается, что тревога, вызванная угрозой, также может быть связана с недостатками в решении проблем, что приводит к неопределенности.[33] Когда человек испытывает неуверенность, активность DLPFC возрастает. Другими словами, такая активность может быть связана с тревогой, вызванной угрозой.
Социальное познание
Среди префронтальных долей DLPFC, по-видимому, наименее прямое влияние на социальное поведение, но придает ясность и организованность. социальное познание.[10] DLPFC, по-видимому, вносит свой вклад в социальные функции, выполняя свою главную специальность - исполнительные функции, например, при работе со сложными социальными ситуациями.[10] Социальные области, в которых исследуется роль DLPFC, включают, среди прочего, социальную перспективу.[7] и выводя намерения других людей,[7] или же теория разума;[10] подавление эгоистичного поведения,[7][34] и приверженность в отношениях.[35]
Отношение к нейротрансмиттерам
Поскольку DLPFC претерпевает длительные изменения созревания, одним из изменений, которые приписывают DLPFC для достижения ранних когнитивных достижений, является повышение уровня нейротрансмиттера. дофамин в DLPFC.[1] В исследованиях, в которых рецепторы дофамина у взрослых макак были заблокированы, было замечено, что у взрослых макак был дефицит в задаче A-not-B, как если бы DFPLC был полностью отключен. Аналогичная ситуация наблюдалась, когда макакам вводили инъекции MPTP, который снижает уровень дофамина в DLPFC.[1] Несмотря на то, что физиологических исследований на предмет участия холинергические действия в подкорковых областях поведенческие исследования показывают, что нейромедиатор ацетилхолин необходим для работы функции памяти DLPFC.[36]
Клиническое значение
Шизофрения
Шизофрения частично может быть связано с недостаточной активностью лобной доли.[17] Дорсолатеральная префронтальная кора особенно неактивна, когда человек страдает хронической шизофренией.[нужна цитата ] Шизофрения также связана с недостатком нейромедиатора дофамина в лобная доля.[нужна цитата ][17] Дисфункции DLPFC уникальны среди пациентов с шизофренией, поскольку те, у кого диагностирована депрессия, обычно не имеют такой же аномальной активации в DLPFC во время задач, связанных с рабочей памятью.[23] Рабочая память зависит от стабильности и функциональности DLPFC, поэтому сниженная активация DLPFC приводит к тому, что пациенты с шизофренией плохо выполняют задачи, связанные с рабочей памятью. Низкая производительность способствует дополнительным ограничениям емкости рабочей памяти, превышающим ограничения для нормальных пациентов.[37][нужна цитата ] Когнитивные процессы, которые в значительной степени имеют дело с DLPFC, такие как память, внимание и обработка более высокого порядка, - это функции, которые однажды искажены, способствуют заболеванию.[23][страница нужна ]
Депрессия
Наряду с областями мозга, такими как лимбическая система, дорсолатеральная префронтальная кора в значительной степени занимается сильное депрессивное расстройство (MDD). DLPFC может способствовать депрессии из-за участия в расстройстве на эмоциональном уровне во время стадии подавления.[38] Хотя задачи с рабочей памятью обычно активируют DLPFC,[39] его уменьшение объема серого вещества коррелирует со снижением активности. DLPFC также может быть связан с вентромедиальная префронтальная кора в своих функциях с депрессией.[38] Это может быть связано с тем, что когнитивные функции DLPFC могут также включать эмоции, а эмоциональные эффекты VMPFC также могут включать самосознание или саморефлексию. Повреждение или повреждение DLPFC также может привести к усилению выраженности симптомов депрессии.
Стресс
Воздействие тяжелого стресс также может быть связано с повреждением DLPFC.[40] В частности, острый стресс отрицательно влияет на высшую когнитивную функцию, известную как рабочая память (WM), который также считается функцией DLPFC.[40] В эксперименте исследователи использовали функциональная магнитно-резонансная томография (фМРТ) для записи нейронной активности у здоровых людей, которые участвовали в выполнении заданий в стрессовой среде.[40] Когда стресс успешно воздействовал на испытуемых, их нейронная активность показывала снижение активности, связанной с рабочей памятью, в DLPFC.[40] Эти данные не только демонстрируют важность области DLPFC в отношении стресса, но также предполагают, что DLPFC может играть роль в других психических расстройствах. У пациентов с пост-травматическое стрессовое растройство (ПТСР), например, ежедневные сеансы правого дорсолатерального префронтального повторяющаяся транскраниальная магнитная стимуляция (rTMS) с частотой 10 Гц приводили к более эффективной терапевтической стимуляции.[41]
Злоупотребление алкоголем или наркотиками
Злоупотребление психоактивными веществами или расстройство, связанное с употреблением психоактивных веществ (SUD), может коррелировать с дисфункцией дорсолатеральной префронтальной коры.[42] Было показано, что те, кто злоупотребляет наркотиками, проявляют повышенное рискованное поведение, что, возможно, коррелирует с дисфункцией DLPFC. Функции исполнительного контроля DLPFC у лиц, демонстрирующих злоупотребление наркотиками, могут иметь меньшую связь с такими областями фактора риска, как передняя поясная кора и островок.[42] Эта ослабленная связь проявляется даже у здоровых субъектов, таких как пациент, который продолжал принимать рискованные решения с разрывом между их DLPFC и островком. Поражение DLPFC может привести к безответственности и свободе от запретов,[43] и злоупотребление наркотиками может вызвать такую же реакцию желания или вдохновения участвовать в смелой деятельности.
Алкоголь
Алкоголь нарушает функцию префронтальной коры.[44] Поскольку передняя поясная часть коры головного мозга работает, чтобы подавить любое несоответствующее поведение, обрабатывая информацию в исполнительной сети DLPFC,[44] как отмечалось ранее, это нарушение связи может привести к выполнению этих действий. Было показано, что в задаче, известной как Кембриджская задача риска, участники SUD имеют более низкую активацию их DLPFC. В частности, в тесте, связанном с алкоголизмом, в задаче под названием «Колесо фортуны» (WOF) подростки с семейным анамнезом алкоголизма имели более низкую активацию DLPFC.[42] У подростков, в семье которых не было алкоголизма, не наблюдалось такого же снижения активности.[нужна цитата ]
Смотрите также
- Внимание против памяти в префронтальной коре
- Внимательный сдвиг
- Площадь Бродмана 46
- Когнитивный контроль
- Дорсомедиальная префронтальная кора
- Мезокортикальный путь
- Тест сортировки карточек Висконсина
- Лобно-теменная сеть
Рекомендации
- ^ а б c d е ж грамм час я Лучиана, изд. Чарльз А. Нельсон; Моника (2001). Справочник по когнитивной нейробиологии развития. Кембридж, Массачусетс [u.a.]: MIT Press. ISBN 978-0-262-14073-7.CS1 maint: дополнительный текст: список авторов (связь)[страница нужна ]
- ^ Бродманн, 1909 г.
- ^ Уокер, 1940
- ^ Хоши, Э. (2001). «Функциональная специализация в дорсолатеральной префронтальной коре: обзор анатомических и физиологических исследований нечеловеческих приматов». Нейробиологические исследования. 54 (2): 73–84. Дои:10.1016 / j.neures.2005.10.013. PMID 16310877. S2CID 17212229.
- ^ Милиус, В. (2013). «Определение DLPFC и M1 в соответствии с анатомическими ориентирами для управляемой стимуляции мозга: надежность, точность и влияние пола и возраста». NeuroImage. 78: 224–32. Дои:10.1016 / j.neuroimage.2013.03.061. PMID 23567888. S2CID 1535066.
- ^ Чеслик, Э. (2013). "Есть ли" один "DLPFC в управлении когнитивными действиями? Доказательства гетерогенности от парцелляции на основе совместной активации". Кора головного мозга. 23 (11): 2677–2689. Дои:10.1093 / cercor / bhs256. ЧВК 3792742. PMID 22918987.
- ^ а б c d Мосс, Симмон. «Дорсолатеральная префронтальная кора». Психлопедия. Архивировано из оригинал 11 ноября 2013 г.. Получено 11 ноября 2013.
- ^ Такахаши, Эми; Оки, Кеничи; Ким, Дэ-Шик (15.01.2013). «Диссоциация и конвергенция дорсального и вентрального визуальных потоков в префронтальной коре человека». NeuroImage. 65: 488–498. Дои:10.1016 / j.neuroimage.2012.10.002. ISSN 1053-8119. ЧВК 4384683. PMID 23063444.
- ^ Каплан, Дж. Т .; и другие. (2016). «Нейронные корреляты сохранения политических убеждений перед лицом контрдоказательства». Научные отчеты. Природа. 6: 39589. Bibcode:2016НатСР ... 639589K. Дои:10.1038 / srep39589. ЧВК 5180221. PMID 28008965.
- ^ а б c d Редакторы: Брюс Л. Миллер и Джеффри Л. Каммингс (2007). Лобные доли человека: функции и нарушения. Гилфорд Пресс. п. 355. ISBN 978-1-59385-329-7.CS1 maint: дополнительный текст: список авторов (связь)
- ^ а б Джеймс Б. Хейл; Екатерина А. Фиорелло (2004). Школьная нейропсихология: Справочник практикующего врача. Guilford Press. стр.64 –65. ISBN 978-1593850111.
- ^ Гольдман-Ракич, Патриция С. (1995). «Архитектура префронтальной коры и центрального исполнительного органа». Летопись Нью-Йоркской академии наук. 769 (1): 71–83. Bibcode:1995НЯСА.769 ... 71Г. Дои:10.1111 / j.1749-6632.1995.tb38132.x. PMID 8595045. S2CID 40870968.
- ^ Эллиотт Р. (2003). Исполнительные функции и их нарушения. Британский медицинский бюллетень. (65); 49–59
- ^ Monsell S (2003). «Переключение задач». Тенденции в когнитивных науках. 7 (3): 134–140. Дои:10.1016 / S1364-6613 (03) 00028-7. PMID 12639695. S2CID 12847511.
- ^ Чан, Р. К. К., Шум, Д., Тулопулу, Т. и Чен, Е. Ю. Х., Р.; Шум, Д; Toulopoulou, T; Чен, Э (2008). «Оценка исполнительных функций: обзор инструментов и выявление критических проблем». Архив клинической нейропсихологии. 2. 23 (2): 201–216. Дои:10.1016 / j.acn.2007.08.010. PMID 18096360.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Goldman-Rakic, под редакцией Джеральдин Доусон, Курта В. Фишера; предисловие Патрисии С. (1994). Человеческое поведение и развивающийся мозг. Нью-Йорк: Guilford Press. ISBN 978-0898620924.CS1 maint: дополнительный текст: список авторов (связь)[страница нужна ]
- ^ а б c d Картер, Рита (1999). Отображение разума. Беркли: Калифорнийский университет Press. ISBN 978-0520224612.[страница нужна ]
- ^ а б c d Миллер, Брюс Л. (1999). Лобные доли человека. Нью-Йорк, Нью-Йорк: Гилфорд Пресс.[страница нужна ]
- ^ Greene, J.D .; Соммервилл, РБ; Nystrom, LE; Дарли, JM; Коэн, JD (2001). «Исследование ФМРТ эмоциональной вовлеченности в моральное суждение». Наука. 293 (5537): 2105–8. Bibcode:2001Sci ... 293.2105G. Дои:10.1126 / science.1062872. PMID 11557895. S2CID 1437941.
- ^ Дункан, Джон; Оуэн, Адриан М (2000). «Общие области лобной доли человека задействованы различными когнитивными потребностями». Тенденции в неврологии. 23 (10): 475–83. Дои:10.1016 / S0166-2236 (00) 01633-7. PMID 11006464. S2CID 15268389.
- ^ Knoch, D .; Фер, Э. (2007). «Сопротивление силе соблазнов: правая префронтальная кора и самоконтроль» (PDF). Летопись Нью-Йоркской академии наук. 1104 (1): 123–34. Bibcode:2007 НЯСА1104..123К. Дои:10.1196 / летопись.1390.004. PMID 17344543. S2CID 1745375.
- ^ а б Барби А.К., Кенигс М., Графман Дж. (Май 2013 г.). «Дорсолатеральный префронтальный вклад в рабочую память человека» (PDF). Кора. 49 (5): 1195–1205. Дои:10.1016 / j.cortex.2012.05.022. ЧВК 3495093. PMID 22789779.
- ^ а б c Хемби, под редакцией С.Э .; Бан, С. (2006). Функциональная геномика и протеомика в клинической нейробиологии (1. ред.). Амстердам: Academic Press. ISBN 978-0444518538.CS1 maint: дополнительный текст: список авторов (связь)
- ^ Рыцарь, изд. Дональд Т. Стусс; Роберт Т. (2002). Принципы функции лобной доли ([Online-Ausg.]. Ред.). Оксфорд [u.a.]: Oxford Univ. Нажмите. ISBN 978-0195134971.CS1 maint: дополнительный текст: список авторов (связь)
- ^ Smith, E. E .; Jonides, J .; Коппе, Р. А. (1996). «Разделение вербальной и пространственной рабочей памяти с помощью ПЭТ». Кора головного мозга. 6 (1): 11–20. Дои:10.1093 / cercor / 6.1.11. PMID 8670634.
- ^ Мерфи, Д. Г. М., Дейли, Э. М., Ван Амельсвоорт, Т., Робертсон, Д., Симмонс, А., и Кричли, Г. Д. (1998). Функциональная нейроанатомическая диссоциация вербальной, зрительной и пространственной рабочей памяти. Исследование шизофрении. http://doi.org/10.1016/S0920-9964(97)88566-0
- ^ Рейтер-Лоренц; Jonides, J .; Smith, E. E .; Hartley, A .; Миллер, А .; Marshuetz, C .; Коппе (2000). «Возрастные различия во фронтальной латерализации вербальной и пространственной рабочей памяти, выявленные методом ПЭТ». Журнал когнитивной неврологии. 12 (1): 174–187. CiteSeerX 10.1.1.544.9130. Дои:10.1162/089892900561814. PMID 10769314. S2CID 10138007.
- ^ Ито, Аяхито; Абэ, Нобухито; Фудзи, Тошикацу; Хаяси, Акико; Уэно, Ая; Мугикура, Сюндзи; Такахаши, Шоки; Мори, Эцуро (2012). «Вклад дорсолатеральной префронтальной коры в подготовку к обману и установлению истины». Исследование мозга. 1464: 43–52. Дои:10.1016 / j.brainres.2012.05.004. PMID 22580084. S2CID 37185586.
- ^ Картон, Инга; Бахманн, Талис (2011). «Влияние префронтальной транскраниальной магнитной стимуляции на спонтанное правдивое высказывание». Поведенческие исследования мозга. 225 (1): 209–14. Дои:10.1016 / j.bbr.2011.07.028. PMID 21807030. S2CID 25457056. Сложить резюме – Новый ученый (6 сентября 2011 г.).
- ^ Mansouri, F.A .; Бакли, М. Дж .; Танака, К. (2007). «Мнемоническая функция дорсолатеральной префронтальной коры в индуцированной конфликтом поведенческой адаптации». Наука. 318 (5852): 987–90. Bibcode:2007Наука ... 318..987М. Дои:10.1126 / science.1146384. PMID 17962523. S2CID 31089526.
- ^ а б c Mansouri, Farshad A .; Танака, Кейджи; Бакли, Марк Дж. (Февраль 2009 г.). «Поведенческая адаптация, вызванная конфликтом: ключ к исполнительным функциям префронтальной коры». Обзоры природы Неврология. 10 (2): 141–152. Дои:10.1038 / номер 2538. PMID 19153577. S2CID 15181627.
- ^ а б Кейн, Майкл Дж .; Энгл, Рэндалл В. (2002). «Роль префронтальной коры в емкости рабочей памяти, исполнительном внимании и общем подвижном интеллекте: перспектива индивидуальных различий». Психономический бюллетень и обзор. 9 (4): 637–71. Дои:10.3758 / BF03196323. PMID 12613671.
- ^ а б c d е Шакман, Александр; Брентон В. Макменамин; Джеффри С. Максвелл; Лоуренс Л. Грейшар; Ричард Дж. Дэвидсон (апрель 2009 г.). «Правая дорсолатеральная префронтальная кортикальная активность и поведенческое торможение». Психологическая наука. 20 (12): 1500–1506. Дои:10.1111 / j.1467-9280.2009.02476.x. ЧВК 2858783. PMID 19906125.
- ^ Van Den Bos, W .; Van Dijk, E .; Westenberg, M .; Rombouts, S.A.R.B .; Крон, Э. А. (2010). "Изменение мозга, изменение перспектив: нейрокогнитивное развитие взаимности". Психологическая наука. 22 (1): 60–70. Дои:10.1177/0956797610391102. PMID 21164174. S2CID 5026096.
- ^ Петрикан, Ралука; Шиммак, Ульрих (2008). «Роль дорсолатеральной префронтальной функции в приверженности отношениям». Журнал исследований личности. 42 (4): 1130–5. Дои:10.1016 / j.jrp.2008.03.001.
- ^ Ян, Ян; Паспалас, Константинос Д .; Джин, Лу Э .; Picciotto, Marina R .; Арнстен, Эми Ф. Т .; Ван, Мин (2013). «Никотиновые рецепторы α7 усиливают когнитивные цепи NMDA в дорсолатеральной префронтальной коре». Труды Национальной академии наук. 110 (29): 12078–83. Bibcode:2013ПНАС..11012078Y. Дои:10.1073 / pnas.1307849110. ЧВК 3718126. PMID 23818597.
- ^ Калликотт, Джозеф Х. (2000). "Психологическая дисфункция дорсолатеральной префронтальной коры при шизофрении повторно". Кора головного мозга (Нью-Йорк, Нью-Йорк: 1991). 10 (11): 1078–92. Дои:10.1093 / cercor / 10.11.1078. PMID 11053229. Получено 15 ноября 2013.
- ^ а б Кенигс, Майкл, Графманб, Джордан (2009). «Функциональная нейроанатомия депрессии: разные роли вентромедиальной и дорсолатеральной префронтальной коры». Поведенческие исследования мозга. 201 (2): 239–243. Дои:10.1016 / j.bbr.2009.03.004. ЧВК 2680780. PMID 19428640.
- ^ Ошибка цитирования: указанная ссылка Functional_Genomics была вызвана, но не определена
- ^ а б c d Цинь, Шаочжэн; Hermans, Erno J .; Van Marle, Hein J.F .; Ло, Цзин; Фернандес, Гильен (2009). «Острый психологический стресс снижает активность, связанную с рабочей памятью, в дорсолатеральной префронтальной коре». Биологическая психиатрия. 66 (1): 25–32. Дои:10.1016 / j.biopsych.2009.03.006. PMID 19403118. S2CID 22601360.
- ^ Cohen, H .; Каплан, З; Котлер, М; Куперман, я; Моиса, Р; Грисару, Н. (2004). «Повторяющаяся транскраниальная магнитная стимуляция правой дорсолатеральной префронтальной коры при посттравматическом стрессовом расстройстве: двойное слепое плацебо-контролируемое исследование». Американский журнал психиатрии. 161 (3): 515–24. Дои:10.1176 / appi.ajp.161.3.515. PMID 14992978.
- ^ а б c Говин, Джошуа Л., Макки, Скотт, Паулюс, Мартин П. (2013). «Измененная обработка рисков у потребителей психоактивных веществ: дисбаланс между болью и выгодами». Наркотическая и алкогольная зависимость. 132 (1–2): 13–21. Дои:10.1016 / j.drugalcdep.2013.03.019. ЧВК 3748224. PMID 23623507.
- ^ Лурия, изд. К.Х. Прибрам, А. (1973). Психофизиология лобных долей. Нью-Йорк: Academic Press. ISBN 978-0125643405.CS1 maint: дополнительный текст: список авторов (связь)
- ^ а б Абернати, Кеннет, Чендлер, Л. Джадсон, Wooward, Джон Дж. (2010). «Алкоголь и префронтальная кора». Функциональная пластичность и генетическая изменчивость: взгляд на нейробиологию алкоголизма. Int. Преподобный Neurobiol. Международный обзор нейробиологии. 91. С. 289–320. Дои:10.1016 / S0074-7742 (10) 91009-X. ISBN 9780123812766. ЧВК 3593065. PMID 20813246.