Нейрула - Википедия - Neurula

А нейрула это позвоночное животное эмбрион на ранней стадии развития, в которой нейруляция происходит. Стадии нейрулы предшествует гаструла сцена; следовательно, нейруляции предшествует гаструляция.[1] Невруляция знаменует собой начало процесса органогенез.[2]
Мыши, цыплята и лягушки - обычные экспериментальные модели для изучения нейрулы. В зависимости от вида эмбрионы достигают стадии нейрулы в разное время и проводят разное количество времени на этой стадии.[3][4] За яйцекладущий Для организмов температура инкубации также влияет на продолжительность нейруляции.[2] Помимо разработки нервная трубка другие процессы происходят в эмбрионе на стадии нейрулы в зависимости от вида. Например, у рептилий внеэмбриональная мембрана ткани становятся отличными от эмбриона.[2]
Эмбрион нейрулы имеет пять областей мезодерма которые окружают нервную трубку.[5] Передняя мезодерма развивается в область головы, а задняя мезодерма - в туловище.[1] Различные молекулы, в том числе протеогликаны в внеклеточный матрикс, и гены, в том числе Pax Факторы транскрипции необходимы для развития и закрытия нервной трубки у эмбриона на стадии нейрулы.[6][7]
Невруляция
Невруляция это процесс у эмбрионов позвоночных на стадии нейрулы, в которой нервная трубка сформирован.[6][8] Есть два типа нейруляции: начальный и вторичная нейруляция. Первичная нейруляция относится к формированию и складыванию нервной пластинки внутрь самой себя с образованием нервной трубки.[6][8] При вторичной нейруляции нервная трубка формируется путем слияния полостей в мозговом канатике.[6][8][9] У амфибий и рептилий первичная нейруляция формирует целую нервную трубку, и нервная трубка закрывается одновременно по ее длине.[8] Напротив, у рыб вторичная нейруляция формирует нервную трубку.[10] Как первичная, так и вторичная нейруляция наблюдаются у птиц и млекопитающих, хотя и с небольшими различиями. Первичная нейруляция происходит в краниальных и верхних отделах спинного мозга, что дает начало головному и верхним отделам спинного мозга. Вторичная нейруляция происходит в нижних крестцовых и каудальных областях, в результате чего формируются нижние области спинного мозга.[6][10] У птиц нервная трубка закрывается спереди назад, а у млекопитающих сначала закрывается середина, а затем оба конца.[8]
Время развития
Точка, в которой эмбрион достигает стадии нейрулы, различается у разных видов, в то время как у яйцекладущих организмов продолжительность нейруляции дополнительно зависит от температуры инкубации. В целом, чем ниже температура, тем больше продолжительность нейруляции. Куриные эмбрионы достигают стадии нейрулы на 2-й день после оплодотворения и подвергаются нейруляции до 5-го дня. Рептилии, включая крокодилов, ящериц и черепах, имеют тенденцию проводить более длительное время на стадии нейрулы.[2] Типичный эмбрион лягушки, инкубируемый при 18 ° C, представляет собой нейрулу на ранней стадии через 50 часов после оплодотворения и нейрулу на поздней стадии через 67 часов.[3] Эмбрион мыши начинает нейруляцию на 7,5 день беременности и остается в стадии нейрулы до дня 9.[4]
Морфология
В мезодерма эмбриона позвоночного на стадии нейрулы можно разделить на пять областей. Вентрально к нервная трубка это хордамезодерма. Боковой по обе стороны от нервной трубки находится параксиальная мезодерма, а промежуточная латеральная область нервной трубки - это промежуточная мезодерма. Четвертый регион - это латеральная пластинка мезодермы, а последняя область - головная мезенхима.[5] Передние части мезодермы развиваются в ростральные области организма, такие как голова, в то время как задняя часть мезодермы развивается в каудальные области, такие как туловище или хвост.[1] Параксиальная мезодерма, также называемая сомитной мезодермой, развивается в сомиты, блоки ткани, расположенные сегментарно. Сомиты, в свою очередь, дают начало позвонкам, ребрам, скелетным мышцам, хрящам, сухожилиям и коже.[8][11]
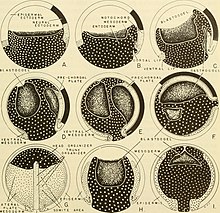
В Xenopus laevisпереход от гаструлы к нейруле включает морфологические изменения в двух областях, окружающих бластопор: дорсальной инволюционной маргинальной зоне (IMZ) и вышележащей неинволюционной маргинальной зоне (NIMZ) гаструлы. После инволюции на стадии средней гаструлы IMZ подвергается конвергентное расширение, при котором боковые области сужаются и смещаются к средней линии, а передний конец удлиняется. Это имеет эффект сужения бластопор. NIMZ, который не является эвольвентным, одновременно расширяется в противоположном направлении и с большей скоростью, чтобы покрыть области, больше не занятые IMZ. Конвергентное расширение IMZ и NIMZ начинается во второй половине гаструляции и продолжается до стадии поздней нейрулы. В конечном итоге глубокая ткань IMZ образует центральную нотохорда и окружающие параксиальная мезодерма. К ранней стадии нейрулы отчетливо выделяется хорда. Нотохордовые клетки объединяются в формацию, представляющую стопку монет, в процессе, называемом кольцевым интеркалированием. Поверхностный слой ИМЗ перерастает в кровлю архентерон, или примитивный кишечник, в то время как лежащий в основе энтодерма образует пол архентерона. НИМЗ развивается в структуру, напоминающую раннюю нервная трубка. Внешний эктодермальный слой нейрулы образован за счет равномерного разрастания клеток в полюс животного, известный как шапка животного. В эктодерма затем дифференцируется на нервный и эпидермальный ткань.[12]
У рептильных эмбрионов, начиная с поздней стадии нейрулы и переходя на ранние стадии органогенеза, внеэмбриональная мембрана ткани, содержащие желточный мешок, хорион, и амнион становятся отличными от тканей эмбриона. В мезодерма расщепляется с образованием внеэмбрионального целома, состоящего из двух слоев. Васкуляризированный внутренний слой мезодермы-энтодермы, называемый спланхноплеврой, развивается в желточный мешок, в то время как неваскуляризованный внешний слой эктодермы-мезодермы, называемый соматоплеврой, становится амнионом и хорионом. Во время органогенеза эти три внеэмбриональные ткани становятся полностью развитыми. Кроме того, в нейруле рептилий ткани мозга начинают дифференцироваться, и начинают формироваться сердце и кровеносные сосуды.[2]
Химический состав
Ткани нейрулы мыши делятся быстро, в среднем клеточный цикл продолжительностью 8–10 часов. Протеогликаны в внеклеточный матрикс (ECM) клеток на стадии нейрулы играют важную роль в обеспечении функциональной нейруляции черепа и подъеме нервной складки; гиалуроновая кислота (HA) синтезируется и накапливается, в то время как клетка поддерживает низкий уровень сульфатированного гликозаминогликаны (GAGs). HA участвует в создании двояковыпуклых нервных складок, в то время как сульфатированные GAG играют решающую роль в преобразовании нервной борозды в V-образную форму, а также в закрытии нервной трубки. ECM не играет главной роли в нейруляции позвоночника из-за плотной упаковки мезодермальных клеток в области позвоночника, что оставляет мало межклеточного пространства. Кроме того, считается, что актин-содержащие микрофиламенты необходимы для нейруляции черепа. Они могут действовать как механизм нервной складки или они могут стабилизировать нервные складки, которые уже сформировались; однако их точная роль не установлена. Есть некоторые свидетельства того, что факторы роста, Такие как инсулин или же трансферрин, также играют роль в нейруляции, но эта связь недостаточно изучена.[6]
Активация гена
Было обнаружено, что в эмбрионе на стадии нейрулы экспрессируются различные гены. Разные гены активируются для разных событий нейруляции, например, происходящих в отдельных регионах развивающейся нервной трубки.[6] Эти гены необходимы для правильной нейруляции и закрытия нервной трубки. Сигнальные молекулы, такие как Wnts, FGFs, и BMFs наряду с факторами транскрипции, которые включают Msx, Улиткаs, Sox8 / 9/10, и Pax3/7 гены играют ключевую роль в формировании нервного гребня.[6]
Pax транскрипционные факторы играют важную роль в раннем развитии, особенно в отношении ЦНС и нервный гребень. Pax3 и Pax7 являются промоторами как выживания клеток нервного гребня, так и устойчивости к стрессу окружающей среды.[7] В эмбрионах мыши Pax3 блокирует ген-супрессор опухоли p53, который необходим для контролируемой пролиферации и стабильности генома, экспрессируется во всех клетках нейрулы.[6][7] Во время раннего развития Pax3 экспрессируется в задней и боковой областях нервной пластинки, в той же области, из которой возникает нервный гребень.[7] Было обнаружено, что дефекты нервного гребня встречаются у мутантов Pax3 мыши и человека, что указывает на важность функциональности.[7][13] У цыплят, лягушек и рыб Pax3 / Pax7 активируются с помощью передачи сигналов Wnt и FGF.[7] Pax3 и Pax7 также необходимы для индукции нервного гребня после того, как истощение двух генов привело к отсутствию активации специфических генов нервного гребня Snail2 и Foxd3, что не позволяло дальнейшее развитие или эмиграцию нервного гребня.[7] Использование нокаутов было полезно для понимания роли и функций нескольких генов, обнаруженных в нейруле. Например, Wnt-1 было обнаружено, что он не играет никакой роли в закрытии нервной пластинки, несмотря на то, что присутствует на кончике нервной складки, когда она закрывается. Хотя мутанты Wnt-1 действительно приводят к дефектам структуры мозга. Notch1 участвует в образовании сомитов. HNF-3 нужен для развития хорды и узла.[14] Ген Аполипопротеин B, который участвует в транспортировке и метаболизме жирорастворимых молекул в крови, экспрессируется в желточном мешке и печени плода.[6] Внутри нейрулы в Xenopus laevis, присутствуют гены развития Xwnt-3 и Xwnt-4.[15]
Рекомендации
- ^ а б c Стерн, Клаудио Д. (февраль 2001 г.). "Первоначальный паттерн центральной нервной системы: сколько организаторов?" (PDF). Обзоры природы Неврология. 2 (2): 92–98. Дои:10.1038/35053563. PMID 11252999.
- ^ а б c d е Эндрюс, Р. М. (2004). «Закономерности эмбрионального развития» (PDF). Инкубация рептилий: окружающая среда, эволюция и поведение: 75–102.
- ^ а б Хилл, M.A. (30 ноября 2018 г.) Эмбриология Развитие лягушки. Получено с https://embryology.med.unsw.edu.au/embryology/index.php/Frog_Development.
- ^ а б Хилл, M.A. (30 ноября 2018 г.) Эмбриология Подробная временная шкала мыши. Получено с https://embryology.med.unsw.edu.au/embryology/index.php/Mouse_Timeline_Detailed.
- ^ а б Гилберт, Скотт Ф. (2000). «Параксиальная и промежуточная мезодерма». Биология развития. 6-е издание.
- ^ а б c d е ж грамм час я j Флеминг, А; Геррелли, Д; Грин, Северная Дакота; Копп, А. Дж (2002-03-01). «Механизмы нормальной и аномальной нейруляции: данные исследований культуры эмбрионов». Международный журнал биологии развития. 41 (2). ISSN 0214-6282.
- ^ а б c d е ж грамм Монсоро-Бурк, Энн Х. (01.08.2015). «Факторы транскрипции PAX в развитии нервного гребня». Семинары по клеточной биологии и биологии развития. 44: 87–96. Дои:10.1016 / j.semcdb.2015.09.015. ISSN 1084-9521. PMID 26410165.
- ^ а б c d е ж Слончевский, Жанна. «Глава 14. Гаструляция и невруляция». biology.kenyon.edu. Получено 2018-11-16.
- ^ Симокита, Эйсуке; Такахаши, Ёсико (апрель 2011 г.). «Вторичная нейруляция: картирование судьбы и генная манипуляция нервной трубки в хвостовой зачатке». Развитие, рост и дифференциация. 53 (3): 401–410. Дои:10.1111 / j.1440-169x.2011.01260.x. ISSN 0012-1592. PMID 21492152.
- ^ а б Schoenwolf, Gary C .; Смит, Джоди Л. (2000), «Механизмы нейруляции», Протоколы биологии развития, Humana Press, 136, стр.125–134, Дои:10.1385/1-59259-065-9:125, ISBN 978-1592590650, PMID 10840705
- ^ Ларсен, Уильям Дж. (2001). Эмбриология человека (3-е изд.). Филадельфия, Пенсильвания: Черчилль Ливингстон. С. 53–86. ISBN 0-443-06583-7.
- ^ Келлер, Р. Э., Данильчик, М., Гимлих, Р., и Ши, Дж. (1985). «Функция и механизм конвергентного расширения во время гаструляции Xenopus laevis "(PDF). Разработка, 89(Приложение), 185-209.
- ^ Тассабехджи, Майада; Прочтите, Эндрю П .; Ньютон, Валери Э .; Харрис, Родни; Баллинг, Руди; Грусс, Питер; Страчан, Том (1992-02-13). «Пациенты с синдромом Ваарденбурга имеют мутации в человеческом гомологе парного бокс-гена Pax-3». Природа. 355 (6361): 635–636. Дои:10.1038 / 355635a0. ISSN 0028-0836. PMID 1347148.
- ^ Lenkiewicz, E .; Ferencowa, A .; Szewczykowa, E. (2012-03-17). «Субконъюнктивальная аутогемотерапия ожогов глаз в наших случаях». Klinika Oczna. 94 (4): 113–4. Дои:10.4016/46352.01. PMID 1405409.
- ^ Christian, Jan L .; Гэвин, Брайан Дж .; McMahon, Andrew P .; Луна, Рэндалл Т. (1991-02-01). «Выделение кДНК, частично кодирующих четыре родственных XenopusWnt-1int-1 белка, и характеристика их временной экспрессии во время эмбрионального развития». Биология развития. 143 (2): 230–234. Дои:10.1016 / 0012-1606 (91) 90073-С. ISSN 0012-1606.