Обонятельный рецептор нейрона - Olfactory receptor neuron
| Обонятельный рецептор нейрона | |
|---|---|
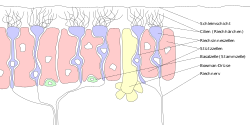 Этикетки на немецком языке. "Зеллен" = "ячейка "," riech "=" запах "," Riechnerv "= обонятельный нерв, "cillien" = реснички. | |
| подробности | |
| Система | Запах |
| Расположение | обонятельный эпителий в нос |
| Форма | Биполярный сенсорный рецептор |
| Функция | Обнаруживать следы химических веществ во вдыхаемом воздухе (Чувство обоняния ) |
| Нейротрансмиттер | Глутамат[1] |
| Пресинаптические связи | Никто |
| Постсинаптические связи | Обонятельная луковица |
| Идентификаторы | |
| MeSH | D018034 |
| НейроЛекс МНЕ БЫ | nifext_116 |
| TH | H3.11.07.0.01003 |
| Анатомические термины нейроанатомии | |
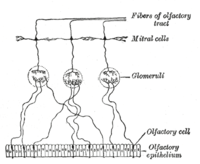

An нейрон обонятельного рецептора (ORN), также называемый обонятельный сенсорный нейрон (OSN), является сенсорный нейрон в пределах обонятельная система.[2]
Структура
У людей от 10 до 20 миллионов нейронов обонятельных рецепторов.[3] В позвоночные, ORN биполярные нейроны с участием дендриты обращены к внешней поверхности решетчатой пластинки с аксонами, которые проходят через решетчатые отверстия терминальным концом у обонятельных луковиц. ORN расположены в обонятельный эпителий в полости носа. Тела ORN распределены между всеми тремя стратифицированный слои обонятельного эпителия.[4]
Многие крошечные волосы реснички выступают из обонятельных рецепторных клеток дендрит в слизь покрывающие поверхность обонятельного эпителия. Поверхность этих ресничек покрыта обонятельные рецепторы, тип Рецептор, связанный с G-белком. Каждая обонятельная рецепторная клетка выражает только один тип обонятельных рецепторов (OR), но многие отдельные клетки обонятельных рецепторов экспрессируют OR, которые связывают один и тот же набор запахов. Аксоны обонятельных рецепторных клеток, которые экспрессируют одно и то же ИЛИ, сходятся, образуя клубочки в обонятельная луковица.[5]
Функция
ОР, которые расположены на мембранах ресничек, были классифицированы как сложный тип лиганд-управляемый метаботропные каналы.[6] Существует около 1000 различных генов, кодирующих OR, что делает их самым большим семейством генов. Одорант растворяется в слизи обонятельного эпителия, а затем связывается с OR. OR могут связываться с различными молекулами запаха с различным сродством. Различие в сродстве вызывает различия в моделях активации, что приводит к уникальным профилям одорантов.[7][8] Активированный OR, в свою очередь, активирует внутриклеточный G-белок, GOLF (GNAL ), аденилатциклаза и производство циклический AMP (cAMP) открывается ионные каналы в клеточная мембрана, что привело к притоку натрий и кальций ионов в клетку, и отток хлористый ионы. Этот приток положительных ионов и отток отрицательных ионов заставляет нейрон деполяризоваться, создавая потенциал действия.
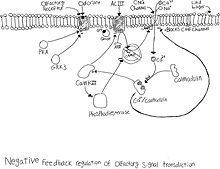
Десенсибилизация
Нейрон обонятельного рецептора быстро реагирует на отрицательную обратную связь при деполяризация. Когда нейрон деполяризуется, Ионный канал КПГ открыто, позволяя натрий и кальций ворваться в камеру. Приток кальция запускает каскад событий внутри клетки. Кальций сначала связывается с кальмодулином с образованием CaM. Затем CaM свяжется с каналом CNG и закроет его, остановив приток натрия и кальция.[9] CaMKII активируется присутствием СаМ, который фосфорилирует ACIII и снижает выработку цАМФ.[10] CaMKII также активирует фосфодиэстераза, который затем будет гидролизовать цАМФ.[11] Эффект этой отрицательной обратной связи препятствует дальнейшей активации нейрона, когда вводится другая молекула запаха.
Количество различимых запахов
Широко разрекламированное исследование показало, что люди могут обнаруживать более триллиона различных запахов.[12] Однако это открытие было оспорено. Критики утверждали, что методология, использованная для оценки, была в корне ошибочной, показывая, что применение того же аргумента для более понятных сенсорных модальностей, таких как зрение или слух, приводит к неправильным выводам.[13] Другие исследователи также показали, что результат чрезвычайно чувствителен к точным деталям расчета, с небольшими вариациями, изменяющими результат на десятки порядков величины, возможно, вплоть до нескольких тысяч.[14] Авторы оригинального исследования утверждали, что их оценка верна, пока предполагается, что пространство запаха достаточно многомерно.[15]
Другие животные
Эта секция нуждается в расширении. Вы можете помочь добавляя к этому. (Февраль 2018 г.) |
Смотрите также
использованная литература
- ^ Беркович, Д. А .; Trombley, P.Q .; Шеперд, Г. М. (1994). «Доказательства глутамата как нейромедиатора обонятельных рецепторных клеток». Журнал нейрофизиологии. 71 (6): 2557–61. Дои:10.1152 / jn.1994.71.6.2557. PMID 7931535.
- ^ Vermeulen, A; Роспарс, Дж. П. (1998). «Дендритная интеграция в обонятельных сенсорных нейронах: устойчивый анализ того, как структура нейрона и нейронная среда влияют на кодирование интенсивности запаха». Журнал вычислительной неврологии. 5 (3): 243–66. Дои:10.1023 / А: 1008826827728. PMID 9663551.
- ^ Саладин, Кеннет (2012). Анатомия и физиология: единство формы и функции (6-е изд.). Макгроу-Хилл. п. 593. ISBN 978-0073378251.
- ^ Каннингем, AM; Manis, P.B .; Reed, R.R .; Роннетт, Г.В. (1999). «Нейроны обонятельных рецепторов существуют как отдельные подклассы незрелых и зрелых клеток в первичной культуре». Неврология. 93 (4): 1301–12. Дои:10.1016 / s0306-4522 (99) 00193-1. PMID 10501454.
- ^ МакИвен, Д. П. (2008). «Обонятельные реснички: наша прямая нейронная связь с внешним миром». Curr. Верхний. Dev. Биол. Актуальные темы биологии развития. 85: 333–370. Дои:10.1016 / S0070-2153 (08) 00812-0. ISBN 9780123744531. PMID 19147011.
- ^ Тухара, Казусигэ (2009). "Обонятельный рецепторный комплекс насекомых функционирует как лиганд-зависимый ионотропный канал". Летопись Нью-Йоркской академии наук. 1170 (1): 177–80. Bibcode:2009НЯСА1170..177Т. Дои:10.1111 / j.1749-6632.2009.03935.x. PMID 19686133.
- ^ Bieri, S .; Монастырская, К; Шиллинг, Б. (2004). «Профилирование нейронов обонятельных рецепторов с использованием запахов сандалового дерева». Химические чувства. 29 (6): 483–7. Дои:10.1093 / chemse / bjh050. PMID 15269120.
- ^ Фань, Джинхонг; Нгаи, Джон (2001). «Начало экспрессии гена пахучих рецепторов во время регенерации обонятельных сенсорных нейронов». Биология развития. 229 (1): 119–27. Дои:10.1006 / dbio.2000.9972. PMID 11133158.
- ^ Брэдли, Дж; Reuter, D; Фрингс, S (2001). «Облегчение опосредованной кальмодулином адаптации запаха с помощью субъединиц цАМФ-управляемых каналов». Наука. 294 (5549): 2176–2178. Bibcode:2001Sci ... 294.2176B. Дои:10.1126 / science.1063415. PMID 11739960.
- ^ Вэй, Дж; Чжао, Аризона; Чан, GC; Бейкер, LP; Импей, S; Beavo, JA; Буря, Д.Р. (1998). «Фосфорилирование и ингибирование обонятельной аденилилциклазы CaM-киназой II в нейронах: механизм ослабления обонятельных сигналов». Нейрон. 21 (3): 495–504. Дои:10.1016 / s0896-6273 (00) 80561-9. PMID 9768837.
- ^ Ян, С; Чжао, Аризона; Bentley, JK; Loughney, K; Фергюсон, К; Биво, Дж. А. (1995). «Молекулярное клонирование и характеристика кальмодулин-зависимой фосфодиэстеразы, обогащенной обонятельными сенсорными нейронами». Proc Natl Acad Sci USA. 92 (21): 9677–9681. Bibcode:1995PNAS ... 92.9677Y. Дои:10.1073 / pnas.92.21.9677. ЧВК 40865. PMID 7568196.
- ^ Бушдид, C .; Magnasco, M.O .; Vosshall, L.B .; Келлер, А. (2014). «Люди могут различать более 1 триллиона обонятельных стимулов». Наука. 343 (6177): 1370–2. Bibcode:2014Научный ... 343.1370B. Дои:10.1126 / science.1249168. ЧВК 4483192. PMID 24653035.
- ^ Мейстер, Маркус (2015). «О размерности запахового пространства». eLife. 4: e07865. Дои:10.7554 / eLife.07865. ЧВК 4491593. PMID 26151672.
- ^ Геркин, Ричард С .; Кастро, Джейсон Б. (2015). «Число обонятельных стимулов, которые люди могут различать, до сих пор неизвестно». eLife. 4: e08127. Дои:10.7554 / eLife.08127. ЧВК 4491703. PMID 26151673.
- ^ Magnasco, Marcelo O .; Келлер, Андреас; Фосхолл, Лесли Б. (2015). «О размерности обонятельного пространства». Дои:10.1101/022103. Цитировать журнал требует
| журнал =(Помогите)
внешние ссылки
| Авторитетный контроль |
|---|