Эпихлоя - Epichloë
| Эпихлоя | |
|---|---|
 | |
| «Удушающая болезнь»: Эпихлоя тифина строма на мятлик | |
| Научная классификация | |
| Королевство: | |
| Разделение: | |
| Класс: | |
| Подкласс: | |
| Заказ: | |
| Семья: | |
| Племя: | |
| Род: | Эпихлоя |
| Типовой вид | |
| Эпихлоя тифина (Пт) Тюль. & C. Tul. | |
| Разнообразие | |
| 35 видов, см. Текст | |
Эпихлоя это род аскомицет грибы формирование эндофитный симбиоз с травы. Трава удушье является симптомом трав, вызванным некоторыми Эпихлоя виды, образующие спора -подшипниковые коврики (стромы ) на культиваторы и подавить развитие их растение-хозяин с соцветие. Однако большую часть своего жизненного цикла Эпихлоя растут в межклеточном пространстве стеблей, листьев, соцветий и семян травянистого растения, не вызывая симптомов болезни. Фактически, они обеспечивают хозяину несколько преимуществ, включая производство различных средств отпугивания травоядных животных. алкалоиды, повышенная стрессоустойчивость и стимуляция роста.
В семье Clavicipitaceae, Эпихлоя вложен в группу эндофитных и патогенные растения грибы, общий предок которых, вероятно, произошел от патогена животных. В род входят оба вида с половым размножением (телеоморфный ) этап и бесполый, анаморфный виды. Последние ранее размещались в род формы Неотифодиум но включен в Эпихлоя после молекулярная филогенетика показали, что бесполые и половые виды смешиваются в одном клады. Гибридное видообразование сыграл важную роль в эволюции рода.
Эпихлоя виды являются экологически значимыми из-за их воздействия на растения-хозяева. Было показано, что их присутствие изменяет состав растительные сообщества и пищевые полотна. Сорта трав, особенно овсяница высокая и райграс, с симбиотическим Эпихлоя штаммы эндофитов, коммерциализируются и используются для пастбище и дерн.
Таксономия
Элиас Фрис, в 1849 г. впервые определил Эпихлоя как подрод из Кордицепс.[1] Так как типовой вид, он обозначил Кордицепс тифина,[1] первоначально описанный Кристиан Хендрик Персун.[2] Братья Чарльз и Луи Рене Туласн затем поднял подрод до ранга рода в 1865 году.[3] Эпихлоя тифина останется единственным видом в этом роде до открытия грибковых эндофитов трав, вызывающих интоксикацию домашнего скота в 1970-х и 1980-х годах, что стимулировало описание новых видов.[4] Несколько видов из Африки и Азии, у которых развиваются стромы на травах, были выделены в отдельный род. Парепихлоя в 1998 г.[5]
Много Эпихлоя У видов есть формы, которые размножаются половым путем, и несколько чисто бесполых видов тесно связаны с ними. Эти анаморфы долго классифицировались отдельно: Morgan-Jones and Gams (1982) собрали их в секции (Альбо-ланоса) рода Акремониум.[6] В молекулярный филогенетический исследование, проведенное в 1996 году, Гленн и его коллеги обнаружили, что этот род полифилетический и предложил новый род Неотифодиум для анаморфных видов, относящихся к Эпихлоя.[7] Некоторые виды продолжали описываться в обоих родах до тех пор, пока Leuchtmann с коллегами (2014) не включили большую часть род формы Неотифодиум в Эпихлоя.[4] Филогенетические исследования показали, что оба рода смешаны, и номенклатурный код с 2011 г. требуется, чтобы одно название использовалось для всех стадий развития вида грибов. Только Neotyphodium starrii, неясного статуса, и N. chilense, который не связан, были исключены из Эпихлоя.[4]


Разновидность
По состоянию на 2020 год в роду насчитывается 35 признанных видов, из которых 3 подвид и 6 разновидности описано. 14 видов, 3 подвида и 5 разновидностей гаплоидны. 21 вид и 1 сорт - гибриды (аллополиплоиды ). Некоторые таксоны известны только как анаморфные (бесполые) формы, большинство из которых ранее были классифицированы как Неотифодиум.[4]
| Гаплоидные таксоны | Известное распространение | Половое размножение | Вертикальная передача | Известный диапазон хостов | Ссылка на описание вида |
|---|---|---|---|---|---|
| Epichloë amarillans Дж. Ф. Уайт | Северная Америка | Наблюдаемый | Подарок | Agrostis hyemalis, Agrostis perennans, Calamagrostis canadensis, Elymus virginicus, Sphenopholis nitida, Sphenopholis obtusata, Sphenopholis × pallens, Ammophila breviligulata | Белый, Джеймс Ф. (1994). «Сообщества эндофитов-хозяев злаков. XX. Структурные и репродуктивные исследования Epichloë amarillans sp. ноя и сравнения с E. typhina". Микология. 86 (4): 571–580. Дои:10.1080/00275514.1994.12026452. ISSN 0027-5514. |
| Epichloë aotearoae (C.D. Moon, C.O. Miles & Schardl) Leuchtm. И Schardl | Новая Зеландия, Австралия | Не наблюдается | Подарок | Echinopogon ovatus | Луна, Кристина Д .; Майлз, Кристофер О .; Ярфорс, Улла; Шардл, Кристофер Л. (2002). "Эволюционное происхождение трех новых Неотифодиум виды эндофитов трав, произрастающих в Южном полушарии ". Микология. 94 (4): 694–711. Дои:10.1080/15572536.2003.11833197. ISSN 0027-5514. PMID 21156542. S2CID 12259278. |
| Эпихлоя беконий Дж. Ф. Уайт | Европа | Наблюдаемый | Отсутствует | Agrostis capillaris, Agrostis stolonifera, Calamagrostis villosa, Calamagrostis varia, Calamagrostis purpurea | Белый, Джеймс Ф. (1993). "Сообщества эндофитов-хозяев в травах. XIX. Систематическое изучение некоторых симпатрических видов растений". Эпихлоя в Англии". Микология. 85 (3): 444–455. Дои:10.1080/00275514.1993.12026295. ISSN 0027-5514. |
| Epichloë brachyelytri Schardl & Leuchtm. | Северная Америка | Наблюдаемый | Подарок | Brachyelytrum erectum | Schardl, Christopher L .; Leuchtmann, Адриан (1999). "Три новых вида Эпихлоя симбиотик с североамериканскими травами ". Микология. 91 (1): 95–107. Дои:10.1080/00275514.1999.12060996. ISSN 0027-5514. |
| Эпихлоя бромикола Leuchtm. И Schardl | Европа, Азия | Наблюдается на Bromus erectus, Elymus repens и Elymus tsukushiensis | Присутствует в Bromus Benekenii, Bromus ramosus и Hordelymus europaeus, Hordeum brevisubulatum, Leymus chinensis и Elymus tsukushiensis; отсутствует в Bromus erectus и Elymus repens | Европа: Bromus Benekenii, Бром прямоходящий, Bromus ramosus, Elymus repens, Hordelymus europaeus, Hordeum brevisubulatum. Азия: Leymus chinensis, Elymus tsukushiensis | Leuchtmann, Адриан; Шардл, Кристофер Л. (1998). "Совместимость спаривания и филогенетические отношения между двумя новыми видами Эпихлоя и другие родственные европейские виды ". Микологические исследования. 102 (10): 1169–1182. Дои:10.1017 / S0953756298006236. ISSN 0953-7562. |
| Эпихлоя Элими Schardl & Leuchtm. | Северная Америка | Наблюдаемый | Подарок | Bromus kalmii, Элим виды (включая Elymus hystrix ) | Schardl, Christopher L .; Leuchtmann, Адриан (1999). "Три новых вида Эпихлоя симбиотик с североамериканскими травами ". Микология. 91 (1): 95–107. Дои:10.1080/00275514.1999.12060996. ISSN 0027-5514. |
| Epichloë festucae Leuchtm., Schardl & M.R. Siegel | Европа, Азия, Северная Америка | Наблюдаемый | Подарок | Festuca виды, Koeleria виды, Щедонор виды | Лойхтманн, Адриан; Schardl, Christopher L .; Сигел, Малкольм Р. (1994). «Сексуальная совместимость и таксономия нового вида Эпихлоя симбиотик с мелкими овсяницами ». Микология. 86 (6): 802–812. Дои:10.1080/00275514.1994.12026487. ISSN 0027-5514. |
| Epichloë festucae var. лолии (Защелка, M.J. Chr. & Samuels) К.У. Бэкон и Шардл | Европа, Азия, Северная Африка, завезен в Новой Зеландии, Австралии и других странах. | Не наблюдается | Подарок | Lolium perenne subsp. Perenne | Защелка, G.C.M .; Christensen, M.J .; Сэмюэлс, Г.Дж. (1984). «Пять эндофитов Лолиум и Festuca в Новой Зеландии ». Микотаксон. 20: 535–550. |
| Epichloë gansuensis (Си Джей Ли и Нэн) Шардл | Азия | Не наблюдается | Подарок | Achnatherum inebrians, Achnatherum sibiricum, Achnatherum pekinense | Li, C.J .; Nan, Z.B .; Paul, V.H .; Dapprich, P.D .; Лю, Ю. (2004). "Новый Неотифодиум вид-симбиотик с пьяной конской травой (Achnatherum inebrians) в Китае". Микотаксон. 90: 141–147. |
| Epichloë gansuensis var. алкоголики (C.D. Moon & Schardl) Шардл | Азия | Не наблюдается | Подарок | Achnatherum inebrians | Луна, Кристина Д .; Гийомен, Жан-Жак; Равель, Екатерина; Ли, Чунджи; Craven, Kelly D .; Шардл, Кристофер Л. (2007). "Новый Неотифодиум виды эндофитов травяных триб Stipeae и Meliceae ». Микология. 99 (6): 895–905. Дои:10.1080/15572536.2007.11832521. ISSN 0027-5514. PMID 18333513. S2CID 19953493. |
| Epichloë glyceriae Schardl & Leuchtm. | Северная Америка | Наблюдаемый | Отсутствует | Глицерия виды | Schardl, Christopher L .; Leuchtmann, Адриан (1999). "Три новых вида Эпихлоя симбиотик с североамериканскими травами ". Микология. 91 (1): 95–107. Дои:10.1080/00275514.1999.12060996. ISSN 0027-5514. |
| Эпихлоя моллис (Morgan-Jones & W. Gams) Leuchtm. И Schardl | Европа | Наблюдаемый | Подарок | Holcus mollis | Morgan-Jones, G .; Гамс, В. (1982). «Заметки о гифомицетах. XLI. Эндофит Festuca arundinacea и анаморф Эпихлоя тифина, новые таксоны в одном из двух новых разрезов Акремониум". Микотаксон. 15: 311–318. ISSN 0093-4666. |
| Эпихлоя сибирская (X. Zhang, Y.B. Gao) Тадыч | Азия | Не наблюдается | Подарок | Achnatherum sibiricum | Чжан, Синь; Рен, Ань-Чжи; Вэй, Ю-Кун; Линь, Фэн; Ли, Чуан; Лю, Чжи-Цзянь; Гао, Юй-Бао (2009). «Таксономия, разнообразие и происхождение симбиотических эндофитов Achnatherum sibiricum в степи Внутренней Монголии Китая ». Письма о микробиологии FEMS. 301 (1): 12–20. Дои:10.1111 / j.1574-6968.2009.01789.x. ISSN 0378-1097. PMID 19863662. |
| Эпихлоя строматолонга (Y.L. Ji, L.H. Zhan & Z.W. Wang) Leuchtm. | Азия | Не наблюдается | Подарок | Calamagrostis epigejos | Цзи, Янь-лин; Чжань, Ли-хуэй; Канг, Ян; Сунь, Сян-хуэй; Ю, Хань-шоу; Ван, Чжи-вэй (2009). "Новый продуцент стромы Неотифодиум вид-симбиотик с клоновой травой Calamagrostis epigeios (Л.) Рот. выращен в Китае ». Микология. 101 (2): 200–205. Дои:10.3852/08-044. ISSN 0027-5514. PMID 19397192. S2CID 32237846. |
| Epichloë sylvatica Leuchtm. И Schardl | Европа, Азия | Наблюдаемый | Подарок | Brachypodium sylvaticum, Hordelymus europaeus | Лойхтманн, Адриан; Шардл, Кристофер Л. (1998). "Совместимость спаривания и филогенетические отношения между двумя новыми видами Эпихлоя и другие родственные европейские виды ". Микологические исследования. 102 (10): 1169–1182. Дои:10.1017 / S0953756298006236. ISSN 0953-7562. |
| Epichloë sylvatica subsp. pollinensis Leuchtm. И М. Оберхофер | Европа | Наблюдаемый | Подарок | Hordelymus europaeus | Лойхтманн, Адриан; Оберхофер, Мартина (2013). "The Эпихлоя эндофиты, связанные с лесной травой Hordelymus europaeus в том числе четыре новых таксона ". Микология. 105 (5): 1315–1324. Дои:10.3852/12-400. ISSN 0027-5514. PMID 23921239. S2CID 21024362. |
| Эпихлоя тифина (Чел.) Tul. & C. Tul. | Европа, завезен в Северной Америке и других странах. | Наблюдаемый | Присутствует в Puccinellia distans; отсутствует в других хостах | Anthoxanthum odoratum, Brachypodium phoenicoides, Brachypodium pinnatum, Dactylis glomerata, Lolium perenne, Milium effusum, Phleum pratense, Мятлик тривиальный, Мятлик сильвикола, Puccinellia distans | Туласне, Л.Р .; Туласне, К. (1865). "Nectriei-Phacidiei-Pezizei". Selecta Fungorum Carpologia. Империал: Париж. 3: 24. |
| Эпихлоя тифина subsp. Кларки (Дж. Ф. Уайт) Leuchtm. И Schardl | Европа | Наблюдаемый | Отсутствует | Holcus lanatus | Белый, Джеймс Ф. (1993). "Сообщества эндофитов-хозяев в травах. XIX. Систематическое изучение некоторых симпатрических видов растений". Эпихлоя в Англии". Микология. 85 (3): 444–455. Дои:10.1080/00275514.1993.12026295. ISSN 0027-5514. |
| Эпихлоя тифина subsp. poae (Тадыч, К. В. Амвросий, Ф. К. Белэнджер, Дж. Ф. Уайт) Тадыч | Европа, Северная Америка | Наблюдается на Мятлик неморалис и Мятлик луговой | Присутствует в Мятлик неморалис, Poa secunda subsp. Juncifolia; отсутствует в Мятлик луговой | Европа: Мятлик неморалис, Мятлик луговой. Северная Америка: Poa secunda subsp. Juncifolia, Мятлик лесной | Тадыч, Мариуш; Амвросий, Карен В .; Bergen, Marshall S .; Белэнджер, Фейт К .; Белый, Джеймс Ф. (2012). "Таксономическое размещение Epichloë poae sp. ноя и горизонтальное распространение на сеянцы через конидии ". Грибковое разнообразие. 54 (1): 117–131. Дои:10.1007 / s13225-012-0170-0. ISSN 1560-2745. S2CID 17813728. |
| Эпихлоя тифина subsp. poae var. Аоникенкана Янноне и Шардль | Аргентина (Санта-Крус) | Не наблюдается | Подарок | Bromus setifolius | Mc Cargo, Patricia D .; Iannone, Leopoldo J .; Виньяле, Мария Виктория; Schardl, Christopher L .; Росси, Мария Сусана (2017). "Видовое разнообразие Эпихлоя симбиотик с двумя травами из южной аргентинской Патагонии ". Микология. 106 (2): 339–352. Дои:10.3852/106.2.339. ISSN 0027-5514. PMID 24782501. S2CID 44547876. |
| Эпихлоя тифина subsp. poae var. canariensis (C.D. Moon, B. Scott и M.J. Chr.) Leuchtm. | Канарские острова | Не наблюдается | Подарок | Лолиум Эдвардии | Луна, Кристина Д .; Скотт, Барри; Schardl, Christopher L .; Кристенсен, Майкл Дж. (2000). "Эволюционное происхождение Эпихлоя эндофиты райграса однолетнего ». Микология. 92 (6): 1103–1118. Дои:10.1080/00275514.2000.12061258. ISSN 0027-5514. S2CID 218589443. |
| Эпихлоя тифина subsp. poae var. Уэрфана (Дж. Ф. Уайт, Г. Т. Коул и Морган-Джонс) Tadych & Leuchtm. | Северная Америка | Не наблюдается | Подарок | Festuca arizonica | Гленн, Энтони Э .; Бэкон, Чарльз У .; Прайс, Роберт; Ханлин, Ричард Т. (1996). «Молекулярная филогения Акремониум и его таксономические последствия ". Микология. 88 (3): 369–383. Дои:10.1080/00275514.1996.12026664. ISSN 0027-5514. |
| Гибридные таксоны | Виды-прародители | Известное распространение | Половое размножение | Вертикальная передача | Известный диапазон хостов | Ссылка на описание вида |
|---|---|---|---|---|---|---|
| Эпихлоя также Шиманович Т., Янг, Н.Д. Чарльтон и С.Х. Фаэт | Epichloë amarillans × Эпихлоя тифина subsp. poae | Северная Америка | Не наблюдается | Подарок | Poa alsodes | Шиманович, Татьяна; Чарльтон, Никки Д .; Musso, Ashleigh M .; Ширер, Джонатан; Чех, Надя Б .; Faeth, Stanley H .; Янг, Кэролайн А. (2017). «Межвидовой и внутривидовой гибрид. Эпихлоя вид-симбиотик с местной травой Северной Америки Poa alsodes" (PDF). Микология. 109 (3): 459–474. Дои:10.1080/00275514.2017.1340779. ISSN 0027-5514. PMID 28723242. S2CID 25290203. |
| Эпихлоя австралийская (C.D. Moon & Schardl) Leuchtm. И Schardl | Epichloë festucae × Эпихлоя тифина сложный (от Мятлик луговой ) | Австралия | Не наблюдается | Подарок | Echinopogon ovatus | Луна, Кристина Д .; Майлз, Кристофер О .; Ярфорс, Улла; Шардл, Кристофер Л. (2017). "Эволюционное происхождение трех новых Неотифодиум виды эндофитов трав, произрастающих в Южном полушарии ". Микология. 94 (4): 694–711. Дои:10.1080/15572536.2003.11833197. ISSN 0027-5514. PMID 21156542. S2CID 12259278. |
| Epichloë cabralii Янноне, М.С. Росси и Шардл | Epichloë amarillans × Эпихлоя тифина сложный (от Мятлик неморалис ) | Аргентина (Санта-Крус, Огненная Земля) | Не наблюдается | Подарок | Phleum alpinum | Mc Cargo, Patricia D .; Iannone, Leopoldo J .; Виньяле, Мария Виктория; Schardl, Christopher L .; Росси, Мария Сусана (2017). "Видовое разнообразие Эпихлоя симбиотик с двумя травами из южной аргентинской Патагонии ". Микология. 106 (2): 339–352. Дои:10.3852/106.2.339. ISSN 0027-5514. PMID 24782501. S2CID 44547876. |
| Epichloë canadensis Н.Д. Чарльтон и К.А. Молодой | Epichloë amarillans × Эпихлоя Элими | Северная Америка | Не наблюдается | Подарок | Elymus canadensis | Charlton, N.D .; Craven, K. D .; Mittal, S .; Хопкинс, А. А .; Янг, К. А. (2012). "Epichloë canadensis, новый межвидовой эпихлоидный гибридный симбиотик с канадской дикой рожью (Elymus canadensis)". Микология. 104 (5): 1187–1199. Дои:10.3852/11-403. ISSN 0027-5514. PMID 22675049. S2CID 43674700. |
| Epichloë Chisosa (Дж. Ф. Уайт и Морган-Джонс) Шардл | Epichloë amarillans × Эпихлоя бромикола × Эпихлоя тифина сложный (от Мятлик луговой ) | Северная Америка | Не наблюдается | Подарок | Achnatherum eminens | Гленн, Энтони Э .; Бэкон, Чарльз У .; Прайс, Роберт; Ханлин, Ричард Т. (2018). «Молекулярная филогения Акремониум и его таксономические последствия ". Микология. 88 (3): 369–383. Дои:10.1080/00275514.1996.12026664. ISSN 0027-5514. |
| Epichloë coenophiala (Морган-Джонс и В. Гэмс) К.В. Бэкон и Шардл | Эпихлоя беконий (Лолиум ассоциированная клада) × Epichloë festucae × Эпихлоя тифина сложный (из Мятлик неморалис ) | Европа, Северная Африка, завезен в Северной Америке и других местах. | Не наблюдается | Подарок | Schedonorus arundinaceus [синонимы: Festuca arundinacea, Лолиум арундинацеум] | Morgan-Jones, G .; Гамс, В. (1982). «Заметки о гифомицетах. XLI. Эндофит Festuca arundinacea и анаморф Эпихлоя тифина, новые таксоны в одном из двух новых разрезов Акремониум". Микотаксон. 15: 311–318. ISSN 0093-4666. |
| Эпихлоя Даника Leuchtm. И М. Оберхофер | Эпихлоя бромикола × Epichloë sylvatica | Европа | Не наблюдается | Подарок | Hordelymus europaeus | Лойхтманн, Адриан; Оберхофер, Мартина (2017). "The Эпихлоя эндофиты, связанные с лесной травой Hordelymus europaeus в том числе четыре новых таксона ". Микология. 105 (5): 1315–1324. Дои:10.3852/12-400. ISSN 0027-5514. PMID 23921239. S2CID 21024362. |
| Epichloë disjuncta Leuchtm. И М. Оберхофер | 'Эпихлоя тифина комплекс × второй неизвестный предок | Европа | Не наблюдается | Подарок | Hordelymus europaeus | Лойхтманн, Адриан; Оберхофер, Мартина (2017). "The Эпихлоя эндофиты, связанные с лесной травой Hordelymus europaeus в том числе четыре новых таксона ". Микология. 105 (5): 1315–1324. Дои:10.3852/12-400. ISSN 0027-5514. PMID 23921239. S2CID 21024362. |
| Эпихлоя фанки (К.Д. Крейвен и Шардл) Дж. Ф. Уайт | Эпихлоя Элими × Epichloë festucae | Северная Америка | Не наблюдается | Подарок | Achnatherum robustum | Луна, Кристина Д .; Гийомен, Жан-Жак; Равель, Екатерина; Ли, Чунджи; Craven, Kelly D .; Шардл, Кристофер Л. (2017). "Новый Неотифодиум виды эндофитов травяных триб Stipeae и Meliceae ». Микология. 99 (6): 895–905. Дои:10.1080/15572536.2007.11832521. ISSN 0027-5514. PMID 18333513. S2CID 19953493. |
| Epichloë Guerinii (Guillaumin, Ravel и C.D. Moon) Leuchtm. И Schardl | Epichloë gansuensis × Эпихлоя тифина сложный | Европа | Не наблюдается | Подарок | Melica ciliata, Melica transsilvanica | Луна, Кристина Д .; Гийомен, Жан-Жак; Равель, Екатерина; Ли, Чунджи; Craven, Kelly D .; Шардл, Кристофер Л. (2017). "Новый Неотифодиум виды эндофитов травяных триб Stipeae и Meliceae ». Микология. 99 (6): 895–905. Дои:10.1080/15572536.2007.11832521. ISSN 0027-5514. PMID 18333513. S2CID 19953493. |
| Эпихлоя горделыми Leuchtm. И М. Оберхофер | Эпихлоя бромикола × Эпихлоя тифина сложный | Европа | Не наблюдается | Подарок | Hordelymus europaeus | Leuchtmann, Адриан; Оберхофер, Мартина (2017). "The Эпихлоя эндофиты, связанные с лесной травой Hordelymus europaeus в том числе четыре новых таксона ". Микология. 105 (5): 1315–1324. Дои:10.3852/12-400. ISSN 0027-5514. PMID 23921239. S2CID 21024362. |
| Эпихлоя гибридная М.П. Кокс и М.А.Кэмпелл | Epichloë festucae var. лолии × Эпихлоя тифина | Европа | Не наблюдается | Подарок | Lolium perenne | Кэмпбелл, Мэтью А .; Tapper, Brian A .; Симпсон, Уэйн Р .; Джонсон, Ричард Д .; Мейс, Уэйд; Рам, Арвина; Лукито, Йонатан; Дюпон, Пьер-Ив; Джонсон, Линда Дж .; Скотт, Д. Барри; Ganley, Austen R.D .; Кокс, Мюррей П. (2017). "Эпихлоя гибридная, sp. nov., новая модельная система для исследования аллополиплоидии грибов ». Микология. 109 (5): 1–15. Дои:10.1080/00275514.2017.1406174. ISSN 0027-5514. PMID 29370579. |
| Epichloë liyangensis Z.W. Ван, Ю. Кан и Х. Мяо | Эпихлоя бромикола × Эпихлоя тифина сложный (от Мятлик неморалис ) | Азия | Наблюдаемый | Подарок | Мятлик луговой subsp. луговой | Ян, Канг; Янлин, Цзи; Кунран, Чжу; Хуэй, Ван; Хуйминь, Мяо; Чживэй, Ван (2017). "Новый Эпихлоя виды с межвидовым гибридным происхождением от Мятлик луговой ssp. луговой в Лияне, Китай ". Микология. 103 (6): 1341–1350. Дои:10.3852/10-352. ISSN 0027-5514. PMID 21659456. S2CID 22672333. |
| Epichloë melicicola (C.D. Moon & Schardl) Шардл | Epichloë aotearoae × Epichloë festucae | Южная Африка | Не наблюдается | Подарок | Melica racemosa, Melica decumbens | Луна, Кристина Д .; Майлз, Кристофер О .; Ярфорс, Улла; Шардл, Кристофер Л. (2017). "Эволюционное происхождение трех новых Неотифодиум виды эндофитов трав, произрастающих в Южном полушарии ". Микология. 94 (4): 694–711. Дои:10.1080/15572536.2003.11833197. ISSN 0027-5514. PMID 21156542. S2CID 12259278. |
| Epichloë novae-zelandiae Leuchtm. И А.В. Стюарт | Epichloë amarillans × Эпихлоя бромикола × Эпихлоя тифина subsp. poae | Новая Зеландия | Не наблюдается | Подарок | Poa Matthewsii | Leuchtmann, Адриан; Янг, Кэролайн А .; Стюарт, Алан В .; Симпсон, Уэйн Р .; Хьюм, Дэвид Э .; Скотт, Барри (2019). "Epichloë novae-zelandiae, новый эндофит из эндемичной новозеландской травы Poa Matthewsii". Новозеландский ботанический журнал. 57 (4): 271–288. Дои:10.1080 / 0028825X.2019.1651344. ISSN 0028-825X. S2CID 202012350. |
| Epichloë occultans (C.D. Moon, B. Scott и M.J. Chr.) Шардл | Эпихлоя беконий (Лолиум ассоциированная клада) × Эпихлоя бромикола | Европа, Северная Африка, завезен в Новую Зеландию и другие страны. | Не наблюдается | Подарок | Лолиум мультифлорум, Лолиум ригидум u.a. | Луна, Кристина Д .; Скотт, Барри; Schardl, Christopher L .; Кристенсен, Майкл Дж. (2019). "Эволюционное происхождение Эпихлоя эндофиты райграса однолетнего ». Микология. 92 (6): 1103–1118. Дои:10.1080/00275514.2000.12061258. ISSN 0027-5514. S2CID 218589443. |
| Эпихлоя пампеана (Янноне и Кабрал) Ианноне и Шардл | Epichloë festucae × Эпихлоя тифина сложный (от Мятлик неморалис ) | Южная Америка | Не наблюдается | Подарок | Bromus auleticus | Янноне, Леопольдо Хавьер; Кабрал, Даниэль; Шардл, Кристофер Льюис; Росси, Мария Сусана (2017). «Филогенетическая дивергенция, морфологические и физиологические различия отличают новый Неотифодиум виды эндофитов в траве Bromus auleticus из Южной Америки ». Микология. 101 (3): 340–351. Дои:10.3852/08-156. ISSN 0027-5514. PMID 19537207. S2CID 24799520. |
| Эпихлоя Шардлии (Ghimire, Rudgers & K.D. Craven) Leuchtm. | Эпихлоя тифина сложный (подп. poae × подп. poae) | Северная Америка | Не наблюдается | Подарок | Cinna arundinacea | Ghimire, Sita R .; Rudgers, Jennifer A .; Чарльтон, Никки Д .; Янг, Кэролайн; Крейвен, Келли Д. (2017). «Распространенность внутривидового Неотифодиум гибрид в природных популяциях камыша крепкого (Cinna arundinacea L.) из восточной части Северной Америки ». Микология. 103 (1): 75–84. Дои:10.3852/10-154. ISSN 0027-5514. PMID 20943524. S2CID 13556418. |
| Эпихлоя Шардлии var. Pennsylvanica Шиманович Т., Янг, Н.Д. Чарльтон и С.Х. Фаэт | Эпихлоя тифина сложный (subsp. poae × подп. poae) | Северная Америка | Не наблюдается | Подарок | Poa alsodes | Шиманович, Татьяна; Чарльтон, Никки Д .; Musso, Ashleigh M .; Ширер, Джонатан; Чех, Надя Б .; Faeth, Stanley H .; Янг, Кэролайн А. (2017). «Межвидовой и внутривидовой гибрид. Эпихлоя вид-симбиотик с местной травой Северной Америки Poa alsodes" (PDF). Микология. 109 (3): 459–474. Дои:10.1080/00275514.2017.1340779. ISSN 0027-5514. PMID 28723242. S2CID 25290203. |
| Epichloë siegelii (K.D. Craven, Leuchtm. & Schardl) Leuchtm. И Schardl | Эпихлоя бромикола × Epichloë festucae | Европа | Не наблюдается | Подарок | Schedonorus pratensis (синонимы: Festuca pratensis, Лолиум пратенс) | Craven, K.D .; Бланкеншип, J.D .; Leuchtmann, A .; Hinight, K .; Шардл, К. (2001). «Гибридные грибковые эндофиты - симбиотические с травой. Лолиум пратенс". Sydowia. 53: 44–73. |
| Эпихлоя Синица (Z.W. Wang, Y.L. Ji & Y. Kang) Leuchtm. | Эпихлоя бромикола × Эпихлоя тифина сложный | Азия | Не наблюдается | Подарок | Roegneria виды | Ян, Канг; Янлин, Цзи; Сянхуи, Солнце; Лихуэй, Чжань; Вэй, Ли; Ханьшоу, Ю; Чживэй, Ван (2017). "Таксономия Неотифодиум эндофиты китайского происхождения Roegneria растения ». Микология. 101 (2): 211–219. Дои:10.3852/08-018. ISSN 0027-5514. PMID 19397194. S2CID 6307330. |
| Epichloë sinofestucae (Y.G. Chen, Y.L. Ji & Z.W. Wang) Leuchtm. | Эпихлоя бромикола × Эпихлоя тифина сложный | Азия | Не наблюдается | Подарок | Festuca parvigluma | Чен, Юн-ган; Цзи, Янь-лин; Ю, Хань-шоу; Ван, Чжи-вэй (2017). "Новый Неотифодиум виды из Festuca parvigluma Steud. выращен в Китае ». Микология. 101 (5): 681–685. Дои:10.3852/08-181. ISSN 0027-5514. PMID 19750947. S2CID 27915317. |
| Epichloë tembladerae (Кабрал и Дж. Ф. Уайт) Ианноне и Шардл | Epichloë festucae × Эпихлоя тифина сложный (от Мятлик неморалис ) | Северная Америка | Не наблюдается | Подарок | Северная Америка: Festuca arizonica. Южная Америка: Bromus auleticus, Bromus setifolius, Festuca argentina, Festuca hieronymi, Festuca magellanica, Festuca superba, Melica stuckertii, Phleum alpinum, Phleum commutatum, Poa Huecu, Мятлик ригиднолистный | Кабрал, Даниэль; Cafaro, Matías J .; Saidman, B .; Луго, М .; Редди, Понака В .; Белый, Джеймс Ф. (2019). «Свидетельства, подтверждающие появление нового вида эндофитов в некоторых травах Южной Америки». Микология. 91 (2): 315–325. Дои:10.1080/00275514.1999.12061021. ISSN 0027-5514. |
| Epichloë uncinata (В. Гамс, Петрини и Д. Шмидт) Leuchtm. И Schardl | Эпихлоя бромикола × Эпихлоя тифина сложный | Европа | Не наблюдается | Подарок | Schedonorus pratensis (синонимы: Festuca pratensis, Лолиум пратенс) | Gams, W .; Петрини, О. Дж .; Шмидт, Д. (1990). "Acremonium uncinatum, новый эндофит в Festuca pratensis". Микотаксон. 37: 67–71.A1: G25 |
Жизненный цикл и рост
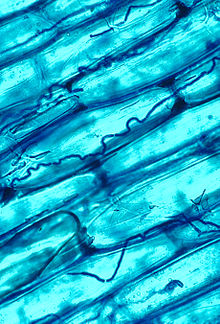
Эпихлоя виды специализируются на формировании и поддержании системных, конститутивных (долгосрочных) симбиозы с растениями, часто с ограниченным заболеванием или без него.[8] Наиболее изученные из этих симбионтов связаны с травы и осока, при котором они заражают листья и другие воздушные ткани, разрастаясь между клетками растений (эндофитный рост) или на поверхности над или под кутикулой (эпифитный рост).Отдельное инфицированное растение обычно несет только один генетический индивидуальный ключицкий симбионт, поэтому система растение-гриб составляет генетическую единицу, называемую симбиотумом (мн. Симбиота).
Симптомы и признаки грибковой инфекции, если они вообще проявляются, возникают только на определенной ткани или участке хозяина. культиватор, где появляется грибковая строма или склероций. Строма (pl. Stromata) представляет собой мицелиальную подушку, которая в первую очередь дает начало бесполым спорам (конидии ), затем к половым плодовым телам (аскокарпы; перитеция). Склероции представляют собой твердые покоящиеся структуры, которые позже (после инкубации на земле) прорастают, образуя строматы прилистников. В зависимости от вида гриба тканями хозяина, на которых образуются стромы или склероции, могут быть молодые соцветия и окружающие их листья, отдельные соцветия, узлы или небольшие сегменты листьев. Молодые стромы прозрачные (бесцветные), по мере созревания становятся темно-серыми, черными или желто-оранжевыми. Зрелые стромы выделяют споры мейотического происхождения (аскоспоры ), которые выбрасываются в атмосферу и вызывают новые инфекции растений (горизонтальная передача ). В некоторых случаях не образуется ни строма, ни склероций, но грибок заражает семена, продуцируемые инфицированным растением, и тем самым передается вертикально следующему поколению хозяев. Наиболее Эпихлоя виды и все бесполые виды могут вертикально передавать.
Таксономическая дихотомия особенно интересна для этой группы симбионтов, поскольку вегетативное размножение грибных мицелий происходит путем вертикальной передачи, то есть роста грибов во вновь развивающихся побегах-хозяевах (= отдельных травяных растениях). Важно отметить, что многие Эпихлоя виды заражают новые травяные растения исключительно путем прорастания семян своих травяных хозяев и заражения растущих саженцев.[9][10] Проявление сексуального состояния, которое происходит только в Эпихлоя вид - вызывает "удушье", состояние, при котором трава соцветия охвачены быстрым грибковым ростом, образующим строму. Строма гриба подавляет производство семян хозяина и достигает высшей точки в выбросе мейоспор (аскоспоры ), которые опосредуют горизонтальную (заразную) передачу грибка новым растениям.[9] Таким образом, два режима передачи исключают друг друга, хотя во многих травяныхЭпихлоя Symbiota гриб фактически демонстрирует оба способа передачи одновременно, заглушая некоторые побеги и передавая семена, полученные от незащищенных побегов.
В то же время облигатные симбионты в природе большинство эпихлоев легко культивируются в лаборатория на средства массовой информации такие как картофельный агар с декстрозой или минимальный солевой бульон с добавлением тиамина, сахаров или сахарных спиртов и органического азота или аммония.[11]
Эпихлоя виды обычно распространяются мухи рода Ботанофила. Мухи откладывают яйца в растущих тканях грибов, и личинки питаются ими.[12]

Эволюция
Эпихлоэ имеет ряд центральных особенностей, которые предполагают очень сильную и древнюю связь с их травяными хозяевами. Симбиоз, кажется, существовал уже во время ранней травы. эволюция что породило сегодняшний тупой травы. Это предлагается филогенетический исследования, указывающие на преобладание содивергенции Эпихлоя виды с травяными хозяевами, которые они населяют.[13] Рост грибного симбионта очень жестко регулируется в его траве-хозяине, на что указывает в основном неразветвленный мицелиальный морфология и замечательная синхронность травяного листа и гиф распространение грибка;[14][15] последнее, по-видимому, происходит через механизм, который включает вызванное растяжением или вставной удлинение гиф эндофита - процесс, который до сих пор не обнаружен ни у одного другого вида грибов, что указывает на специализированную адаптацию гриба к динамической среде роста внутри его хозяина.[16] Комплекс НАДФН оксидаза на основе ферментов ROS -генерирующая система в Эпихлоя вид незаменим для поддержания этой синхронности роста. Таким образом, было продемонстрировано, что удаление генов, кодирующих эти ферменты в Epichloë festucae вызывает серьезное нарушение роста грибков в тканях травы и даже гибель травяных растений.[17][18]
Молекулярно-филогенетические данные демонстрируют, что бесполое Эпихлоя виды происходят либо от полового Эпихлоя виды, или чаще, являются гибридами двух или более предков Эпихлоя виды.[19][20]
Биоактивные соединения

Много Эпихлоя эндофиты производят широкий спектр натуральный продукт соединения с биологической активностью против широкого круга травоядных.[21][22] Назначение этих соединений - как токсичность или кормовая защита от насекомых и млекопитающих травоядные животные.[23] Ergoline алкалоиды (алкалоиды спорыньи, названные в честь спорынья грибок Claviceps purpurea, близкий родственник epichloae) характеризуются кольцевой системой, производной от 4-пренил триптофан.[24] Одним из наиболее распространенных алкалоидов спорыньи в эпихлоэсимбиотических травах является эрговалин, содержащий эрголин. часть прикреплен к бициклическому трипептид содержащие аминокислоты L-пролин, L-аланин, и L-валин. Ключевые гены и ферменты биосинтеза алкалоидов спорыньи были идентифицированы в эпихлоях и включают: dmaW, кодировка диметилаллил-триптофансинтаза и lpsA, а нерибосомальная пептидная синтетаза.[24]
Другая группа эпихлоалкалоидов - это индол -дитерпеноиды, такие как лолитрем B, которые производятся в результате активности нескольких ферментов, в том числе пренилтрансферазы и различные монооксигеназы.[25] И эрголин, и индол-дитерпеноидные алкалоиды имеют биологическая активность против травоядных млекопитающих, а также против некоторых насекомых.[21] Перамин - это алкалоид пирролопиразина, который, как полагают, биосинтезируется из гуанидиний -группа-содержащая аминокислота L-аргинин, и пирролидин-5-карбоксилат, предшественник L-пролин,[26] и является средством, отпугивающим насекомых. В алкалоиды лолина[27] представляют собой 1-аминопирролизидины с атомом кислорода, связывающим мостиковые атомы углерода 2 и 7, и биосинтезируются из аминокислот L-пролин и L-гомосерин.[28] Лолинии имеют инсектицидный и деятельность по отпугиванию насекомых, сопоставимая с никотин.[27] Накопление лолина сильно индуцируется в молодых растущих тканях.[29] или повреждением симбиотума растение-гриб.[30] Многие, но не все, эпихлои производят до трех классов этих алкалоидов в различных комбинациях и количествах.[21] Недавно было показано, что Epichloë uncinata инфекция и содержание лолина позволяют × Фестулолиум защита трав от черного жука (Heteronychus arator ).[31]
Многие виды в Эпихлоя производят биологически активные алкалоиды, такие как алкалоиды спорыньи, индол-дитерпеноиды (например, лолитрем B ), алкалоиды лолина, и необычный гуанидиний алкалоид, перамин.[21]
Экология
Воздействие на траву
Было предложено, чтобы вертикально передаваемый симбионты должен развиваться, чтобы быть мутуалисты поскольку их репродуктивная фитнес тесно связано с хозяевами.[32] Фактически, некоторые положительные эффекты epichloae на их растения-хозяева включают ускоренный рост, засухоустойчивость и травоядное животное и возбудитель сопротивление.[9][33] Устойчивость к травоядным животным приписывают алкалоиды производятся симбиотическими эпихлоями.[21] Хотя симбиоз травы и эпихлои широко известен как мутуалистический во многих диких и культурных травах взаимодействия могут быть очень разнообразными, а иногда и антагонистическими, особенно в условиях бедности питательными веществами почвы.[34]
Динамика экосистемы
Из-за относительно большого числа видов трав, укрывающих эпихлоэ, и разнообразия окружающей среды, в которой они встречаются, механизмы, лежащие в основе благоприятных или антагонистических результатов симбиоза эпихлоэ и травы, трудно определить в естественных, а также сельскохозяйственных средах.[9][35] Некоторые исследования предполагают связь между выпасом травоядных животных и усилением заражения эпихлоем трав, которыми они питаются.[36][37] тогда как другие указывают на сложное взаимодействие между видами растений и грибными симбионтами в ответ на травоядность или условия окружающей среды.[38] Сильная активность нескольких биологически активных соединений, продуцируемых эпихлоэ, против травоядных [21][26] и относительно умеренное прямое влияние эпихлоев на рост и физиологию растений[39][40] предполагают, что эти соединения играют важную роль в сохранении симбиоза.
Рекомендации
- ^ а б Фрис, Э.М. (1849). Summa Vegetabilium Scandinaviae (на латыни). Стокгольм, Лейпциг: Бонье. п. 572.
- ^ Persoon, C.H. (1798). Icones et Descriptiones Fungorum Minus Cognitorum (на латыни). Лейпциг: Bibliopolii Breitkopf-Haerteliani impensis.

- ^ Туласне, Л.Р .; Туласне, К. (1865). Selecta Fungorum Carpologia: Nectriei - Phacidiei - Pezizei (на латыни). 3.
- ^ а б c d Leuchtmann, A .; Bacon, C.W .; Schardl, C.L .; White, J. F .; Тадыч, М. (2014). "Номенклатурная перестройка Неотифодиум виды с родом Эпихлоя" (PDF). Микология. 106 (2): 202–215. Дои:10.3852/13-251. ISSN 0027-5514. PMID 24459125. S2CID 25222557. Архивировано из оригинал (PDF) на 2016-03-07. Получено 2016-02-28.
- ^ White, J.F .; Редди, П.В. (1998). «Исследование строения и молекулярно-филогенетических связей некоторых злаковых симбионтов в родах». Эпихлоя и Парепихлоя". Микология. 90 (2): 226. Дои:10.2307/3761298. ISSN 0027-5514. JSTOR 3761298.
- ^ Morgan-Jones, G .; Гамс, В. (1982). «Заметки о гифомицетах. XLI. Эндофит Festuca arundinacea и анаморф Epichloe typhina, новые таксоны в одном из двух новых разрезов Акремониум". Микотаксон. 15: 311–318. ISSN 0093-4666.
- ^ Гленн А.Е., Бэкон К.В., Прайс Р., Хэнлин Р.Т. (1996). «Молекулярная филогения Акремониум и его таксономические последствия ". Микология. 88 (3): 369–383. Дои:10.2307/3760878. JSTOR 3760878.
- ^ Спатафора Дж. В., Сун Г. Х., Сун Дж. М., Хиуэл-Джонс Н. Л., Уайт Дж. Ф. младший (2007). «Филогенетические доказательства происхождения патогенов спорыньи и травяных эндофитов». Мол. Ecol. 16 (8): 1701–1711. Дои:10.1111 / j.1365-294X.2007.03225.x. PMID 17402984.
- ^ а б c d Schardl CL, Leuchtmann A, Spiering MJ (2004). «Симбиозы злаков с грибными эндофитами семенного происхождения». Анну Рев Завод Биол. 55: 315–340. Дои:10.1146 / annurev.arplant.55.031903.141735. PMID 15377223.
- ^ Фримен Э.М. (1904). "Семенной гриб Lolium temulentum Л., Дарнель ". Философские труды Лондонского королевского общества, серия B. 196 (214–224): 1–27. Дои:10.1098 / рстб.1904.0001.
- ^ Бланкеншип Дж. Д., Спиринг М. Дж., Уилкинсон Х. Х., Фаннин Ф. Ф., Буш LP, Шардл С. Л. (2001). "Производство алкалоидов лолина эндофитом травы, Neotyphodium uncinatum, в определенных носителях ". Фитохимия. 58 (3): 395–401. Дои:10.1016 / S0031-9422 (01) 00272-2. PMID 11557071.
- ^ Górzyńska, K .; и другие. (2010). "Необычный Ботанофила–Эпихлоя ассоциация в популяции садовых трав (Dactylis glomerata) в Польше". Журнал естественной истории. 44 (45–46): 2817–24. Дои:10.1111 / j.1570-7458.2006.00518.x.
- ^ Шардл К.Л., Крейвен К.Д., Спикман С., Стромберг А., Линдстрем А., Ёсида Р (2008). «Новый тест на совместное расхождение между хозяином и симбионтом указывает на древнее происхождение грибковых эндофитов в травах». Syst. Биол. 57 (3): 483–498. arXiv:q-bio / 0611084. Дои:10.1080/10635150802172184. PMID 18570040. S2CID 2703622.
- ^ Tan YY, Spiering MJ, Scott V, Lane GA, Christensen MJ, Schmid J (2001). «In planta регуляция роста эндофитного гриба и поддержание высоких уровней метаболизма в его мицелии при отсутствии апикального распространения». Appl. Environ. Микробиол. 67 (12): 5377–5383. Дои:10.1128 / AEM.67.12.5377-5383.2001. ЧВК 93319. PMID 11722882.
- ^ Кристенсен MJ, Беннетт RJ, Шмид J (2002). "Рост Эпихлоя / Неотифодиум и р-эндофиты в листьях Лолиум и Festuca травы ». Mycol. Res. 96: 93–106. Дои:10.1017 / S095375620100510X.
- ^ Кристенсен М.Дж., Беннетт Р.Дж., Ансари Н.А., Кога Х., Джонсон Р.Д., Брайан Г.Т., Симпсон В.Р., Кулаард Дж.П., Никлесс Э.М., Voisey CR (2008). «Эндофиты эпихлои растут за счет удлинения вставочного гифа в удлиненных листьях травы». Fungal Genet. Биол. 45 (2): 84–93. Дои:10.1016 / j.fgb.2007.07.013. PMID 17919950.
- ^ Танака А., Кристенсен М.Дж., Такемото Д., Парк П., Скотт Б. (2006). «Активные формы кислорода играют роль в регулировании мутуалистического взаимодействия между грибами и многолетним райграсом». Растительная клетка. 18 (4): 1052–1066. Дои:10.1105 / tpc.105.039263. ЧВК 1425850. PMID 16517760.
- ^ Такемото Д., Танака А., Скотт Б. (2006). «Регулятор, подобный p67Phox, задействован для контроля ветвления гиф в мутуалистическом симбиозе грибов и трав». Растительная клетка. 18 (10): 2807–2821. Дои:10.1105 / tpc.106.046169. ЧВК 1626622. PMID 17041146.
- ^ Цай Х.Ф., Лю Дж.С., Стабен С., Кристенсен М.Дж., Latch GC, Сигель М.Р., Шардл С.Л. (1994). «Эволюционная диверсификация грибковых эндофитов овсяницы высокорослой путем гибридизации с Эпихлоя разновидность". Proc. Natl. Акад. Sci. Соединенные Штаты Америки. 91 (7): 2542–2546. Bibcode:1994PNAS ... 91.2542T. Дои:10.1073 / пнас.91.7.2542. ЧВК 43405. PMID 8172623.
- ^ Moon CD, Craven KD, Leuchtmann A, Clement SL, Schardl CL (2004). «Распространенность межвидовых гибридов среди бесполых грибковых эндофитов трав». Мол Экол. 13 (6): 1455–1467. Дои:10.1111 / j.1365-294X.2004.02138.x. PMID 15140090.
- ^ а б c d е ж Буш LP, Уилкинсон HH, Schardl CL (1997). «Биозащитные алкалоиды травяно-грибковых эндофитных симбиозов». Физиология растений. 114 (1): 1–7. Дои:10.1104 / pp.114.1.1. ЧВК 158272. PMID 12223685.
- ^ Скотт Б. (2001). "Эпихлоя эндофиты: грибные симбионты трав ». Curr. Мнение. Микробиол. 4 (4): 393–398. Дои:10.1016 / S1369-5274 (00) 00224-1. PMID 11495800.
- ^ Робертс CA, West CP, Spiers DE, eds (2005). Неотифодиум в травах прохладного сезона. Блэквелл. ISBN 978-0-8138-0189-6.CS1 maint: несколько имен: список авторов (ссылка на сайт) CS1 maint: дополнительный текст: список авторов (ссылка на сайт)
- ^ а б Schardl CL, Panaccione DG, Tudzynski P (2006). Алкалоиды спорыньи - биология и молекулярная биология. Алкалоиды: химия и биология. 63. С. 45–86. Дои:10.1016 / S1099-4831 (06) 63002-2. ISBN 9780124695634. PMID 17133714.
- ^ Молодой CA, Фелитти С., Шилдс К., Спангенберг Г., Джонсон Р. Д., Брайан Г. Т., Сайкия С., Скотт Б. (2006). «Сложный кластер генов для биосинтеза индол-дитерпена в эндофите травы. Неотифодиум лолии". Грибок Genet Biol. 43 (10): 679–693. Дои:10.1016 / j.fgb.2006.04.004. PMID 16765617.
- ^ а б Танака А., Таппер Б.А.; Попай А., Паркер; EJ, Скотт Б. (2005). «Симбиоз экспрессируемой не рибосомальной пептидной синтетазы из мутуалистического грибкового эндофита райграса многолетнего обеспечивает защиту симбиотума от травоядных насекомых». Мол. Микробиол. 57 (4): 1036–1050. Дои:10.1111 / j.1365-2958.2005.04747.x. PMID 16091042.
- ^ а б Шардл С.Л., Гроссман Р.Б., Нагабхиру П., Фолкнер-младший, Маллик Ю.П. (2007). «Лолайн алкалоиды: валюта мутуализма». Фитохимия. 68 (7): 980–996. Дои:10.1016 / j.phytochem.2007.01.010. PMID 17346759.
- ^ Бланкеншип Дж. Д., Хаускнехт Дж. Б., Пал С., Буш Л. П., Гроссман Р. Б., Шардл С. Л. (2005). «Биосинтетические предшественники грибных пирролизидинов, алкалоидов лолина». ChemBioChem. 6 (6): 1016–1022. Дои:10.1002 / cbic.200400327. PMID 15861432. S2CID 13461396.
- ^ Zhang, DX, Nagabhyru, P, Schardl CL (2009). «Регулирование химической защиты против травоядных, продуцируемых симбиотическими грибами у травяных растений». Физиология растений. 150 (2): 1072–1082. Дои:10.1104 / стр.109.138222. ЧВК 2689992. PMID 19403726.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Гонтье DJ, Салливан Т.Дж., Браун К.Л., Вуртцель Б., Лаваль Р., Ванден Увер К., Бьюкен З., Бултман Т.Л. (2008). «Стромообразующий эндофит Epichloe glyceriae обеспечивает травоядному хозяину травоядное сопротивление, вызванное раной ". Ойкос. 117 (4): 629–633. Дои:10.1111 / j.0030-1299.2008.16483.x.
- ^ Баркер GM; Патчетт Би Джей; Кэмерон NE (2014). «Инфекция Epichloë uncinata и содержание лолина обеспечивают защиту трав Festulolium от черного жука (Heteronychus arator). Landcare Research, Гамильтон, Новая Зеландия и Cropmark Seeds Ltd, Крайстчерч, Новая Зеландия, 20 декабря 2014 г.». Новозеландский журнал сельскохозяйственных исследований. 58: 35–56. Дои:10.1080/00288233.2014.978480. S2CID 85409823.
- ^ Эвальд П.В. (1987). «Способы передачи и эволюция континуума паразитизма-мутуализма». Энн Нью-Йорк Академия наук. 503 (1): 295–306. Bibcode:1987НЯСА.503..295E. Дои:10.1111 / j.1749-6632.1987.tb40616.x. PMID 3304078.
- ^ Малиновский Д.П., Белеский Д.П. (2000). «Адаптация инфицированных эндофитами трав прохладного сезона к стрессам окружающей среды: механизмы засухи и устойчивости к минеральным стрессам». Crop Sci. 40 (4): 923–940. Дои:10.2135 / сельскохозяйственных культурci2000.404923x.
- ^ Сайкконен К., Ион Д., Гилленберг М. (2002). «Сохранение вертикально передающихся грибов в метапопуляциях травы». Proc Biol Sci. 269 (1498): 1397–1403. Дои:10.1098 / rspb.2002.2006. ЧВК 1691040. PMID 12079664.
- ^ Сайкконен К., Лехтонен П., Хеландер М., Коричева Дж., Фаэт С.Х. (2006). «Модельные системы в экологии: обзор эндофитно-травяной литературы». Тенденции Plant Sci. 11 (9): 428–433. Дои:10.1016 / j.tplants.2006.07.001. PMID 16890473.
- ^ Клей К., Холах Дж., Раджерс Дж. А. (2005). «Травоядные вызывают быстрое увеличение наследственного симбиоза и изменяют состав растительного сообщества». Proc. Natl. Акад. Sci. Соединенные Штаты Америки. 102 (35): 12465–12470. Bibcode:2005PNAS..10212465C. Дои:10.1073 / pnas.0503059102. ЧВК 1194913. PMID 16116093.
- ^ Кон С., Хик Д.С. (2007). «Травоядность опосредует отношения между травой и эндофитом». Экология. 88 (11): 2752–2757. Дои:10.1890/06-1958.1. PMID 18051643.
- ^ Гранат Г., Викари М., Базели Д.Р., Болл Дж. П., Пуэнтес А., Ракочевич Т. (2007). «Изменение численности грибковых эндофитов в овсянице по высотным и пастбищным уклонам». Экография. 30 (3): 422–430. Дои:10.1111 / j.0906-7590.2007.05027.x.
- ^ Хан Х, Макманус М.Т., Варнсторфф К., Монахан Б.Дж., Янг Калифорния, Дэвис Е., Таппер Б.А., Скотт, Б. (2007). "Неотифодиум грибковые эндофиты обеспечивают физиологическую защиту райграса многолетнего (Lolium perenne L.) испытывает недостаток воды ». Env. Exp. Бот. 63 (1–3): 183–199. Дои:10.1016 / j.envexpbot.2007.10.021.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Охота MG .; Расмуссен S; Ньютон PCD; Парсонс AJ; Ньюман Дж. А. (2005). "Краткосрочное воздействие повышенного содержания CO2, азота и грибковой инфекции эндофитов на Lolium perenne L. рост, химический состав и производство алкалоидов ». Растительная клеточная среда. 28 (11): 1345–1354. Дои:10.1111 / j.1365-3040.2005.01367.x.