Ретроназальный запах - Википедия - Retronasal smell
Ретроназальный запах, ретроназальный обоняние, это способность воспринимать вкус размеры продуктов и напитков. Ретроназальный запах - это сенсорный способ создания аромата. Лучше всего его описать как комбинацию традиционного запаха (ортоназальный запах) и вкусовых ощущений.[1] Ретроназальный запах создает аромат из-за молекул запаха в пище или напитках, проходящих через носовые проходы во время жевания. Когда люди используют термин «запах», они обычно имеют в виду «ортоназальный запах» или восприятие молекул запаха, которые попадают прямо через нос и поднимаются по носовым ходам. Ретроназальный запах имеет решающее значение для ощущения вкус продуктов и напитков. Аромат должен контрастировать с вкус, который относится к пяти конкретным параметрам: (1) милая, (2) соленый, (3) горький, (4) кислый, и (5) умами. Восприятие чего-либо, выходящего за рамки этих пяти измерений, например, различения вкуса яблока и груши, требует ощущения ретроназального запаха.
История
С эволюционной точки зрения запах долгое время считался менее важным. смысл для людей, особенно по сравнению со зрением. Похоже, что зрение доминирует над восприятием стимулов человеком, но теперь исследователи утверждают, что запахи очень информативны для человека, хотя и менее очевидны. Перед его смертью в 1826 году французский гастроном Brillat-Savarin опубликовал свою книгу, Физиология вкуса; Или «Медитации о трансцендентальной гастрономии: теоретическая, историческая и практическая работа»., в котором он впервые упоминает о важности запаха в «комбинированном чувстве вкуса». Он определяет вкус с точки зрения пяти параметров вкуса в дополнение к аромату, создаваемому носовым аппаратом.[1] Эйвери Гилберт, в его книге Нос знает, рассматривает работу Генри Т. Финка, американского философа конца 1800-х годов, опубликовавшего новаторское эссе под названием «Гастрономическая ценность запахов». Флинк называл аромат «вторым способом обоняния», и многие последующие научные исследования в начале 1900-х были сосредоточены на попытке разбить параметры запаха на основные категории, что оказалось слишком сложным из-за огромного количества и сложности запахов.
Ценители еды и повара все больше используют новое понимание роли запаха во вкусе. Пищевые ученые Николас Курти и Эрве Это расширил физиологию аромата и его важность в кулинарное искусство. В 2006 году он опубликовал свою книгу, Молекулярная гастрономия: изучение науки о вкусе, в котором он исследует физические механизмы, которые вызывают восприятие аромата. Курти и это повлияли на других, таких как Гарольд МакГи, чья книга 1984 года, О еде и кулинарии: наука и знания о кухне, был тщательно пересмотрен в 2004 году и остается ключевым справочником в научном понимании приготовления пищи. Его книгу описал телеведущий. Альтон Браун как «Розеттский камень кулинарного мира». Такой прорыв в понимании механизмов ощущения вкуса различных продуктов, вероятно, продолжит вдохновлять кулинаров на создание новых комбинаций и рецептов.
Сегодня один из самых активных диетологов, Павел Розин был первым, кто успешно определил роль ретроназального запаха в аромате. В 1982 году он объяснил, что запах - это «двойное чувство», и провел четкое различие между ретроназальным запахом и ортоназальным запахом.[1] Розин описывает ортоназальный запах как «вдох», а ретроназальный запах как «выдох». В 1982 году он разработал эксперимент, в котором обучал участников точно распознавать запахи ортоназально, прежде чем вводить их в заднюю часть рта, после чего процент успеха резко упал, демонстрируя, что запах действует через два различных механизма.[2] Его любимым примером этой двойственности является сыр Лимбургер, который известен своим отталкивающим действием для носа, но приятным для рта.
Первоначально опубликовано в 2012 г. Нейрогастрономия к Гордон М. Шеперд дает обзор того, как запах воспринимается людьми. Книга содержит подробный обзор того, как ретроназальный запах в сочетании со вкусом создает аромат. После тщательного изучения нейронной основы идентификации, распознавания и предпочтения определенных вкусов, Шеперд исследует потенциальные политические и социальные последствия более глубокого понимания вкусового восприятия, такие как причины ожирения и проблемы потери чувствительности к запахам в пожилом возрасте.[1]
Обзор пути запаха
Чтобы лучше понять этот механизм, ниже представлена простая разбивка запахов. Когда люди жуют, летучие ароматические соединения проходят через носоглотку и обонятельные рецепторы.
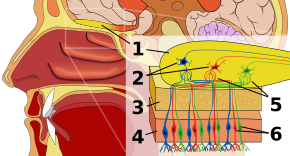
Обонятельный эпителий
Первая остановка в обонятельная система это обонятельный эпителий или ткань, лежащая на крыше носовой полости, в которой находятся рецепторы запаха. Рецепторы запаха биполярные нейроны которые связывают одоранты из воздуха и собираются в обонятельном нерве перед передачей аксонов к дендритам митральных клеток в обонятельной луковице.[3] Сенсорные рецепторы во рту и носу поляризованы в состоянии покоя и деполяризуются в ответ на некоторые изменения в окружающей среде, например, на контакт с молекулами запаха. Молекулы запаха, состоящие из углеводородных цепей с функциональные группы, связываются с сенсорными рецепторами в носу и рту. Свойства функциональных групп включают: (1) длину углеродной цепи, (2) концевую группу, которая согласуется с различиями, связанными с разными запахами, (3) боковую группу, (4) хиральность, (5) форму и (6) размер. . Когда молекулы запаха связываются с сенсорными рецепторами, они делают это в соответствии с этими свойствами. Каждая обонятельная клетка имеет рецептор одного типа, но этот рецептор может быть «широко настроен», и молекулы запаха дополнительно взаимодействуют на уровне рецептора, а это означает, что в некоторых случаях молекула запаха сама по себе может не связываться с рецептором, но в При наличии другой молекулы запаха оригинал будет связываться и, таким образом, создавать ощущение запаха только в присутствии второй молекулы.[4]
Обонятельная луковица
В обонятельной луковице молекулы запаха нанесены на карту пространственно. Эти пространственные представления известны как «образы запахов».[1] Разрешения на пространственное представление боковое торможение, или усиление контраста и сжатие усиления. Повышение контрастности чувствительно к изменениям и выделяет стимулы в мозгу, которые меняются, а не в состоянии покоя. Сжатие усиления повышает чувствительность к стимулам низкой интенсивности и снижает чувствительность к стимулам высокой интенсивности. Обонятельная луковица, еще находящаяся на начальной стадии своего понимания исследователями, отличает запах от других чувств, потому что он отмечает отклонение сенсорного пути от того, что характерно для всех других чувств. А именно, вся не обонятельная сенсорная информация проходит через таламус после уровня рецепторов, но тот факт, что информация об запахе вместо этого входит в свою собственную специализированную область, может указывать на примитивную историю запаха и / или особый тип обработки информации об запахе на ее пути к коре головного мозга. В обонятельной луковице находятся клубочки или клеточные узлы, на которых расположены тысячи рецепторов одного и того же типа, помимо митральные клетки, сходятся. Такая организация позволяет кратко представить огромный объем информации, не требуя одинаково большого количества типов рецепторов. Полученная комбинация информации об запахе дублируется изображением запаха на уровне обонятельной луковицы.[5]
Визуализация обонятельной луковицы
2DG метод
В 1977 году биохимик Лу Соколов, Сеймур Кети, и Флойд Э. Блум разработали способ картирования активности мозга путем отслеживания метаболизма кислорода мозгом крысы. Нервным клеткам необходим кислород и глюкоза для получения энергии. 2-дезоксиглюкоза (2DG) - это радиоактивный изотоп глюкозы, который можно отслеживать в мозге, поскольку он оставляет след в клетке, где он обычно метаболизировался бы для получения энергии, если бы это была глюкоза. После стимуляции определенной области клеток можно нарезать рентгеновские снимки, чтобы выявить, какие клетки были активными, особенно в синаптических соединениях.[6]
Функциональная магнитно-резонансная томография (фМРТ) также может использоваться для измерения метаболизма запаха. Этот метод не является терминальным, как 2-дезоксиглюкоза метод, так что одно животное можно измерить с множеством запахов, а полученные изображения можно будет сравнить.
Метод зеленого флуоресцентного белка
Наконец, зеленый флуоресцентный белок Метод генетически конструирует мышей для экспрессии белка в активных нейронах, а затем камеру можно поместить в череп мыши для измерения активности.[7]
Результаты
Эти методы показывают, в первую очередь, что информация об запахе в обонятельной луковице организована пространственно. Сходные молекулярные паттерны приводят к сходным паттернам активации в отношении клубочков, и клубочки, которые расположены ближе друг к другу, кодируют сходные особенности информации запаха.[1][6][7]
Обонятельная кора
Трехслойная обонятельная кора, содержащая пирамидные клетки это следующий ориентир на пути запаха. Одна пирамидная клетка получает информацию от множества митральных клеток из обонятельной луковицы, создавая ранее организованный паттерн клубочков, распределенный в обонятельной коре. Эта дисперсия информации митральных клеток позволяет создавать самовозбуждающие обратные связи, латеральное возбуждение, а также само- и латеральное торможение. Эти процессы способствуют Hebbian обучение, названный в честь Дональд О. Хебб, и часто упрощается поговоркой «нейроны, которые возбуждаются вместе, соединяются вместе». Долгосрочное потенцирование, нейронный механизм обучения Хебба, позволяет формировать память на уровне пирамидных клеток. Таким образом, обучение Хебба - это, по сути, феномен, с помощью которого обонятельная кора «запоминает» выход комбинаций молекул запаха и позволяет распознавать ранее воспринимаемые комбинации быстрее, чем новые, путем сопоставления их с сохраненными входными данными. Полученные запахи, которые раньше назывались образами запахов, хранятся в обонятельной коре для распознавания, теперь называются объектами запаха.[5] Таким образом, опыт улучшает отношение сигнал / шум, поскольку ранее ощущаемый объект запаха может быть более легко различим на фоне большего фонового шума.[8]
Орбитофронтальная кора
В орбитофронтальная кора (OFC) - это конечный пункт назначения информации об запахе, и именно здесь возникает сознательное восприятие запаха. Информация об запахе поступает сразу после прохождения через обонятельную кору, что указывает на отличие от другой сенсорной информации, которая сначала проходит через таламус. OFC расположен дорсальнее префронтальная кора, позволяя обонятельной информации напрямую поступать в префронтальную кору или основную зону мозга, принимающую решения. Есть только три типа нейронов, которые проходят через обонятельные молекулы, прежде чем попасть в OFC: обонятельные рецепторные клетки в обонятельном эпителии, митральные клетки и обонятельные пирамидные нейроны.[1]
На уровне OFC создаются ассоциации с другими областями мозга, включая информацию изо рта (соматосенсорное ощущение ), эмоциональный ввод (миндалины), визуальная информация и оценочная информация (префронтальная кора). OFC отвечает за выборочную настройку запаха, слияние сенсорных доменов и гедонистическую оценку запахов.
Домашние доказательства роли запаха в аромате
Опыт употребления любимых блюд при простуде часто разочаровывает. Это связано с тем, что заложенность носа блокирует носовые проходы, через которые входят и выходят молекулы воздуха и ароматизатора, таким образом временно снижая способность ретроназального запаха.
Еще один способ изолировать ретроназальный запах - использовать «тест на защемление носа». Когда едят, зажимая ноздри закрытыми, кажется, что вкус пищи рассеивается, а именно потому, что путь выхода воздуха из носа, создающего образ аромата, перекрывается.
Спекулятивное эволюционное значение
Более глубокое понимание роли ретроназального запаха в аромате привело многих к переосмыслению эволюционного значения запаха у людей. Чтобы развеять представление о том, что зрение полностью превосходит людей и выше приматы обонянию, Гордон М. Шеперд противопоставляет анатомия человеческого носа на собачий.[1] У собак рецепторы запаха расположены в задней части носовой полости. У них есть уникальный орган, похожий на картридж, который служит воздушным фильтром. Во время спокойного дыхания этот картридж обычно направляет поток воздуха, но во время активного обоняния скорость направления информации увеличивается, позволяя собаке обнюхивать в шесть-восемь раз быстрее, чем человек.[1]
Это говорит о том, что клыки приспособлены к более сильному ортоназальному запаху. В отличие от людей, кажется, что они обладают превосходной ретроназальной способностью обоняния. В двуногий осанка человека снижает потребность в патроне, который функционирует у собак, чтобы в основном очищать поступающий воздух. Короткий носоглотка для ретроназального запаха у людей - это то, что позволяет летучим веществам из продуктов и напитков перемещаться изо рта к рецепторам запаха в полости носа. Что остается менее очевидным, так это тот факт, что собаки по-прежнему обладают сильной способностью различать пищу.
Другие предположения включают идею о том, что короткий путь ото рта к носовой полости возник в результате отбора в результате бега на длинные дистанции, когда люди мигрировали из Африки 2 миллиона лет назад.[9] Идея состоит в том, что более короткий носовой аппарат поможет уравновесить голову и облегчить бег на длинные дистанции. Либерман ссылается на другие эволюционные изменения, которые могли возникнуть в результате отбора к бегу, такие как более широкие суставные хрящи и более длинные кости в ногах.[10]
Рекомендации
- ^ а б c d е ж грамм час я Шеперд, Гордон М. (2012). Нейрогастрономия. Нью-Йорк: издательство Колумбийского университета. ISBN 9780231159104.
- ^ Розин, Павел (1982-07-01). ""Смешение вкуса и запаха "и двойственность обонятельного чувства". Восприятие и психофизика. 31 (4): 397–401. Дои:10.3758 / BF03202667. ISSN 0031-5117. PMID 7110896.
- ^ Моран, Д. Т .; Rowley, J.C .; Jafek, B.W .; Ловелл, М.А. (1982-10-01). «Тонкая структура обонятельной слизистой оболочки человека». Журнал нейроцитологии. 11 (5): 721–746. Дои:10.1007 / bf01153516. ISSN 0300-4864. PMID 7143026. S2CID 25263022.
- ^ Duchamp-Viret, P .; Chaput, M. A .; Дюшан, А. (1999-06-25). "Свойства отклика запаха нейронов обонятельного рецептора крысы". Наука. 284 (5423): 2171–2174. Дои:10.1126 / science.284.5423.2171. ISSN 0036-8075. PMID 10381881.
- ^ а б Эволюция нервных систем. Академическая пресса. 2016-11-23. п. 17. ISBN 9780128040966.
- ^ а б Стюарт, Уильям Б.; Кауэр, Джон С .; Шеперд, Гордон М. (1979-06-15). «Функциональная организация обонятельной луковицы крысы проанализирована методом 2-дезоксиглюкозы». Журнал сравнительной неврологии. 185 (4): 715–734. Дои:10.1002 / cne.901850407. ISSN 1096-9861. PMID 447878. S2CID 41466291.
- ^ а б Brazelton, T. R .; Росси, Ф. М .; Keshet, G.I .; Блау, Х. М. (2000-12-01). «От костного мозга к мозгу: выражение нейрональных фенотипов у взрослых мышей». Наука. 290 (5497): 1775–1779. Дои:10.1126 / наука.290.5497.1775. ISSN 0036-8075. PMID 11099418. S2CID 16216476.
- ^ Уилсон, Дональд А .; Стивенсон, Ричард Дж. (11 мая 2006 г.). Обучение обонянию: обонятельное восприятие от нейробиологии к поведению. JHU Press. ISBN 9780801883682.
- ^ Либерман, Дэниел Э. и Деннис М. Брамбл. 2007. Эволюция марафонского бега: Возможности человека. Спортивная медицина 37 (4-5): 288-290.
- ^ Либерман, Даниэль. Эволюция головы человека. Кембридж, Массачусетс: Белкнап из Гарвардского университета, 2011. Печать.