Взрослый нейрогенез - Википедия - Adult neurogenesis

{kind=link}

Взрослый нейрогенез это процесс, в котором нейроны генерируются из нервные стволовые клетки у взрослого. Этот процесс отличается от пренатальный нейрогенез.
У большинства млекопитающих новые нейроны рождаются во взрослом возрасте в двух областях мозга:[3]
- В субгранулярная зона (SGZ), часть зубчатые извилины из гиппокамп,[4][5] где нервные стволовые клетки рождают гранулярные клетки (участвует в формировании памяти и обучении).[нужна цитата ]
- В субвентрикулярная зона (СВЗ) боковые желудочки, можно разделить на три микродомена - латеральный, дорсальный и медиальный).[6] Нервные стволовые клетки мигрируют в обонятельная луковица сквозь ростральный миграционный поток где они дифференцируются на интернейроны участие в Чувство обоняния. У людей, однако, после рождения образуется мало нейронов обонятельной луковицы, если они вообще возникают.[7][8]
Больше внимания было уделено нейрогенезу в зубчатые извилины чем в полосатое тело. В грызуны, многие из нейронов зубчатой извилины новорожденных умирают вскоре после рождения,[4] но некоторые из них становятся функционально интегрированными в окружающую ткань мозга.[9][10][11] Количество нейронов, рожденных в гиппокампе человека, остается спорным; в некоторых исследованиях сообщается, что у взрослых людей каждый день в гиппокамп добавляется около 700 новых нейронов,[12] в то время как другие исследования показывают, что нейрогенез гиппокампа у взрослых людей не существует или, если он существует, находится на неопределяемом уровне.[13] Таким образом, роль новых нейронов в работе мозга взрослых остается неясной. Сообщается, что нейрогенез у взрослых играет роль в обучении и памяти, эмоциях, стрессе, депрессии, реакции на травмы и других состояниях.[14]
Механизм
Взрослые нервные стволовые клетки
Нейронный стволовые клетки (НСК) являются самообновляющимися, мультипотентный ячеек, которые генерируют основные фенотипы из нервная система.
Перепрограммирование клонов (транс-дифференциация)
Новые данные свидетельствуют о том, что перициты нервных микрососудов под руководством резидентных глиальных клеток перепрограммируются в интернейроны и обогащают местные нейронные микросхемы.[15] Этот ответ усиливается сопутствующим ангиогенез.
Модельные организмы нейрогенеза
Планарий
Планарий являются одними из самых ранних модельные организмы раньше учился регенерация с Паллада как основоположник планариеведения. Планарии - это классическая модель беспозвоночных, которая в последние десятилетия использовалась для изучения нейрогенеза. Центральная нервная система планарии проста, но полностью сформирована с двумя долями, расположенными в голове, и двумя. вентральные нервные тяжи. Эта модель воспроизводит бесполым путем, производя полную и полностью функционирующую нервную систему после разделения, что позволяет последовательно исследовать нейрогенез.
Аксолотль
В аксолотль используется реже, чем другие позвоночные, но по-прежнему является классической моделью для изучения регенерации и нейрогенеза. Хотя аксолотль нашел свое место в биомедицинских исследованиях с точки зрения регенерации конечностей,[16][17] модельный организм продемонстрировал сильную способность генерировать новые нейроны после повреждения.[18][19] Аксолотли сыграли роль связующего звена между беспозвоночными и млекопитающими, поскольку этот вид обладает регенеративной способностью проходить полный нейрогенез, формируя широкий спектр популяций нейронов, не ограничиваясь небольшой нишей,[20] тем не менее, сложность и архитектура сложны и во многом аналогичны развитию нервной системы человека.
Данио
Данио давно стали классикой развивающий модель за счет их прозрачности во время органогенез и широко использовались в нейрогенезе раннего развития.[21][22]). Рыбки данио демонстрируют сильную нейрогенеративную способность, способную регенерировать множество тканей и полное разнообразие нейронов (за исключением астроциты, так как они еще не идентифицированы в мозге рыбок данио) с продолжающимся нейрогенезом на протяжении всей жизни. В последние десятилетия эта модель укрепила свою роль в регенерации и нейрогенезе взрослых после повреждения.[23][24][25] Рыбки данио, как и аксолотли, сыграли ключевую роль в качестве промежуточного организма между беспозвоночными и млекопитающими. Рыбки данио - это быстро развивающийся организм, который относительно недороги в обслуживании, при этом обеспечивая простоту генетических манипуляций и сложную нервную систему.
Цыпленок
Хотя птиц использовали в основном для изучения раннего эмбрионального развития, в последние десятилетия развивающиеся цыпленок сыграл решающую роль в изучении нейрогенеза и регенерации, поскольку молодой цыпленок способен к обновлению нейронов в молодом возрасте, но теряет нейрогенеративную способность во взрослом возрасте.[26] Утрата нейрорегенеративной способности по мере созревания позволила исследователям продолжить изучение генетических регуляторов нейрогенеза.
Грызунов
Грызунов, мыши и крысы, были наиболее известными модельными организмами с момента открытия современных нейронов Сантьяго Рамон-и-Кахаль. Грызуны имеют очень похожую архитектуру и сложную нервную систему с очень небольшой регенерационной способностью, как у людей. По этой причине грызуны широко используются в доклинические испытания. Грызуны демонстрируют широкий спектр нейронных цепей, ответственных за сложное поведение, что делает их идеальными для изучения дендритной обрезки и аксональной обрезки.[27] В то время как организм представляет собой сильный человеческий аналог, модель имеет свои ограничения, которых не было в предыдущих моделях: более высокая стоимость содержания, меньшее количество размножающихся и ограниченные нейрогенеративные способности.
Осьминог
Также известен как обыкновенный осьминог а, головоногие моллюски, этот организм имеет сложную нервную систему, которая демонстрирует способность мозга производить новые клетки. В этом случае и в других таксонах, если сравнивать, эти организмы приспосабливаются к непредсказуемой окружающей среде, используя недавно сформированные клетки мозга.[28] Это короткая продолжительность жизни (самка около одного года), когда дикие обыкновенные осьминоги сосредотачивают большую часть своей энергии на спаривании и уходе за потомством.[29][30] Полученные данные предполагают, что осьминог обыкновенный, как и другие короткоживущие виды, имеет сложную пролиферацию гиппокампа,[31][32] необходим для пространственной / навигационной, краткосрочной и долгосрочной памяти.[33][циркулярная ссылка ]
Отслеживание нейрогенеза
Создание нового функционала нейроны можно измерить несколькими способами,[34] резюмируется в следующих разделах.
Маркировка ДНК
Маркированный ДНК может проследить происхождение делящейся клетки и определить местоположение ее дочерних клеток. А нуклеиновая кислота аналог вставлен в геном клетки, генерирующей нейрон (такой как глиальная клетка или же нервная стволовая клетка ).[35] Тимин аналоги (3H) тимидин[36] и Брду[37] обычно используются ДНК-метки и используются для радиоактивная маркировка и иммуногистохимия соответственно.
Определение судьбы по маркерам нейронального происхождения
ДНК маркировка может использоваться вместе с маркеры нейронального происхождения определить судьба нового функционала клетки мозга. Во-первых, включены помеченные нуклеотиды используются для выявления популяций новых разделенные дочерние клетки. Затем определяются конкретные типы клеток с уникальными различиями в их выражение из белки, который можно использовать как антигены в иммуноанализ. Например, NeuN / Fox3 и GFAP находятся антигены обычно используется для обнаружения нейроны, глия, и эпендимные клетки. Ki67 является наиболее часто используемым антигеном для обнаружения распространение клеток. Немного антигены может использоваться для измерения конкретных стволовая клетка этапы. Например, стволовым клеткам требуется sox2 ген для поддержания плюрипотентность и используется для определения устойчивой концентрации стволовых клеток в Ткань ЦНС. В белок нестин является промежуточная нить, что существенно для радиального роста аксоны, и поэтому используется для обнаружения образования новых синапсы.
Cre-Lox рекомбинация
Некоторые исследования генетического отслеживания используют Cre-lox рекомбинация связать промоутер к репортерный ген, Такие как lacZ или же Ген GFP.[35][38] Этот метод можно использовать для долгосрочной количественной оценки деления и маркировки клеток, тогда как ранее упомянутые процедуры полезны только для краткосрочной количественной оценки.
Вирусные векторы
В последнее время все чаще стали использовать рекомбинантный вирусы для вставки генетическая информация кодирование конкретных маркеры (обычно белок флуорофоры Такие как GFP ) которые только выразил в клетки определенного вида. Маркерный ген вставлен вниз по течению из промоутер, что приводит к транскрипция этого маркера только в ячейках, содержащих фактор (ы) транскрипции которые привязаны к этому промоутеру. Например, рекомбинантный плазмида может содержать промоутер для даблкортин, белок, экспрессируемый преимущественно нейроны, перед последовательностью, кодирующей GFP, тем самым делая инфицированные клетки флуоресценция зеленый при воздействии света от синего до ультрафиолетового диапазона[39] оставляя не даблкортин экспрессирующие клетки незатронутыми, даже если они содержат плазмида. Многие клетки будут содержать несколько копий плазмиды и самого флуорофора, что позволит передавать флуоресцентные свойства по инфицированной клетке. родословная.
Маркируя клетку, которая дает начало нейронам, например, нервные стволовые клетки или нейронные клетки-предшественники, можно отслеживать создание, пролиферацию и даже миграцию вновь созданных нейронов.[40] Однако важно отметить, что, хотя плазмида стабильна в течение длительных периодов времени, ее белковые продукты могут сильно варьировать. полужизни и их флуоресценция может уменьшаться, а также становиться слишком разбавленной, чтобы быть видимой, в зависимости от количества циклов репликации, которым они подверглись, что делает этот метод более полезным для отслеживания самоподобных нервных предшественников или нервных стволовых клеток, а не самих нейронов. Включение генетический материал через вирусный вектор имеет тенденцию быть спорадическим и нечастым по отношению к общему количеству клеток в данной области ткань, что делает количественную оценку клеточного деления неточной. Однако описанный выше метод может предоставить очень точные данные относительно того, когда ячейка была родившийся а также полная сотовая морфологии.[41]
Способы подавления нейрогенеза
Многие исследования, анализирующие роль нейрогенеза у взрослых, используют метод подавления пролиферации клеток в определенных областях мозга, имитируя подавление нейрогенеза, для наблюдения за влиянием на поведение.[14]
Фармакологическое ингибирование
Фармакологическое ингибирование широко используется в различных исследованиях, так как оно дает много преимуществ. Это обычно недорого по сравнению с другими методами, такими как облучение, может использоваться для различных видов животных и не требует каких-либо инвазивных процедур или операций для субъектов.
Однако это действительно создает определенные проблемы, поскольку эти ингибиторы не могут использоваться для подавления пролиферации в определенных регионах, что приводит к неспецифическим эффектам со стороны других систем. Чтобы избежать этих эффектов, необходимо провести дополнительную работу по определению оптимальных доз, чтобы минимизировать воздействие на системы, не связанные с нейрогенезом.
Обычным фармакологическим ингибитором нейрогенеза у взрослых является метилазоксиметанолацетат (МАМ), химиотерапевтическое средство. Другими ингибиторами клеточного деления, обычно используемыми в исследованиях, являются цитарабин и темозоломид.
Фармакогенетика
Другой метод, используемый для изучения эффектов нейрогенеза у взрослых, - это использование фармакогенетических моделей. Эти модели обеспечивают различные преимущества от фармакологического пути, поскольку они обеспечивают большую специфичность за счет нацеливания на конкретные предшественники нейрогенеза и определенные промоторы стволовых клеток. Это также учитывает временную специфичность взаимодействия определенных лекарств. Это полезно для изучения нейрогенеза в зрелом возрасте после нормального развития других областей мозга.
Тимидинкиназа вируса простого герпеса (HSV-TK) использовалась в исследованиях в сочетании с противовирусными препаратами для подавления нейрогенеза у взрослых. Он работает путем нацеливания на стволовые клетки с использованием глиальных фибриллярных кислых белков и экспрессии нестина. Эти стволовые клетки-мишени подвергаются гибели клеток вместо пролиферации при воздействии противовирусных препаратов.
Белок Cre также обычно используется для нацеливания на стволовые клетки, которые претерпевают генные изменения при лечении тамоксифеном.
Облучение
Облучение - это метод, который позволяет очень специфично подавлять нейрогенез у взрослых. Он может быть направлен на мозг, чтобы избежать воздействия на другие системы и неспецифических эффектов. Его даже можно использовать для нацеливания на определенные области мозга, что важно для определения того, как нейрогенез взрослых в различных областях мозга влияет на поведение.
Однако облучение дороже, чем другие методы, а также требует большого оборудования с обученными людьми.
Подавление нейрогенеза взрослых в гиппокампе
Многие исследования наблюдали, как ингибирование нейрогенеза взрослых у других млекопитающих, таких как крысы и мыши, влияет на их поведение.[14] Было показано, что ингибирование нейрогенеза взрослых в гиппокампе оказывает различное влияние на обучение и память, обусловленность и исследовательское поведение.
Ослабленная обусловленность страха наблюдалась в исследованиях с участием крыс с отсутствием нейрогенеза у взрослых в гиппокампе.[42] Подавление нейрогенеза взрослых в гиппокампе также было связано с изменениями в поведении при выполнении задач, связанных с исследованием.[43] Крысы также демонстрируют сниженное контекстуализированное замораживание в ответ на контекстуализированный страх и нарушение обучения пространственным местоположениям при отсутствии нейрогенеза у взрослых.[44][45]
Влияние на разделение рисунков
Считается, что изменения в обучении и памяти, наблюдаемые в упомянутых ранее исследованиях, связаны с ролью взрослого нейрогенеза в регуляции разделения паттернов.[14] Разделение шаблонов определяется как «процесс удаления избыточности из одинаковых входных данных, чтобы события можно было отделить друг от друга и уменьшить помехи, и, кроме того, он может производить более ортогональный, разреженный и категоризированный набор выходных данных».[46]
Это нарушение разделения паттернов могло бы объяснить нарушения, наблюдаемые при выполнении других задач обучения и памяти. Снижение способности уменьшать помехи может привести к большим трудностям в формировании и сохранении новых воспоминаний.,[14] хотя трудно различить эффекты нейрогенеза в обучении и разделения паттернов из-за ограничений в интерпретации поведенческих результатов ».[47]
Исследования показывают, что крысы с подавленным нейрогенезом взрослых демонстрируют трудности в дифференциации и обучении контекстуализированному условию страха.[14] Крысы с заблокированным нейрогенезом взрослых также демонстрируют нарушение дифференциального замораживания, когда им необходимо различать сходные условия.[48] Это также влияет на их пространственное распознавание в тестах лабиринта с радиальными рукавами, когда руки расположены ближе друг к другу, а не дальше друг от друга.[49] Мета-анализ поведенческих исследований, оценивающих влияние нейрогенеза в различных тестах разделения паттернов, показал постоянное влияние абляции нейрогенеза на производительность, хотя в литературе есть исключения. "[50]
Влияние на поведенческое торможение
Поведенческое торможение важно для крыс и других животных, когда они останавливают все, что они делают в данный момент, чтобы переоценить ситуацию в ответ на угрозу или что-то еще, что может потребовать их внимания.[14]
Крысы с пораженным гиппокампом демонстрируют меньшее сдерживание поведения при воздействии угроз, таких как запах кошки.[51] Нарушение нормальной клеточной пролиферации и развитие зубчатой извилины у развивающихся крыс также ухудшает их реакцию замирания, которая является примером подавления поведения, при воздействии на незнакомого взрослого самца крысы.[52]
Это нарушение поведенческого торможения также связано с процессом обучения и памяти, поскольку подавление неправильных ответов или поведения требует способности подавлять эту реакцию.[14]
Подразумеваемое
Роль в обучении
Функциональная значимость нейрогенеза у взрослых сомнительна,[53] но есть некоторые свидетельства того, что нейрогенез гиппокампа у взрослых важен для учусь и объем памяти.[54] Было предложено множество механизмов взаимосвязи между усилением нейрогенеза и улучшением познания, включая вычислительные теории, демонстрирующие, что новые нейроны увеличивают объем памяти,[55] уменьшить интерференцию между воспоминаниями,[56] или добавить информацию о времени в воспоминания.[57] Учитывая, что скорость нейрогенеза существенно не меняется во взрослом возрасте, было высказано предположение, что уникальные эпизодические воспоминания могут быть созданы, просто полагаясь на возросшую способность молодых нейронов определенного возраста создавать стабильные новые синапсы со сверстниками, представляющими уникальные особенности события, которое нужно запомнить [58] Эксперименты, направленные на устранение нейрогенеза, оказались безрезультатными, но несколько исследований предложили нейрогенную зависимость в некоторых типах обучения,[59] и другие не видят никакого эффекта.[60] Исследования показали, что сам акт обучения связан с увеличением выживаемости нейронов.[61] Однако общие выводы о том, что нейрогенез взрослых важен для любого вида обучения, неоднозначны.
Болезнь Альцгеймера
Некоторые исследования показывают, что снижение нейрогенеза в гиппокампе может привести к развитию Болезнь Альцгеймера (ОБЪЯВЛЕНИЕ).[62] Тем не менее, другие предполагают, что у пациентов с БА увеличен нейрогенез в области СА1 рога Аммона (основная область патологии гиппокампа при БА), чтобы компенсировать потерю нейронов.[63] Хотя точная природа взаимосвязи между нейрогенезом и болезнью Альцгеймера неизвестна, инсулиноподобный фактор роста 1 -стимулированный нейрогенез вызывает серьезные изменения в пластичности гиппокампа и, по-видимому, участвует в патологии Альцгеймера.[64] Аллопрегнанолон, а нейростероид, способствует продолжению нейрогенеза в головном мозге. Уровень аллопрегнанолона в головном мозге снижается старость и болезнь Альцгеймера.[65] Аллопрегнанолон, обращая вспять нарушение нейрогенеза, обращает вспять когнитивные нарушения в модель мыши болезни Альцгеймера.[66] Рецепторы Eph и эфрин передача сигналов регулирует нейрогенез взрослых в гиппокамп и были изучены как потенциальные мишени для лечения некоторых симптомов AD.[67] Молекулы, связанные с патологией БА, в том числе ApoE, PS1 и ПРИЛОЖЕНИЕ, также было обнаружено, что они влияют на нейрогенез взрослых в гиппокампе.[68]
Роль в шизофрении
Исследования показывают, что люди с шизофрения имеют уменьшенный объем гиппокампа, что, как полагают, вызвано уменьшением нейрогенеза у взрослых. Соответственно, это явление может быть основной причиной многих симптомов болезни. Кроме того, в нескольких исследовательских работах упоминается, что четыре гена, связывающий дистробревин белок 1 (DTNBP1), нейрегулин 1 (NRG1), нарушенный при шизофрении 1 (DISC1), и рецептор нейрегулина 1 (ERBB4), возможно, ответственны за этот дефицит в норме. регенерация нейронов.[69][70] Сходство между депрессией и шизофренией предполагает возможную биологическую связь между этими двумя заболеваниями. Однако необходимо провести дальнейшие исследования, чтобы четко продемонстрировать эту взаимосвязь.[71]
Нейрогенез у взрослых и большое депрессивное расстройство
Исследования показывают, что нейрогенез гиппокампа у взрослых обратно пропорционален большому депрессивному расстройству (БДР).[72] Нейрогенез снижен в гиппокампе животных, моделирующих большое депрессивное расстройство, и при многих методах лечения этого расстройства, включая антидепрессант лекарства и электрошоковой терапии, увеличивают нейрогенез гиппокампа. Было высказано предположение, что снижение нейрогенеза в гиппокампе у людей с большим депрессивным расстройством может быть связано с высоким уровнем гормонов стресса, называемых глюкокортикоиды, которые также связаны с расстройством. Гиппокамп инструктирует гипоталамо-гипофизарно-надпочечниковая система для выработки меньшего количества глюкокортикоидов при высоком уровне глюкокортикоидов. Таким образом, неисправный гиппокамп может объяснить хронически высокие уровни глюкокортикоидов у людей с большим депрессивным расстройством. Однако некоторые исследования показали, что нейрогенез гиппокампа не ниже у людей с большим депрессивным расстройством и что уровни глюкокортикоидов в крови не изменяются при изменении нейрогенеза гиппокампа, поэтому связь все еще не ясна.
Стресс и депрессия
Многие теперь считают, что стресс - самый важный фактор в возникновении депрессия, помимо генетики. Как обсуждалось выше, клетки гиппокампа чувствительны к стрессу, который может привести к снижению нейрогенеза. Эту область чаще рассматривают при изучении причин и методов лечения депрессии. Исследования показали, что удаление надпочечник у крыс вызывали усиление нейрогенеза в зубчатые извилины.[73] Надпочечник отвечает за производство кортизол в ответ на стрессор, вещество, которое при постоянном производстве вызывает снижение регуляции серотонин рецепторы и подавляет рождение нейронов.[74] В том же исследовании было показано, что введение кортикостерона нормальным животным подавляет нейрогенез - противоположный эффект.[73] Наиболее типичный класс антидепрессантов, назначаемых при этом заболевании: селективные ингибиторы обратного захвата серотонина (СИОЗС)[75] и их эффективность можно объяснить нейрогенезом. В нормальном мозге повышение серотонина вызывает подавление кортикотропин-рилизинг гормон (CRH) через подключение к гиппокампу. Он напрямую воздействует на паравентрикулярное ядро для уменьшения выброса CRH и подавления функции норадреналина в голубое пятно.[73] Поскольку CRH подавляется, снижение нейрогенеза, связанное с его повышенными уровнями, также обращено вспять. Это позволяет производить больше клеток головного мозга, в частности, рецептора 5-HT1a в зубчатой извилине гиппокампа, что, как было показано, улучшает симптомы депрессии. Обычно нейроны созревают от трех до шести недель.[76] примерно столько же времени требуется для того, чтобы СИОЗС начали действовать. Эта корреляция усиливает гипотезу о том, что СИОЗС действуют посредством нейрогенеза, уменьшая симптомы депрессии. Некоторые нейробиологи скептически относятся к функциональному значению нейрогенеза, учитывая, что небольшое количество возникающих нейронов фактически интегрировано в существующие нейронные цепи. Однако в недавнем исследовании использовалось облучение формирующихся нейронов гиппокампа у нечеловеческих приматов (NHP), чтобы продемонстрировать, что нейрогенез необходим для антидепрессивной эффективности.[77]
Нейроны, рожденные взрослыми, по-видимому, играют роль в регуляции стресс.[78][79] Исследования связывают нейрогенез с благотворным действием определенных антидепрессанты, предполагая связь между снижением нейрогенеза в гиппокампе и депрессией.[80][81] В пионерском исследовании ученые продемонстрировали, что поведенческие преимущества приема антидепрессантов в мышей обращается, когда нейрогенез предотвращается с помощью рентгеновское облучение техники.[82] Фактически, новорожденные нейроны более возбудимы, чем старые нейроны из-за дифференциальной экспрессии ГАМК рецепторы.[83] Следовательно, правдоподобная модель состоит в том, что эти нейроны усиливают роль гиппокампа в механизме отрицательной обратной связи Ось HPA (физиологический стресс) и, возможно, в подавлении миндалина (область мозга, отвечающая за пугающие реакции на раздражители).[нечеткий ] В самом деле, подавление нейрогенеза взрослых может привести к усилению стрессовой реакции оси HPA в умеренно стрессовых ситуациях.[78] Это согласуется с многочисленными открытиями, связывающими деятельность по снятию стресса (обучение, воздействие новой, но благоприятной окружающей среды и упражнения) с повышенным уровнем нейрогенеза, а также с наблюдением, что животные подвергались физиологическому стрессу (кортизол) или психологический стресс (например, изоляция) показывают заметно сниженные уровни новорожденных нейронов. В условиях хронического стресса повышение уровня новорожденных нейронов антидепрессантами улучшает зависимый от гиппокампа контроль стрессовой реакции; Без новорожденных нейронов антидепрессанты не могут восстановить регуляцию стрессовой реакции, и выздоровление становится невозможным.[79]
Некоторые исследования выдвинули гипотезу, что обучение и память связаны с депрессией, и что нейрогенез может способствовать нейропластичность. Одно исследование предполагает, что настроение может регулироваться на базовом уровне с помощью пластичности, и, следовательно, не химия. Соответственно, эффекты лечения антидепрессантами будут вторичными по отношению к изменению пластичности.[84] Однако другое исследование продемонстрировало взаимодействие между антидепрессантами и пластичностью; Было показано, что антидепрессант флуоксетин восстанавливает пластичность мозга взрослых крыс.[85] Результаты этого исследования предполагают, что терапия антидепрессантами может способствовать не вторичному изменению пластичности.
Эффекты сокращения сна
Одно исследование связало недостаток сна с уменьшением нейрогенеза гиппокампа у грызунов. Предлагаемый механизм наблюдаемого снижения заключался в повышении уровня глюкокортикоиды. Было показано, что две недели недосыпание действовал как ингибитор нейрогенеза, который отменялся после восстановления нормального сна и даже переходил к временному увеличению нормальной пролиферации клеток.[86] Точнее, когда уровень кортикостерона повышен, лишение сна подавляет этот процесс. Тем не менее, нормальный уровень нейрогенеза после хронического недосыпания возвращается через 2 недели с временным усилением нейрогенеза.[87]Хотя это признано, упускается из виду потребность в глюкозе в крови во время временных диабетических гипогликемических состояний. Американская диабетическая ассоциация среди многих документирует псевдосенилию и возбуждение, обнаруживаемые во время временных гипогликемических состояний. Требуется гораздо больше клинической документации, чтобы компетентно продемонстрировать связь между снижением гематологического уровня глюкозы и нейрональной активностью и настроением.
Возможное использование при лечении болезни Паркинсона
болезнь Паркинсона это нейродегенеративное заболевание, характеризующееся прогрессирующей потерей дофаминергические нейроны в черная субстанция. Трансплантация дофаминергической клетки-предшественники проложил путь к возможности заместительной клеточной терапии, которая может улучшить клинические симптомы у пораженных пациентов.[88] В последние годы ученые предоставили доказательства существования нервных стволовых клеток, способных производить новые нейроны, особенно дофаминергического фенотипа, в мозге взрослых млекопитающих.[89][90][91] Экспериментальное истощение дофамина у грызунов снижает пролиферацию клеток-предшественников как в субэпендимальной зоне, так и в субгранулярной зоне.[92] Пролиферация полностью восстанавливается селективным агонистом D2-подобных (D2L) рецепторов.[92] Нервные стволовые клетки были идентифицированы в нейрогенных областях мозга, где нейрогенез постоянно продолжается, но также и в ненейрогенных зонах, таких как средний мозг и полосатое тело, где нейрогенез, как считается, не происходит в нормальных физиологических условиях.[88] Более новые исследования показали, что в полосатом теле действительно существует нейрогенез.[93]Подробное понимание факторов, управляющих взрослыми нейральными стволовыми клетками in vivo может в конечном итоге привести к элегантной клеточной терапии нейродегенеративных расстройств, таких как болезнь Паркинсона, путем мобилизации аутологичных эндогенных нервных стволовых клеток для замены дегенерированных нейронов.[88]
Травматическое повреждение мозга
Черепно-мозговые травмы различаются по механизму травмы, вызывая тупую или проникающую травму, приводящую к первичная и вторичная травма с эксайтотоксичность и относительно широко распространенная гибель нейронов. В связи с подавляющим числом черепно-мозговых травм в результате Война с терроризмом, огромное количество исследований было направлено на лучшее понимание патофизиологии черепно-мозговых травм, а также нейропротекторный вмешательства и возможные вмешательства, вызывающие восстановительный нейрогенез. Гормональные вмешательства, такие как прогестерон, эстроген и аллопрегнанолон, интенсивно изучались в последние десятилетия как возможные нейропротективные агенты после черепно-мозговых травм для уменьшения воспалительной реакции, замедляющей гибель нейронов.[94][95][96][97] У грызунов, не обладающих регенеративной способностью к нейрогенезу взрослых, активация стволовых клеток после введения агониста никотинового ацетилхолинового рецептора α7, PNU-282987, был идентифицирован в поврежденной сетчатке при последующей работе по изучению активации нейрогенеза у млекопитающих после черепно-мозговой травмы.[98] В настоящее время не прошло ни одного медицинского вмешательства. фаза-III клинические испытания для использования на людях.
Факторы, влияющие
Изменения в старости
Нейрогенез существенно снижен в гиппокампе старых животных, что повышает вероятность того, что это может быть связано с возрастным снижением функции гиппокампа. Например, скорость нейрогенеза у старых животных позволяет прогнозировать память.[99] Однако новорожденные клетки у старых животных функционально интегрированы.[100] Учитывая, что нейрогенез происходит на протяжении всей жизни, можно было ожидать, что гиппокамп будет неуклонно увеличиваться в размерах во взрослом возрасте, и, следовательно, количество гранулярных клеток будет увеличиваться у старых животных. Однако это не так, что указывает на то, что пролиферация уравновешивается гибелью клеток. Таким образом, кажется, что с функциями гиппокампа связано не добавление новых нейронов в гиппокамп, а скорее скорость обновления гранулярных клеток.[101]
Эффекты упражнений
Ученые показали, что физическая активность в виде произвольных упражнений приводит к увеличению количества новорожденных нейронов в гиппокампе мышей и крыс.[102][103] Эти и другие исследования показали, что обучение у обоих видов можно улучшить с помощью физических упражнений.[104] Недавние исследования показали, что нейротрофический фактор головного мозга и инсулиноподобный фактор роста 1 являются ключевыми медиаторами нейрогенеза, вызванного физической нагрузкой.[103][105] Физические упражнения увеличивают продукцию BDNF, а также субъединицу NR2B рецептора NMDA.[103] Физические упражнения увеличивают поступление IGF-1 из кровотока в различные области мозга, включая гиппокамп. Кроме того, IGF-1 изменяет экспрессию c-fos в гиппокампе. Когда IGF-1 заблокирован, упражнения больше не вызывают нейрогенез.[105] Другое исследование показало, что у мышей, которые не вырабатывали бета-эндорфин, гормон, повышающий настроение, нейрогенез не изменился. Тем не менее, у мышей, которые вырабатывали этот гормон, наряду с упражнениями, наблюдалось увеличение количества новорожденных клеток и их выживаемость.[106]Хотя связь между нейрогенезом, опосредованным упражнениями, и улучшением обучения остается неясной, это исследование может иметь серьезные последствия в области старения и / или Болезнь Альцгеймера.
Эффекты каннабиноидов
Эта секция нужно больше медицинские справки за проверка или слишком сильно полагается на основные источники. (Ноябрь 2016) |
Некоторые исследования показали, что стимуляция каннабиноиды приводит к росту новых нервных клеток в гиппокампе как из эмбриональных, так и из взрослых стволовых клеток. В 2005 году клиническое исследование крыс в Университете Саскачевана показало регенерацию нервных клеток в гиппокампе.[107] Исследования показали, что синтетический препарат, напоминающий THC, основной психоактивный ингредиент марихуаны, обеспечивает некоторую защиту от воспаление мозга, что может улучшить память в более старшем возрасте. Это связано с рецепторами в системе, которые также могут влиять на производство новых нейронов.[108]Тем не менее, исследование, проведенное в Университете Рутгерса, продемонстрировало, как синхронизация потенциалов действия в гиппокампе крыс изменилась после введения ТГК. Отсутствие синхронизации соответствовало снижению производительности в стандартном тесте памяти.[109]Недавние исследования показывают, что природный каннабиноид каннабиса, каннабидиол (CBD), увеличивает нейрогенез у взрослых, не влияя на обучение. Однако ТГК ухудшал обучение и не влиял на нейрогенез.[110] Более высокое соотношение CBD и THC в анализах волос у потребителей каннабиса коррелирует с защитой от уменьшения серого вещества в правом гиппокампе.[111] CBD также, как было замечено, ослабляет дефицит памяти прозы и зрительно-пространственной ассоциативной памяти у тех, кто в настоящее время находится под влиянием каннабиса,[112][113] подразумевая нейрозащитные эффекты против сильного воздействия ТГК. Нейрогенез может играть роль в его нейрозащитных эффектах, но необходимы дальнейшие исследования.
Несколько исследований сообщили о положительной связи между ТГК и нейрогенезом гиппокампа.[114][115] Некоторые из них предполагают двухфазный эффект,[114] некоторые из них выражают мнение, что часть негативных эффектов может быть связана с нейроадаптацией из-за воздействия в определенный период жизни и что ее можно обратить вспять.[116]
Регулирование
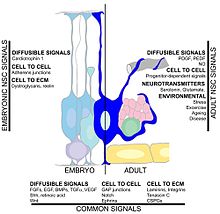
Многие факторы могут влиять на скорость нейрогенеза гиппокампа. Упражнение и обогащенная среда было показано, что способствует выживанию нейронов и успешной интеграции новорожденных клеток в существующий гиппокамп.[117][118][119][120] Другой фактор Центральная нервная система травма, так как нейрогенез происходит после церебральная ишемия,[121] Эпилептические припадки,[122] и бактериальный менингит.[123] С другой стороны, такие условия, как хронический стресс, вирусная инфекция. и старение может привести к снижению нейрональной пролиферации.[124][125][126][127]Факторы циркуляции в крови могут снизить нейрогенез. У здоровых стареющих людей уровни некоторых хемокинов в плазме и спинномозговой жидкости повышены. В модели на мышах уровни этих хемокинов в плазме коррелируют со сниженным нейрогенезом, предполагая, что нейрогенез может модулироваться некоторыми глобальными возрастными системными изменениями. Эти хемокины включают: CCL11, CCL2 и CCL12, которые сильно локализованы на хромосомах мыши и человека, что определяет генетический локус в старении.[54] Другое исследование включало цитокин IL-1beta, который продуцируется глией. Это исследование показало, что блокирование ИЛ-1 может частично предотвратить серьезное нарушение нейрогенеза, вызванное вирусной инфекцией.[128]
Эпигенетическая регуляция также играет большую роль в нейрогенезе. Метилирование ДНК имеет решающее значение в определении судьбы взрослых нервных стволовых клеток в субвентрикулярная зона для постнатального нейрогенеза посредством регуляции нейронных генов, таких как Dlx2, Neurog2, и Sp8. Много микроРНК такие как miR-124 и miR-9, как было показано, влияют на размер коры и наслоение во время развития.[129]
История
Ранние нейроанатомы, в том числе Сантьяго Рамон-и-Кахаль считали нервную систему неподвижной и неспособной к регенерации. Первые свидетельства нейрогенеза взрослых млекопитающих в кора головного мозга был представлен Джозеф Альтман в 1962 г.,[130] за которой последовала демонстрация взрослого нейрогенеза в зубчатой извилине гиппокампа в 1963 году.[131] В 1969 году Джозеф Альтман обнаружил и назвал ростральный миграционный поток в качестве источника взрослых нейронов из гранулярных клеток в обонятельной луковице.[132] Вплоть до 1980-х годов научное сообщество игнорировало эти результаты, несмотря на использование самого прямого метода демонстрации пролиферации клеток в ранних исследованиях, а именно авторадиографии 3H-тимидина. К тому времени Ширли Байер[133][134] (и Майкл Каплан ) снова показал, что нейрогенез у взрослых существует у млекопитающих (крыс), а Ноттебом показал то же явление у птиц.[135] вызвав новый интерес к теме. Исследования в 1990-е годы[136][137] наконец, поставили исследования нейрогенеза взрослых в основное русло. Также в начале 1990-х годов нейрогенез гиппокампа был продемонстрирован у нечеловеческих приматов и людей.[138][139] Совсем недавно был охарактеризован нейрогенез в мозжечке взрослых кроликов.[140] Кроме того, некоторые авторы (в частности, Элизабет Гулд ) предположили, что нейрогенез у взрослых также может происходить в областях мозга, обычно не связанных с нейрогенезом, включая неокортекс.[141][142][143] Однако другие[144] поставили под сомнение научное доказательство этих выводов, утверждая, что новые клетки может быть из глиальное происхождение. Недавние исследования выяснили регулирующий эффект ГАМК на нервные стволовые клетки. Хорошо известное тормозящее действие ГАМК на мозг также влияет на локальные схемы, которые заставляют стволовые клетки переходить в спящее состояние. Они обнаружили, что диазепам (Валиум) имеет аналогичный эффект.[145]
Смотрите также
- Искусственная нервная мембрана
- Нейронное развитие
- Нейропластичность
- Нейротрофин
- Невруляция
- Глиогенез
- Ноогенез
Рекомендации
- ^ Файз М; Акарин L; Castellano B; Гонсалес Б. (2005). «Динамика пролиферации клеток зародышевой зоны интактного и эксайтотоксически пораженного постнатального мозга крысы». BMC Neuroscience. 6: 26. Дои:10.1186/1471-2202-6-26. ЧВК 1087489. PMID 15826306.
- ^ Oomen CA; Girardi CE; Cahyadi R; и другие. (2009). Бауне, Бернхард (ред.). «Противоположные эффекты ранней материнской депривации на нейрогенез у самцов и самок крыс». PLOS ONE. 4 (1): e3675. Bibcode:2009PLoSO ... 4.3675O. Дои:10.1371 / journal.pone.0003675. ЧВК 2629844. PMID 19180242.
- ^ Эрнст, А; Фризен, Дж. (Январь 2015 г.). «Взрослый нейрогенез у человека - общие и уникальные черты у млекопитающих». PLOS Биология. 13 (1): e1002045. Дои:10.1371 / journal.pbio.1002045. ЧВК 4306487. PMID 25621867.
- ^ а б Dayer, A.G .; Ford, A.A .; Кливер, К.М .; Yassaee, M .; Кэмерон, Х.А. (2003). «Краткосрочное и долгосрочное выживание новых нейронов зубчатой извилины крысы». Журнал сравнительной неврологии. 460 (4): 563–572. Дои:10.1002 / cne.10675. PMID 12717714. S2CID 7153599.
- ^ Вадодария, Кришна Ч .; Гейдж, Фред Х. (2014). «SnapShot: нейрогенез гиппокампа взрослых». Клетка. 156 (5): 1114–1114.e1. Дои:10.1016 / j.cell.2014.02.029. PMID 24581504.
- ^ Фиорелли, Роберто; Азим, Касум; Фишер, Бруно; Рейнето, Оливье (15.06.2015). «Добавление пространственного измерения к нейрогенезу постнатальной желудочково-субвентрикулярной зоны». Разработка. 142 (12): 2109–2120. Дои:10.1242 / dev.119966. ISSN 1477-9129. PMID 26081572.
- ^ Эрнст А., Алкасс К., Бернар С., Салехпур М., Перл С., Тисдейл Дж., Посснерт Дж., Друид Х, Фризен Дж. (2014). «Нейрогенез в полосатом теле мозга взрослого человека». Клетка. 156 (5): 1072–83. Дои:10.1016 / j.cell.2014.01.044. PMID 24561062.
- ^ Bergmann, O .; Liebel, J .; Бернард, С .; Alkass, K .; Yeung, M.S.Y .; Steier, P .; Кучера, В .; Johnson, L .; Landen, M .; Друид, H .; Spalding, K.L .; Фризен, Дж. (2012). «Возраст нейронов обонятельной луковицы у человека». Нейрон. 74 (4): 634–639. Дои:10.1016 / j.neuron.2012.03.030. PMID 22632721.
- ^ Toni, N .; Teng, E.M .; Bushong, E.A .; Aimone, J.B .; Zhao, C .; Consiglio, A .; van Praag, H .; Martone, M.E .; Ellisman, M.H .; Гейдж, Ф.Х. (2007). «Формирование синапсов на нейронах, рожденных в гиппокампе взрослых». Природа Неврология. 10 (6): 727–734. Дои:10.1038 / nn1908. PMID 17486101. S2CID 6796849.
- ^ Vivar, C .; Potter, M.C .; Choi, J .; Lee, J .; Stringer, T.P .; Callawy, E.M .; Gage, F.H .; Suh, H .; ван Прааг, Х. (2012). «Моносинаптические входы в новые нейроны зубчатой извилины». Nature Communications. 3 (1038): 1107. Bibcode:2012 НатКо ... 3Э1107В. Дои:10.1038 / ncomms2101. ЧВК 4603575. PMID 23033083.
- ^ Toni, N .; Laplagne, D.A .; Zhao, C .; Lombardi, G .; Ribak, C.E .; Gage, F.H .; Шиндер, А.Ф. (2008). «Нейроны, рожденные во взрослой зубчатой извилине, образуют функциональные синапсы с клетками-мишенями». Природа Неврология. 11 (8): 901–907. Дои:10.1038 / №2156. ЧВК 2572641. PMID 18622400.
- ^ Spalding, Kirsty L .; Бергманн, Олаф; Алкасс, Канар; Бернард, Самуэль; Салехпур, Мехран; Huttner, Hagen B .; Бострем, Эмиль; Вестерлунд, Изабель; Флакон, Селин (06.06.2013). «Динамика нейрогенеза гиппокампа у взрослых людей». Клетка. 153 (6): 1219–1227. Дои:10.1016 / j.cell.2013.05.002. ISSN 1097-4172. ЧВК 4394608. PMID 23746839.
- ^ Сорреллс, Сан-Франциско; Паредес, М.Ф .; Кебриан-Силла, А; Сандовал, К; Ци, Д; Келли, кВт; Джеймс, Д.; Mayer, S; Чанг, Дж; Огюст, KI; Чанг, EF; Gutierrez, AJ; Кригштейн, АР; Mathern, GW; Олдхэм, MC; Huang, EJ; Гарсия-Вердуго, JM; Ян, Z; Альварес-Буйлла, А (15 марта 2018 г.). «Нейрогенез гиппокампа человека у детей резко падает до неопределяемого уровня у взрослых». Природа. 555 (7696): 377–381. Bibcode:2018Натура.555..377S. Дои:10.1038 / природа25975. ЧВК 6179355. PMID 29513649.
- ^ а б c d е ж грамм час Кэмерон, Хизер А .; Гловер, Лукас Р. (2015-01-03). «Взрослый нейрогенез: помимо обучения и памяти». Ежегодный обзор психологии. 66 (1): 53–81. Дои:10.1146 / annurev-psycho-010814-015006. ISSN 0066-4308. ЧВК 5612417. PMID 25251485.
- ^ Farahani, Ramin M .; Резаи ‐ Лотфи, Саба; Симонян, Мэри; Шаймардан, Мунира; Хантер, Нил (2019). «Нервные микрососудистые перициты вносят вклад в нейрогенез взрослого человека». Журнал сравнительной неврологии. 527 (4): 780–796. Дои:10.1002 / cne.24565. ISSN 1096-9861. PMID 30471080. S2CID 53711787.
- ^ Карлсон, Б. М. (декабрь 1975 г.). «Влияние вращения и изменения положения тканей культи на морфогенез регенерирующей конечности аксолотля». Биология развития. 47 (2): 269–291. Дои:10.1016/0012-1606(75)90282-1. HDL:1874/15361. ISSN 0012-1606. PMID 1204936.
- ^ Крагл, Мартин; Кнапп, Дунья; Наку, Евгений; Хаттак, Шахрияр; Маден, Малькольм; Эпперлейн, Ханс Хеннинг; Танака, Элли М. (02.07.2009). «Клетки сохраняют память о своем тканевом происхождении во время регенерации конечностей аксолотлей». Природа. 460 (7251): 60–65. Bibcode:2009Натура.460 ... 60К. Дои:10.1038 / природа08152. ISSN 1476-4687. PMID 19571878. S2CID 4316677.
- ^ Маден, Малькольм; Manwell, Laurie A .; Ормерод, Брэнди К. (17 января 2013 г.). «Зоны пролиферации в мозге аксолотлей и регенерация конечного мозга». Нейронное развитие. 8: 1. Дои:10.1186/1749-8104-8-1. ISSN 1749-8104. ЧВК 3554517. PMID 23327114.
- ^ Clarke, J.D .; Александр, Р .; Холдер Н. (1988-06-17). «Регенерация нисходящих аксонов спинного мозга аксолотля». Письма о неврологии. 89 (1): 1–6. Дои:10.1016/0304-3940(88)90471-5. ISSN 0304-3940. PMID 3399135. S2CID 23650500.
- ^ Амамото, Рёдзи; Уэрта, Виолетта Жиссель Лопес; Такахаши, Эми; Дай, Гуанпин; Грант, Аарон К.; Фу, Жанян; Арлотта, Паола (2016). «Взрослые аксолотли могут регенерировать изначальное разнообразие нейронов в ответ на травму головного мозга». eLife. 5. Дои:10.7554 / eLife.13998. ISSN 2050-084X. ЧВК 4861602. PMID 27156560.
- ^ Zupanc, Г. К. Х. (01.06.2006). «Нейрогенез и регенерация нейронов в мозге взрослых рыб». Журнал сравнительной физиологии А. 192 (6): 649–670. Дои:10.1007 / s00359-006-0104-y. ISSN 0340-7594. PMID 16463148. S2CID 24063389.
- ^ Шмидт, Ребекка; Стрэле, Уве; Шолпп, Штеффен (21 февраля 2013 г.). «Нейрогенез у рыбок данио - от эмбриона до взрослого человека». Нейронное развитие. 8: 3. Дои:10.1186/1749-8104-8-3. ISSN 1749-8104. ЧВК 3598338. PMID 23433260.
- ^ Хентиг, Джеймс Т .; Берд-Джейкобс, Кристин А. (31.08.2016). «Воздействие сульфата цинка приводит к различному воздействию на подтипы обонятельных сенсорных нейронов у взрослых рыбок данио». Международный журнал молекулярных наук. 17 (9): 1445. Дои:10.3390 / ijms17091445. ISSN 1422-0067. ЧВК 5037724. PMID 27589738.
- ^ Gorsuch, Ryne A .; Хайд, Дэвид Р. (июнь 2014 г.). «Регуляция мюллеровской глиальной зависимой регенерации нейронов в поврежденной сетчатке взрослых рыбок данио». Экспериментальные исследования глаз. 123: 131–140. Дои:10.1016 / j.exer.2013.07.012. ISSN 1096-0007. ЧВК 3877724. PMID 23880528.
- ^ Крёне, Фолькер; Фройденрайх, Дориан; Ганс, Стефан; Каслин, Ян; Бренд, Майкл (ноябрь 2011 г.). «Регенерация мозга взрослых рыбок данио из нейрогенных предшественников лучевой глии». Разработка. 138 (22): 4831–4841. Дои:10.1242 / dev.072587. ISSN 1477-9129. PMID 22007133.
- ^ Фишер, Энди Дж. (Март 2005 г.). «Нейронная регенерация сетчатки глаза цыплят». Прогресс в исследованиях сетчатки и глаз. 24 (2): 161–182. Дои:10.1016 / j.preteyeres.2004.07.003. ISSN 1350-9462. PMID 15610972. S2CID 43652371.
- ^ Джонс, Тереза А .; Шаллерт, Тимоти (1992-05-22). «Разрастание и обрезка дендритов у взрослых крыс, восстанавливающихся после повреждения неокортекса». Исследование мозга. 581 (1): 156–160. Дои:10.1016 / 0006-8993 (92) 90356-Е. PMID 1498666. S2CID 34248397.
- ^ Ди Космо, А. Д., Бертапель, К., Порчеллини, А., и Полезе, Г. (2018). Оценка величины нейрогенеза взрослых в мозге Octopus vulgaris с использованием метода на основе проточной цитометрии. Границы физиологии, 9. doi: 10.3389 / fphys.2018.01050
- ^ Ди Космо, А., Полесе, Г. (2014). «Головоногие встречаются с нейроэкологией: роль хеморецепции в репродуктивном поведении осьминога обыкновенного», в книге «Нейроэкология и нейроэтология моллюсков - интерфейс между поведением и окружающей средой», ред. А. Ди Космо и У. Винлоу (Нью-Йорк, Нью-Йорк: издательство NOVA Science) 117–132.
- ^ Polese G, Bertapelle C, Di Cosmo A (январь 2015 г.). «Роль обоняния в размножении Octopus vulgaris». Gen. Comp. Эндокринол. 210: 55–62. Дои:10.1016 / j.ygcen.2014.10.006. PMID 25449183.
- ^ Амрейн I, Липп HP (февраль 2009 г.). "Взрослый гиппокампальный нейрогенез млекопитающих: эволюция и история жизни". Биол. Латыш. 5 (1): 141–144. Дои:10.1098 / рсбл.2008.0511. ЧВК 2657751. PMID 18957357.
- ^ Амрейн И., Ислер К., Липп Х.П. (сентябрь 2011 г.). «Сравнение нейрогенеза гиппокампа у взрослых видов и отрядов млекопитающих: влияние хронологического возраста и стадии жизненного цикла». Евро. J. Neurosci. 34 (6): 978–87. Дои:10.1111 / j.1460-9568.2011.07804.x. PMID 21929629.
- ^ Гиппокамп
- ^ Aimone J. B .; Li Y .; Lee S. W .; Clemenson G.D .; Deng W .; Гейдж Ф. Х. (2014). «Регуляция и функции нейрогенеза взрослых: от генов к познанию». Физиологические обзоры. 94 (4): 991–1026. Дои:10.1152 / Physrev.00004.2014. ЧВК 4280160. PMID 25287858.
- ^ а б Мин Го-Ли, Сун Хунцзюнь (2011). «Взрослый нейрогенез в мозге млекопитающих: важные ответы и важные вопросы». Нейрон. 70 (4): 687–702. Дои:10.1016 / j.neuron.2011.05.001. ЧВК 3106107. PMID 21609825.
- ^ Альтман Дж. (1962). «Формируются ли новые нейроны в головном мозге взрослых млекопитающих?». Наука. 135 (3509): 1127–1128. Bibcode:1962Sci ... 135.1127A. Дои:10.1126 / science.135.3509.1127. PMID 13860748. S2CID 1606140.
- ^ Кун Х.Г., Купер-Кун СМ (2007). «Бромдезоксиуридин и обнаружение нейрогенеза». Curr Pharmaceutical Biotechnol. 8 (3): 127–131. Дои:10.2174/138920107780906531. PMID 17584085.
- ^ Имаёси И., Сакамото М., Оцука Т., Такао К., Миякава Т., Ямагути М., Мори К., Икеда Т., Итохара С., Кагеяма Р. (2008). «Роль непрерывного нейрогенеза в структурной и функциональной целостности переднего мозга взрослого человека». Nat Neurosci. 11 (10): 1153–1161. Дои:10.1038 / №2185. PMID 18758458. S2CID 11627647.
- ^ Prendergast, Franklyn G .; Манн, Кеннет Г. (1978-08-22). «Химические и физические свойства экворина и зеленого флуоресцентного белка, выделенного из Aequorea forskalea». Биохимия. 17 (17): 3448–3453. Дои:10.1021 / bi00610a004. ISSN 0006-2960. PMID 28749.
- ^ Гонсалвес, Ж. Тьягу; Шафер, Саймон Т .; Гейдж, Фред Х. (2016). «Взрослый нейрогенез в гиппокампе: от стволовых клеток к поведению». Клетка. 167 (4): 897–914. Дои:10.1016 / j.cell.2016.10.021. PMID 27814520.
- ^ Чжао С., Тэн Э.М., Саммерс Р.Г. мл., Мин Г.Л., Гейдж ФХ (2006). «Четкие морфологические стадии созревания нейронов зубчатых гранул в гиппокампе взрослых мышей». J Neurosci. 26 (1): 3–11. Дои:10.1523 / jneurosci.3648-05.2006. ЧВК 6674324. PMID 16399667.
- ^ Шорс, Трейси Дж .; Miesegaes, Джордж; Бейлин, Анна; Чжао, Минжруй; Райдел, Трейси; Гулд, Элизабет (2001). «Нейрогенез у взрослого человека участвует в формировании следовых воспоминаний». Природа. 410 (6826): 372–376. Дои:10.1038/35066584. PMID 11268214. S2CID 4430850.
- ^ Денни, Кристина А .; Burghardt, Nesha S .; Schachter, Daniel M .; Хен, Рене; Дрю, Майкл Р. (2012-05-01). «Нейроны гиппокампа, рожденные взрослыми в возрасте от 4 до 6 недель, влияют на исследования, вызванные новизной, и контекстуальные условия страха». Гиппокамп. 22 (5): 1188–1201. Дои:10.1002 / hipo.20964. ISSN 1098-1063. ЧВК 3193906. PMID 21739523.
- ^ Plack, C.J .; Oxenham, A.J .; Дрга, В. (2006). "Журнал Neurosci". Журнал неврологии. 26 (34): 8767–8773. Дои:10.1523 / JNEUROSCI.1134-06.2006. ЧВК 1808348. PMID 16928865.
- ^ Дрю, Майкл Р .; Денни, Кристина А .; Хен, Рене (2010). «Прекращение нейрогенеза гиппокампа у взрослых мышей нарушает контекстуальное обусловливание страха в одном, но не в нескольких испытаниях». Поведенческая неврология. 124 (4): 446–454. Дои:10.1037 / a0020081. ЧВК 2925248. PMID 20695644.
- ^ Кеснер, Раймонд П. (2013-10-01). «Анализ функции зубчатой извилины». Поведенческие исследования мозга. SI: Сети памяти медиальной височной доли. 254: 1–7. Дои:10.1016 / j.bbr.2013.01.012. PMID 23348108. S2CID 5486456.
- ^ França, Thiago F.A .; Bitencourt, Alexandre M .; Maximilla, Naiana R .; Barros, Daniela M .; Монсеррат, Хосе М. (20.07.2017). «Нейрогенез гиппокампа и разделение паттернов: метаанализ поведенческих данных». Гиппокамп. 9. 27 (9): 937–950. Дои:10.1002 / hipo.22746. PMID 28597491. S2CID 205914396.
- ^ Сахай, Амар; Scobie, Kimberly N .; Хилл, Алексис С .; О'Кэрролл, Колин М .; Хейрбек, Мазен А .; Burghardt, Nesha S .; Fenton, André A .; Драновский, Алексей; Хен, Рене (2011). «Повышение нейрогенеза в гиппокампе у взрослых достаточно для улучшения разделения паттернов». Природа. 472 (7344): 466–470. Bibcode:2011Натура.472..466S. Дои:10.1038 / природа09817. ЧВК 3084370. PMID 21460835.
- ^ Clelland, C.D .; Choi, M .; Romberg, C .; Clemenson, G.D .; Fragniere, A .; Tyers, P .; Jessberger, S .; Саксида, Л. М .; Баркер, Р. А. (10 июля 2009 г.). «Функциональная роль нейрогенеза гиппокампа взрослых в пространственном разделении паттернов». Наука. 325 (5937): 210–213. Bibcode:2009Наука ... 325..210C. Дои:10.1126 / science.1173215. ISSN 1095-9203. ЧВК 2997634. PMID 19590004.
- ^ França, Thiago F.A .; Bitencourt, Alexandre M .; Maximilla, Naiana R .; Barros, Daniela M .; Монсеррат, Хосе М. (20.07.2017). «Нейрогенез гиппокампа и разделение паттернов: метаанализ поведенческих данных». Гиппокамп. 9. 27 (9): 937–950. Дои:10.1002 / hipo.22746. PMID 28597491. S2CID 205914396.
- ^ Хейрбек, Мазен А .; Дрю, Лиам Дж .; Burghardt, Nesha S .; Costantini, Daniel O .; Танненхольц, Линдси; Ahmari, Susanne E .; Цзэн, Хункуй; Fenton, André A .; Хен, Рене (2013-03-06). «Дифференциальный контроль обучения и тревожности вдоль дорсовентральной оси зубчатой извилины». Нейрон. 77 (5): 955–968. Дои:10.1016 / j.neuron.2012.12.038. ISSN 1097-4199. ЧВК 3595120. PMID 23473324.
- ^ Гулд Э, Кэмерон HA (1997). «Ранняя блокада рецепторов NMDA нарушает защитное поведение и увеличивает пролиферацию клеток в зубчатой извилине у развивающихся крыс». Behav. Неврологи. 111 (1): 49–56. Дои:10.1037/0735-7044.111.1.49. PMID 9109623.
- ^ Кемперманн Г., Вискотт Л., Гейдж Ф.Х. (апрель 2004 г.). «Функциональное значение взрослого нейрогенеза». Текущее мнение в нейробиологии. 14 (2): 186–91. Дои:10.1016 / j.conb.2004.03.001. PMID 15082323. S2CID 6241827.
- ^ а б G. Neves, G; С.Ф. Кук; Т.В. Блисс (2008). «Синаптическая пластичность, память и гиппокамп: нейросетевой подход к причинности». Обзоры природы Неврология. 9 (1): 65–75. Дои:10.1038 / №2303. PMID 18094707. S2CID 33104731.
- ^ Беккер С (2005). «Вычислительный принцип для обучения гиппокампа и нейрогенеза». Гиппокамп. 15 (6): 722–38. Дои:10.1002 / hipo.20095. PMID 15986407.
- ^ Вискотт Л., Раш М.Дж., Кемперманн Г. (2006). «Функциональная гипотеза нейрогенеза гиппокампа взрослых: предотвращение катастрофического вмешательства в зубчатую извилину». Гиппокамп. 16 (3): 329–43. CiteSeerX 10.1.1.408.8944. Дои:10.1002 / hipo.20167. PMID 16435309.
- ^ Аймон Дж. Б., Уайлс Дж., Гейдж Ф. Х. (июнь 2006 г.). «Возможная роль взрослого нейрогенеза в кодировании времени в новых воспоминаниях». Nat. Неврологи. 9 (6): 723–7. Дои:10.1038 / nn1707. PMID 16732202. S2CID 10586519.
- ^ Ковач К.А. (сентябрь 2020 г.). «Эпизодические воспоминания: как гиппокамп и энторинальные кольцевые аттракторы взаимодействуют для их создания?». Границы системной нейробиологии. 14: 68. Дои:10.3389 / fnsys.2020.559186.
- ^ Шорс Т.Дж., Таунсенд Д.А., Чжао М., Козоровицкий Ю., Гулд Э. (2002). «Нейрогенез может относиться к некоторым, но не ко всем типам гиппокампа-зависимого обучения». Гиппокамп. 12 (5): 578–84. Дои:10.1002 / hipo.10103. ЧВК 3289536. PMID 12440573.
- ^ Меши Д., Дрю М.Р., Сакси М. и др. (Июнь 2006 г.). «Нейрогенез гиппокампа не требуется для поведенческих эффектов обогащения окружающей среды». Nat. Неврологи. 9 (6): 729–31. Дои:10.1038 / nn1696. PMID 16648847. S2CID 11043203.
- ^ Gould, E .; Бейлин, А .; Tanapat, P .; Ривз, А .; Шорс, Т. Дж. (1999). «Обучение усиливает нейрогенез взрослых в формировании гиппокампа». Природа Неврология. 2 (3): 260–265. Дои:10.1038/6365. PMID 10195219. S2CID 12112156.
- ^ Донован, М. Х .; Яздани, У; Norris, R.D .; Игры, D; German, D.C .; Эйш, А. Дж. (2006). «Снижение нейрогенеза гиппокампа взрослых в модели мышей PDAPP болезни Альцгеймера». Журнал сравнительной неврологии. 495 (1): 70–83. Дои:10.1002 / cne.20840. PMID 16432899. S2CID 20559097.
- ^ Джин, К; Peel, A. L .; Мао, X. O .; Се, L; Cottrell, B.A .; Henshall, D.C .; Гринберг, Д. А. (2004). «Повышенный нейрогенез гиппокампа при болезни Альцгеймера». Труды Национальной академии наук. 101 (1): 343–7. Bibcode:2004PNAS..101..343J. Дои:10.1073 / pnas.2634794100. ЧВК 314187. PMID 14660786.
- ^ Foster, P. P .; Rosenblatt, K. P .; Кульиш, Р. О. (2011). «Когнитивная пластичность, вызванная упражнениями, последствия для легких когнитивных нарушений и болезни Альцгеймера». Границы неврологии. 2: 28. Дои:10.3389 / fneur.2011.00028. ЧВК 3092070. PMID 21602910.
- ^ Marx CE, Trost WT, Shampine LJ, et al. (Декабрь 2006 г.). «Нейростероид аллопрегнанолон снижается в префронтальной коре при болезни Альцгеймера». Биол. Психиатрия. 60 (12): 1287–94. Дои:10.1016 / j.biopsych.2006.06.017. PMID 16997284. S2CID 27813000.
- ^ Ван Дж. М., Сингх С., Лю Л., Ирвин Р. У., Чен С., Чунг Е. Дж., Томпсон Р. Ф., Бринтон Р. Д. (2010). «Аллопрегнанолон устраняет нейронный и когнитивный дефициты на мышиной модели болезни Альцгеймера». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 107 (14): 6498–6503. Bibcode:2010PNAS..107.6498W. Дои:10.1073 / pnas.1001422107. ЧВК 2851948. PMID 20231471.
- ^ Cissé, M; Чеклер, Ф (2014). «Рецепторы Eph: новые игроки в патогенезе болезни Альцгеймера». Нейробиология болезней. 73C: 137–149. Дои:10.1016 / j.nbd.2014.08.028. PMID 25193466. S2CID 28922292.
- ^ Mu, Y; Гейдж, Ф. Х. (2011). «Взрослый нейрогенез гиппокампа и его роль в болезни Альцгеймера». Молекулярная нейродегенерация. 6: 85. Дои:10.1186/1750-1326-6-85. ЧВК 3261815. PMID 22192775.
- ^ LeStrat, Y (май 2009 г.). «Роль генов, вовлеченных в нейропластичность и нейрогенез, в наблюдении за взаимодействием генов и окружающей среды (GxE) при шизофрении». Современная молекулярная медицина. 9 (4): 506–18. Дои:10.2174/156652409788167104. PMID 19519407.
- ^ Шрайбер, Руди; Ньюман-Танкреди, Адриан (апрель 2014 г.). «Улучшение познания при шизофрении с помощью нейролептиков, которые вызывают нейрогенез за счет активации рецептора 5-HT1A». Нейробиология обучения и памяти. 110: 72–80. Дои:10.1016 / j.nlm.2013.12.015. PMID 24423786. S2CID 28027825.
- ^ Рейф, Андреас; Шмитт, Анжелика; Фритцен, Сабрина; Леш, Клаус-Петер (27 апреля 2007 г.).«Нейрогенез и шизофрения: делящиеся нейроны в разделенном сознании?». Европейский архив психиатрии и клинической неврологии. 257 (5): 290–299. Дои:10.1007 / s00406-007-0733-3. PMID 17468935. S2CID 28750292.
- ^ Нумакава, Тадахиро; Одака, Харуки; Адачи, Наоки (2017). «Влияние глюкокортикоидов на нейрогенез». Исследование нейронной регенерации. 12 (7): 1028–1035. Дои:10.4103/1673-5374.211174. ЧВК 5558474. PMID 28852377.
- ^ а б c Jacobs, B.L .; Х. ван Прааг; Ф. Х. Гейдж (2000). «Депрессия и рождение и смерть клеток мозга». Американский ученый. 88 (4): 340. Bibcode:2000AmSci..88..340J. Дои:10.1511/2000.4.340.
- ^ Kandel, E. R .; Дж. Х. Шварц и Т. М. Джессел (2012-10-26). Принципы нейронологии (пятое изд.). ISBN 978-0071390118.
- ^ Национальный институт психического здоровья (2010 г.). «Лекарства для психического здоровья» (PDF).
- ^ Дэвис-Сала, М.Г .; Espósito, M. S .; Piatti, V.C .; Mongiat, L.A .; Тринчеро М. Ф. и Шиндер А. Ф. (25 мая 2011 г.). «Время созревания нейронов во взрослом гиппокампе зависит от активности локальной сети». Журнал неврологии. 31 (21): 7715–28. Дои:10.1523 / JNEUROSCI.1380-11.2011. ЧВК 3701257. PMID 21613484.
- ^ Перера Т.Д., Дворк А.Дж., Киган К.А., Тирумангалакуди Л., Липира С.М., Джойс Н., Ланге С., Хигли Д.Д., Росоклия Г., Хен Р., Сакейм А.А., Коплан Д.Д. (2011). «Необходимость нейрогенеза гиппокампа для терапевтического действия антидепрессантов у взрослых нечеловеческих приматов». PLOS ONE. 6 (4): e17600. Bibcode:2011PLoSO ... 617600P. Дои:10.1371 / journal.pone.0017600. ЧВК 3078107. PMID 21525974.
- ^ а б Schloesser RJ, Manji HK, Martinowich K (апрель 2009 г.). «Подавление нейрогенеза у взрослых приводит к усилению реакции системы гипоталамо-гипофиз-надпочечники». NeuroReport. 20 (6): 553–7. Дои:10.1097 / WNR.0b013e3283293e59. ЧВК 2693911. PMID 19322118.
- ^ а б Surget A, Tanti A, Leonardo ED и др. (Декабрь 2011 г.). «Антидепрессанты привлекают новые нейроны для улучшения регуляции стрессовой реакции». Молекулярная психиатрия. 16 (12): 1177–88. Дои:10.1038 / mp.2011.48. ЧВК 3223314. PMID 21537331.
- ^ Мальберг JE, Eisch AJ, Nestler EJ, Думан Р.С. (Декабрь 2000 г.). «Хроническое лечение антидепрессантами увеличивает нейрогенез в гиппокампе взрослых крыс». J. Neurosci. 20 (24): 9104–10. Дои:10.1523 / JNEUROSCI.20-24-09104.2000. ЧВК 6773038. PMID 11124987.
- ^ Манев Х., Уз Т., Смалхайзер Н. Р., Манев Р. (январь 2001 г.). «Антидепрессанты изменяют пролиферацию клеток в мозге взрослого человека in vivo и в культурах нейронов in vitro». Eur J Pharmacol. 411 (1–2): 67–70. Дои:10.1016 / S0014-2999 (00) 00904-3. PMID 11137860.
- ^ Santarelli L, Saxe M, Gross C и др. (Август 2003 г.). «Потребность гиппокампа в нейрогенезе для поведенческих эффектов антидепрессантов». Наука. 301 (5634): 805–9. Bibcode:2003Наука ... 301..805С. Дои:10.1126 / science.1083328. PMID 12907793. S2CID 9699898.
- ^ Брэдли, Джозеф (2015). Зависимость: от страдания к решению. Лас-Вегас, Невада: Breaux Press International. п. 173. ISBN 978-0-9854418-0-7.
- ^ Кастрен Э (март 2005 г.). «Это химия настроения?». Обзоры природы Неврология. 6 (3): 241–6. Дои:10.1038 / №1629. PMID 15738959. S2CID 34523310.
- ^ Vetencourt, J. F. M .; Продажа, А .; Viegi, A .; Baroncelli, L .; De Pasquale, R .; f. o'Leary, O .; Castren, E .; Маффеи, Л. (2008). «Антидепрессант флуоксетин восстанавливает пластичность зрительной коры взрослого». Наука. 320 (5874): 385–8. Bibcode:2008Sci ... 320..385M. Дои:10.1126 / наука.1150516. PMID 18420937. S2CID 39565465.
- ^ Миреску С., Петерс Дж. Д., Нойман Л., Гулд Е. (декабрь 2006 г.). «Недостаток сна подавляет нейрогенез взрослых в гиппокампе за счет повышения уровня глюкокортикоидов». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 103 (50): 19170–5. Bibcode:2006ПНАС..10319170М. Дои:10.1073 / pnas.0608644103. ЧВК 1748194. PMID 17135354.
- ^ Mirescu C .; Peters J. D .; Noiman L .; Гулд Э. (2006). «Недостаток сна подавляет нейрогенез взрослых в гиппокампе за счет повышения уровня глюкокортикоидов». Труды Национальной академии наук. 103 (50): 19170–19175. Bibcode:2006ПНАС..10319170М. Дои:10.1073 / pnas.0608644103. ЧВК 1748194. PMID 17135354.
- ^ а б c Arias-Carrión O, Freundlieb N, Oertel WH, Höglinger GU (октябрь 2007 г.). «Взрослый нейрогенез и болезнь Паркинсона». Мишени для лекарств от неврологического расстройства ЦНС. 6 (5): 326–35. Дои:10.2174/187152707783220875. PMID 18045161. Архивировано из оригинал 14 апреля 2013 г.
- ^ Фэллон Дж., Рид С., Киньяму Р. и др. (Декабрь 2000 г.). «Индукция in vivo массивной пролиферации, направленной миграции и дифференцировки нервных клеток в мозге взрослых млекопитающих». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 97 (26): 14686–91. Bibcode:2000PNAS ... 9714686F. Дои:10.1073 / pnas.97.26.14686. ЧВК 18979. PMID 11121069.
- ^ Ариас-Каррион О, Вердуго-Диас Л., Фериа-Веласко А. и др. (Октябрь 2004 г.). «Нейрогенез в субвентрикулярной зоне после стимуляции транскраниального магнитного поля и нигростриатальных поражений». J Neurosci Res. 78 (1): 16–28. Дои:10.1002 / jnr.20235. PMID 15372495.
- ^ Ариас-Каррион О., Эрнандес-Лопес С., Ибаньес-Сандовал О., Баргас Дж., Эрнандес-Крус А., Друкер-Колин Р. (ноябрь 2006 г.). «Нейрональные предшественники в субвентрикулярной зоне взрослых крыс дифференцируются в дофаминергические нейроны после поражения черной субстанции и трансплантации хромаффинных клеток». J Neurosci Res. 84 (7): 1425–37. Дои:10.1002 / jnr.21068. PMID 17006899.
- ^ а б Höglinger GU, Rizk P, Muriel MP и др. (Июль 2004 г.). «Истощение запасов дофамина снижает пролиферацию клеток-предшественников при болезни Паркинсона». Nat. Неврологи. 7 (7): 726–35. Дои:10.1038 / nn1265. PMID 15195095. S2CID 952173.
- ^ Нейрогенез в полосатом теле мозга взрослого человека
- ^ Го, Цинминь; Сайид, Икбал; Baronne, Lon M .; Хоффман, Стюарт У .; Гэннун, Рашида; Стейн, Дональд Г. (апрель 2006 г.). «Введение прогестерона модулирует экспрессию AQP4 и отек после черепно-мозговой травмы у самцов крыс». Экспериментальная неврология. 198 (2): 469–478. Дои:10.1016 / j.expneurol.2005.12.013. ISSN 0014-4886. PMID 16445913. S2CID 26143265.
- ^ Петроне, Эшли Б .; Гатсон, Джошуа В .; Симпкинс, Джеймс У .; Рид, Миранда Н. (25 мая 2014 г.). «Нефеминизирующие эстрогены: новая нейрозащитная терапия». Молекулярная и клеточная эндокринология. 389 (1–2): 40–47. Дои:10.1016 / j.mce.2013.12.017. ISSN 1872-8057. ЧВК 4040321. PMID 24424441.
- ^ Han, S .; Чжао, Б .; Пан, X .; Песня, З .; Liu, J .; Gong, Y .; Ван, М. (03.12.2015). «Вариант рецептора эстрогена ER-α36 участвует в нейрозащите эстрогена от окислительной токсичности». Неврология. 310: 224–241. Дои:10.1016 / j.neuroscience.2015.09.024. ISSN 1873-7544. PMID 26383254. S2CID 44289364.
- ^ Сингх, Сурджит; Хота, Дебасиш; Пракаш, Аджай; Хандуджа, Кришан Л .; Арора, Сунил К .; Чакрабарти, Амитава (январь 2010 г.). «Аллопрегнанолон, активный метаболит прогестерона, защищает от повреждения нейронов в модели припадков, вызванных пикротоксином, у мышей». Фармакология, биохимия и поведение. 94 (3): 416–422. Дои:10.1016 / j.pbb.2009.10.003. ISSN 1873-5177. PMID 19840816. S2CID 45591868.
- ^ Вебстер, Марк К .; Кули-Темм, Синтия А .; Барнетт, Джозеф Д .; Бах, Харрисон Б .; Вайннер, Джессика М .; Вебстер, Сара Э .; Линн, Синди Л. (27 марта 2017 г.). «Доказательства наличия BrdU-положительных нейронов сетчатки после применения агониста никотинового ацетилхолинового рецептора Alpha7». Неврология. 346: 437–446. Дои:10.1016 / j.neuroscience.2017.01.029. ISSN 1873-7544. ЧВК 5341387. PMID 28147247.
- ^ Drapeau, E .; Mayo, W .; Aurousseau, C .; Moal, M.L .; Пьяцца, П., Абрус, Д.Н. (2003). «Показатели пространственной памяти у старых крыс в водном лабиринте предсказывают уровни нейрогенеза в гиппокампе». PNAS. 100 (24): 14385–14390. Bibcode:2003ПНАС..10014385Д. Дои:10.1073 / пнас.2334169100. ЧВК 283601. PMID 14614143.
- ^ Marrone, D.F .; Рамирес-Амайя, В. и Барнс, К.А. (2012). «Нейроны, образующиеся в процессе старения, сохраняют способность к функциональной интеграции». Гиппокамп. 22 (5): 1134–1142. Дои:10.1002 / hipo.20959. ЧВК 3367380. PMID 21695743.
- ^ фон Болен и Хальбах O (2010). «Участие BDNF в возрастных изменениях в гиппокампе». Фронт старения Neurosci. 2. Дои:10.3389 / fnagi.2010.00036. ЧВК 2952461. PMID 20941325.
- ^ Praag H, Christie BR, Sejnowski TJ, Gage FH (1999). «Бег улучшает нейрогенез, обучение и долгосрочную потенцию у мышей». Proc Natl Acad Sci U S A. 96 (23): 13427–31. Bibcode:1999ПНАС ... 9613427В. Дои:10.1073 / пнас.96.23.13427. ЧВК 23964. PMID 10557337.
- ^ а б c Фермер Дж., Чжао Х, ван Прааг Х, Водтке К., Гейдж ФХ, Кристи Б.Р. (2004). «Влияние произвольных упражнений на синаптическую пластичность и экспрессию генов в зубчатой извилине взрослых самцов крыс Sprague-Dawley in vivo». Неврология. 124 (1): 71–9. Дои:10.1016 / j.neuroscience.2003.09.029. PMID 14960340. S2CID 2718669.
- ^ van Praag H; Christie BR; Sejnowski TJ; Gage FH (ноябрь 1999 г.). «Бег улучшает нейрогенез, обучение и долгосрочную потенцию у мышей». Proc Natl Acad Sci U S A. 96 (23): 13427–31. Bibcode:1999ПНАС ... 9613427В. Дои:10.1073 / пнас.96.23.13427. ЧВК 23964. PMID 10557337.
- ^ а б Карро, Э; Trejo, J. L .; Busiguina, S; Торрес-Алеман, I (2001). «Циркулирующий инсулиноподобный фактор роста I опосредует защитные эффекты физических упражнений от мозговых инсультов различной этиологии и анатомии». Журнал неврологии. 21 (15): 5678–84. Дои:10.1523 / JNEUROSCI.21-15-05678.2001. ЧВК 6762673. PMID 11466439.
- ^ «Взрослый нейрогенез». Мозговые брифинги. Общество неврологии. Июнь 2007 г.. Получено 2011-11-26.
- ^ Вэнь Цзян; Юнь Чжан; Лань Сяо; Джейми Ван Клемпут; Шао-Пин Цзи; Гуан Бай; Ся Чжан (01.11.2005). «Каннабиноиды способствуют нейрогенезу эмбрионального и взрослого гиппокампа и вызывают анксиолитические и антидепрессантные эффекты». Журнал клинических исследований. 115 (11): 3104–16. Дои:10.1172 / JCI25509. ЧВК 1253627. PMID 16224541. Получено 2011-03-02.
- ^ 11-19-2008 Исследование штата Огайо: ученые пришли к выводу, что марихуана снижает ухудшение памяти. OSU.edu
- ^ 29 ноября 2006 г. Исследование: марихуана может влиять на активацию нейронов. United Press International
- ^ Wolf, Susanne A .; Бик-Сандер, Аника; Фабель, Клаус; Леал-Галисия, Перла; Таубер, Свантье; Рамирес-Родригес, Херардо; Мюллер, Анке; Мельник, Андре; Уолтингер, Тим П .; Ульрих, Оливер; Кемперманн, Герд (2010). «Каннабиноидный рецептор CB1 опосредует исходную и вызванную активностью выживание новых нейронов в нейрогенезе гиппокампа взрослых». Сотовая связь и сигнализация. 8 (1): 12. Дои:10.1186 / 1478-811X-8-12. ЧВК 2898685. PMID 20565726.
- ^ Demirakca, T .; Sartorius, A .; Ende, G .; Meyer, N .; Welzel, H .; Skopp, G .; Mann, K .; Германн, Д. (2010). «Уменьшение серого вещества в гиппокампе потребителей каннабиса: возможные защитные эффекты каннабидиола». Наркотическая и алкогольная зависимость. 114 (2–3): 242–245. Дои:10.1016 / j.drugalcdep.2010.09.020. PMID 21050680.
- ^ Райт, М. Дж .; Vandewater, S.A .; Таффе, М.А. (2013). «Каннабидиол ослабляет дефицит зрительно-пространственной ассоциативной памяти, вызванный Δ9-тетрагидроканнабинолом». Британский журнал фармакологии. 170 (7): 1365–1373. Дои:10.1111 / bph.12199. ЧВК 3838683. PMID 23550724.
- ^ Morgan, C.J.A .; Schafer, G .; Freeman, T. P .; Курран, Х. В. (2010). «Влияние каннабидиола на острую память и психотомиметические эффекты курения каннабиса: натуралистическое исследование». Британский журнал психиатрии. 197 (4): 285–290. Дои:10.1192 / bjp.bp.110.077503. PMID 20884951.
- ^ а б Калабрезе, Эдвард Дж .; Рубио-Касильяс, Альберто (май 2018 г.). «Двухфазные эффекты ТГК на память и познание». Европейский журнал клинических исследований. 48 (5): e12920. Дои:10.1111 / eci.12920. PMID 29574698.
- ^ Сулиман, Нур Азуин; Тайб, Че Норма Мат; Моклас, Мохамад Арис Мохд; Басир, Руслиза (21 сентября 2017 г.). «Дельта-9-Тетрагидроканнабинол (∆9-THC) индуцируют нейрогенез и улучшают когнитивные способности самцов крыс Sprague Dawley». Исследования нейротоксичности. 33 (2): 402–411. Дои:10.1007 / s12640-017-9806-х. ЧВК 5766723. PMID 28933048.
- ^ Куккураццу, Бруна; Замберлетти, Эрика; Наззаро, Криштиану; Прини, Памела; Трюсел, Массимо; Grilli, Mariagrazia; Пароларо, Даниэла; Тонини, Рафаэлла; Рубино, Тициана (ноябрь 2018 г.). «Взрослые клеточные нейроадаптации, вызванные воздействием ТГК в подростковом возрасте у самок крыс, спасаются за счет усиления передачи сигналов анандамида». Международный журнал нейропсихофармакологии. 21 (11): 1014–1024. Дои:10.1093 / ijnp / pyy057. ЧВК 6209859. PMID 29982505.
- ^ Лазаров О., Робинсон Дж., Тан Ю.П., Хейрстон И.С., Корад-Мирникс З., Ли В.М., Херш Л.Б., Сапольский Р.М., Мирникс К., Сисодия С.С. (март 2005 г.). «Обогащение окружающей среды снижает уровни Abeta и отложение амилоида у трансгенных мышей». Клетка. 120 (5): 701–13. Дои:10.1016 / j.cell.2005.01.015. PMID 15766532. S2CID 16756661.
- ^ Van Praag, H .; Шуберт, Т .; Zhao, C .; Гейдж, Ф. (2005). «Упражнения улучшают обучаемость и нейрогенез гиппокампа у старых мышей». Журнал неврологии. 25 (38): 8680–8685. Дои:10.1523 / JNEUROSCI.1731-05.2005. ЧВК 1360197. PMID 16177036.
- ^ Van Praag, H .; Kempermann, G .; Гейдж, Ф. (1999). «Бег увеличивает пролиферацию клеток и нейрогенез в зубчатой извилине взрослых мышей». Природа Неврология. 2 (3): 266–270. Дои:10.1038/6368. PMID 10195220. S2CID 7170664.
- ^ Бьорнебекк А., Мате А.А., Брене С. (сентябрь 2005 г.). «Антидепрессивный эффект бега связан с повышенной пролиферацией клеток гиппокампа». Int J Нейропсихофармакол. 8 (3): 357–68. Дои:10.1017 / S1461145705005122. PMID 15769301.
- ^ Джин К., Ван Х, Се Л. и др. (Август 2006 г.). «Доказательства нейрогенеза, вызванного инсультом, в мозге человека». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 103 (35): 13198–202. Bibcode:2006PNAS..10313198J. Дои:10.1073 / pnas.0603512103. ЧВК 1559776. PMID 16924107.
- ^ Родительский JM; Elliott, RC; Удовольствие, SJ; Барбаро, Нью-Мексико; Левенштейн, DH (2006). «Аберрантный нейрогенез, вызванный судорогами при экспериментальной височной эпилепсии». Анналы неврологии. 59 (1): 81–91. Дои:10.1002 / ana.20699. HDL:2027.42/49280. PMID 16261566. S2CID 9655613.
- ^ Gerber J, Tauber SC, Armbrecht I, Schmidt H, Brück W, Nau R (2009). «Повышенная нейрональная пролиферация при бактериальном менингите человека». Неврология. 73 (13): 1026–32. Дои:10.1212 / WNL.0b013e3181b9c892. PMID 19786694. S2CID 26108905.
- ^ Шарма А., Валади Н., Миллер А. Х., Пирс Б. Д. (2002). «Взрослый нейрогенез в гиппокампе нарушается после неонатальной вирусной инфекции». Нейробиология болезней. 11 (2): 246–56. Дои:10.1006 / nbdi.2002.0531. PMID 12505418. S2CID 6405590.
- ^ Ли А.Л., Огл В.О., Сапольски Р.М. (апрель 2002 г.). «Стресс и депрессия: возможные связи с гибелью нейронов в гиппокампе». Биполярное расстройство. 4 (2): 117–28. Дои:10.1034 / j.1399-5618.2002.01144.x. PMID 12071509.
- ^ Шелин Ю.И., Гадо М.Х., Кремер Х.С. (август 2003 г.). «Нелеченная депрессия и потеря объема гиппокампа». Am J Psychiatry. 160 (8): 1516–8. Дои:10.1176 / appi.ajp.160.8.1516. PMID 12900317.
- ^ Jiang, W .; и другие. (2005). «Каннабиноиды способствуют нейрогенезу эмбрионального и взрослого гиппокампа и вызывают анксиолитические и антидепрессантные эффекты». Журнал клинических исследований. 115 (11): 3104–3116. Дои:10.1172 / JCI25509. ЧВК 1253627. PMID 16224541.
- ^ Орр, Анна Г .; Шарма, Ануп; Binder, Николаус Б .; Миллер, Эндрю Х .; Пирс, Брэдли Д. (2010). «Интерлейкин-1 опосредует долгосрочную потерю зубных гранул гиппокампа после послеродовой вирусной инфекции». Журнал молекулярной неврологии. 41 (1): 89–96. Дои:10.1007 / s12031-009-9293-5. PMID 19774496. S2CID 2427299.
- ^ Hu, X.L .; Wang, Y .; Шен, К. (2012). «Эпигенетический контроль выбора клеточной судьбы в нервных стволовых клетках». Белки и клетки. 3 (4): 278–290. Дои:10.1007 / s13238-012-2916-6. ЧВК 4729703. PMID 22549586.
- ^ Альтман, Дж. (1962). «Формируются ли новые нейроны в головном мозге взрослых млекопитающих?». Наука. 135 (3509): 1127–1128. Bibcode:1962Sci ... 135.1127A. Дои:10.1126 / science.135.3509.1127. PMID 13860748. S2CID 1606140.
- ^ Альтман, Дж. (1963). «Авторадиографические исследования пролиферации клеток мозга крыс и кошек». Анатомический рекорд. 145 (4): 573–591. Дои:10.1002 / ар.1091450409. PMID 14012334. S2CID 5340726.
- ^ Альтман, Дж. (1969). «Авторадиографические и гистологические исследования постнатального нейрогенеза. IV. Клеточная пролиферация и миграция в передней части переднего мозга, с особым упором на сохраняющийся нейрогенез в обонятельной луковице». Журнал сравнительной неврологии. 137 (4): 433–457. Дои:10.1002 / cne.901370404. PMID 5361244. S2CID 46728071.
- ^ Bayer, S.A .; Yackel, J. W .; Пури, П. С. (1982). «Нейроны в зернистом слое зубчатой извилины крысы существенно увеличиваются в ювенильном и взрослом возрасте». Наука. 216 (4548): 890–892. Bibcode:1982Наука ... 216..890B. Дои:10.1126 / science.7079742. PMID 7079742.
- ^ Байер, С. А. (1982). «Изменения в общем количестве зубчатых гранулярных клеток у молодых и взрослых крыс: коррелированное волюметрическое и авторадиографическое исследование с 3H-тимидином». Экспериментальное исследование мозга. Experimentelle Hirnforschung. Эксперименты Cerebrale. 46 (3): 315–323. Дои:10.1007 / bf00238626. PMID 7095040. S2CID 18663323.
- ^ Goldman SA, Nottebohm F (апрель 1983 г.). «Производство, миграция и дифференциация нейронов в ядре голосового управления головного мозга взрослой самки канарейки». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 80 (8): 2390–4. Bibcode:1983PNAS ... 80.2390G. Дои:10.1073 / pnas.80.8.2390. ЧВК 393826. PMID 6572982.
- ^ Reynolds, B.A .; Вайс, С. (март 1992 г.). «Генерация нейронов и астроцитов из изолированных клеток центральной нервной системы взрослых млекопитающих». Наука. 255 (5052): 1707–1710. Bibcode:1992Научный ... 255.1707R. Дои:10.1126 / science.1553558. ISSN 0036-8075. PMID 1553558.
- ^ Gage, F. H .; Ray, J .; Фишер, Л. Дж. (1995). «Выделение, характеристика и использование стволовых клеток ЦНС». Ежегодный обзор нейробиологии. 18: 159–92. Дои:10.1146 / annurev.ne.18.030195.001111. PMID 7605059.
- ^ Eriksson PS, Perfilieva E, Björk-Eriksson T, et al. (Ноябрь 1998 г.). «Нейрогенез в гиппокампе взрослого человека». Nat. Med. 4 (11): 1313–7. Дои:10.1038/3305. PMID 9809557.
- ^ Gould, E .; Ривз; Фаллах; Танапат; Валовой; Фукс (1999). «Нейрогенез гиппокампа у взрослых приматов Старого Света». Труды Национальной академии наук Соединенных Штатов Америки. 96 (9): 5263–5267. Bibcode:1999PNAS ... 96.5263G. Дои:10.1073 / пнас.96.9.5263. ЧВК 21852. PMID 10220454.
- ^ Понти, G .; Peretto, B .; Бонфанти, Л. (2008). Рех, Томас А. (ред.). «Генез нейрональных и глиальных предшественников в коре мозжечка периферических и взрослых кроликов». PLOS ONE. 3 (6): e2366. Bibcode:2008PLoSO ... 3.2366P. Дои:10.1371 / journal.pone.0002366. ЧВК 2396292. PMID 18523645.
- ^ Gould, E .; Ривз, А. Дж .; Грациано, М. С .; Гросс, К. Г. (1999). «Нейрогенез в неокортексе взрослых приматов». Наука. 286 (5439): 548–552. Дои:10.1126 / science.286.5439.548. PMID 10521353.
- ^ Чжао, М .; Мама, С .; Delfani, K .; Карлен, М .; Кэссиди, Р. М .; Johansson, C.B .; Brismar, H .; Шупляков, О .; Frisen, J .; Янсон, А. (2003). «Доказательства нейрогенеза в черной субстанции взрослых млекопитающих». Труды Национальной академии наук Соединенных Штатов Америки. 100 (13): 7925–7930. Bibcode:2003PNAS..100.7925Z. Дои:10.1073 / pnas.1131955100. ЧВК 164689. PMID 12792021.
- ^ Голень; Rafii, M. S .; Посадка, Б. Н .; Фэллон, Дж. Х. (1999). «Приблизительное удвоение числа нейронов в постнатальной коре головного мозга человека и в 35 конкретных цитоархитектурных областях от рождения до 72 месяцев». Детская патология и патология развития. 2 (3): 244–259. Дои:10.1007 / с100249900120. PMID 10191348. S2CID 23546857.
- ^ Ракич П. (февраль 2002 г.). «Взрослый нейрогенез у млекопитающих: кризис идентичности». J. Neurosci. 22 (3): 614–8. Дои:10.1523 / JNEUROSCI.22-03-00614.2002. ЧВК 6758501. PMID 11826088.
- ^ Песня, Хуан; Чжун, Чун; Bonaguidi, Michael A .; Sun, Джеральд Дж .; Хсу, Дерек; Гу, Ян; Мелетис, Константинос; Хуанг, З. Джош; Ге, Шаоюй; Ениколопов, Григорий; Дейссерот, Карл; Люшер, Бернхард; Christian, Kimberly M .; Мин, Го-ли; Песня, Хунцзюнь (2012). «Механизм нейронных цепей, регулирующих решение о судьбе покоящихся нервных стволовых клеток взрослых». Природа. 489 (7414): 150–154. Bibcode:2012Натура.489..150S. Дои:10.1038 / природа11306. ЧВК 3438284. PMID 22842902.
- Примечания
- Aimone JB, Jessberger S и Gage FH (2007) Взрослый нейрогенез. Scholarpedia, стр. 8739
- Гулд Э., Ривз А.Дж., Фаллах М., Танапат П., Гросс К.Г., Фукс Э. (апрель 1999 г.). «Нейрогенез гиппокампа у взрослых приматов Старого Света». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 96 (9): 5263–7. Bibcode:1999PNAS ... 96.5263G. Дои:10.1073 / пнас.96.9.5263. ЧВК 21852. PMID 10220454.
- Гулд Э., Ривз А.Дж., Грациано М.С., Гросс К.Г. (октябрь 1999 г.). «Нейрогенез в неокортексе взрослых приматов». Наука. 286 (5439): 548–52. Дои:10.1126 / science.286.5439.548. PMID 10521353.
- Гулд Э., Бейлин А., Танапат П., Ривз А., Шорс Т.Дж. (март 1999 г.). «Обучение усиливает нейрогенез взрослых в формировании гиппокампа». Nat. Неврологи. 2 (3): 260–5. Дои:10.1038/6365. PMID 10195219. S2CID 12112156.
- Могхадам К.С., Чен А., Хиткот, Р.Д. (август 2003 г.). «Установление судьбы вентральных клеток в спинном мозге». Dev. Dyn. 227 (4): 552–62. Дои:10.1002 / dvdy.10340. PMID 12889064.
- Ракич П. (январь 2002 г.). «Нейрогенез в неокортексе взрослых приматов: оценка доказательств». Обзоры природы Неврология. 3 (1): 65–71. Дои:10.1038 / nrn700. PMID 11823806. S2CID 15725675.
- Роллс, Э. Т. и Тревес, А. (1998). Нейронные сети и функции мозга. Оксфорд: ОУП. ISBN 0-19-852432-3.
- Santarelli L, Saxe M, Gross C и др. (Август 2003 г.). «Требование нейрогенеза гиппокампа для поведенческих эффектов антидепрессантов». Наука. 301 (5634): 805–9. Bibcode:2003Наука ... 301..805С. Дои:10.1126 / science.1083328. PMID 12907793. S2CID 9699898.
- Schloesser RJ, Manji HK, Martinowich K (апрель 2009 г.). «Подавление нейрогенеза у взрослых приводит к усилению реакции гипоталамо-гипофизарно-надпочечниковой системы».. NeuroReport. 20 (6): 553–7. Дои:10.1097 / WNR.0b013e3283293e59. ЧВК 2693911. PMID 19322118.
- Shankle WR, Rafii MS, Landing BH, Fallon JH (1999). «Приблизительное удвоение числа нейронов в постнатальной коре головного мозга человека и в 35 конкретных цитоархитектонических областях от рождения до 72 месяцев». Детская патология и патология развития. 2 (3): 244–259. Дои:10.1007 / с100249900120. PMID 10191348. S2CID 23546857.
- Чжао М., Мама С., Дельфани К. и др. (Июнь 2003 г.). «Доказательства нейрогенеза в черной субстанции взрослых млекопитающих». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 100 (13): 7925–30. Bibcode:2003PNAS..100.7925Z. Дои:10.1073 / pnas.1131955100. ЧВК 164689. PMID 12792021.
- Специальный выпуск Философские труды B по стволовым клеткам и ремонту мозга. Некоторые статьи находятся в свободном доступе.[постоянная мертвая ссылка ]
внешняя ссылка
- Краткое введение в нейрогенез из Колледж Уэллсли
- Комплексный сайт о нейрогенезе из Колледж Лафайет
- Ранняя литература по нейрогенезу взрослых
- Нейрогенез во взрослом мозге - Фред Х. Гейдж и Генриетта ван Прааг
- «Нейрогенез и болезнь Паркинсона»
- Статья из Scholarpedia о нейрогенезе у взрослых
- "Тенденции в неврологии, 10 октября 2001 г. (Майкл С. Каплан, доктор медицины)
- New York Times: исследования показывают, что в мозге вырастают новые клетки
- Майкл Спектер: переосмысление мозга - Как песни канареек нарушают фундаментальный принцип науки
- Эксперимент по нейрогенезу - Серия статей о нейрогенезе взрослого человека
- Журнал Seed: переосмысление себя - Исторический фон в области нейрогенеза и значение этого исследования.
- BBC Radio 4: воспоминания - Используйте это или потеряйте
- PBS: Изменение вашего мнения - Развивайте свой собственный мозг
- Стальные доли: аэробные упражнения способствуют нейрогенезу., New York Times, 19 августа 2007 г.
 СМИ, связанные с нейрогенез в Wikimedia Commons
СМИ, связанные с нейрогенез в Wikimedia Commons