Активная зона - Active zone
| Активная зона | |
|---|---|
 Схема типичного синапса центральной нервной системы. Белки активной зоны представлены темно-коричневыми пирамидами на верхнем конце нейрона. | |
| Подробности | |
| Идентификаторы | |
| латинский | Zona Activa |
| TH | H2.00.06.2.00012 |
| Анатомические термины микроанатомии | |
В активная зона или же синаптическая активная зона - термин, впервые использованный Куто и Пеко-Дехавассинином в 1970 году для определения места расположения нейротрансмиттер релиз. Два нейроны установить непосредственный контакт через структуры, называемые синапсы позволяя им общаться друг с другом. Как показано на диаграмме рядом, синапс состоит из пресинаптического бутона одного нейрона, который хранит пузырьки содержащий нейротрансмиттер (вверху на картинке) и второй постсинаптический нейрон, который несет рецепторы для нейромедиатора (внизу), вместе с промежутком между ними, называемым синаптической щелью (с молекулами синаптической адгезии, SAM, удерживающими их вместе[1]). Когда потенциал действия достигает пресинаптического бутона, содержимое везикул попадает в синаптическую щель, и высвобожденный нейромедиатор перемещается через щель к постсинаптическому нейрону (нижняя структура на рисунке) и активирует рецепторы на постсинаптической мембране.
Активная зона - это область в пресинаптическом бутоне, которая опосредует высвобождение нейротрансмиттера и состоит из пресинаптической мембраны и плотного скопления белков, называемых цитоматриксом в активной зоне (CAZ). Под электронным микроскопом CAZ представляет собой темную (электронно-плотную) область рядом с мембраной. Белки внутри CAZ связывают синаптические пузырьки с пресинаптической мембраной и опосредуют синаптические слияние пузырьков, тем самым позволяя нейротрансмиттеру высвобождаться надежно и быстро при достижении потенциала действия.
Функция
Функция активной зоны - гарантировать, что нейротрансмиттеры может надежно высвобождаться в определенном месте нейрона и высвобождаться только тогда, когда нейрон запускает потенциал действия.[2]Как потенциал действия распространяется вниз по аксону, он достигает конца аксона, называемого пресинаптическим бутоном. В пресинаптическом бутоне потенциал действия активирует кальциевые каналы (VDCC), которые вызывают локальный приток кальция. Увеличение кальция обнаруживается белками в активной зоне и заставляет везикулы, содержащие нейротрансмиттер, сливаться с мембраной. Это слияние везикул с мембраной высвобождает нейротрансмиттеры в синаптическую щель (пространство между пресинаптическим бутоном и постсинаптической мембраной). Затем нейротрансмиттеры диффундируют через щель и связываются с лигандные ионные каналы и Рецепторы, сопряженные с G-белком на постсинаптической мембране. Связывание нейромедиаторов с постсинаптическими рецепторами затем вызывает изменение постсинаптического нейрона. Процесс высвобождения нейротрансмиттеров и связывания с постсинаптическими рецепторами, вызывающий изменение постсинаптического нейрона, называется нейротрансмиссией.
Структура
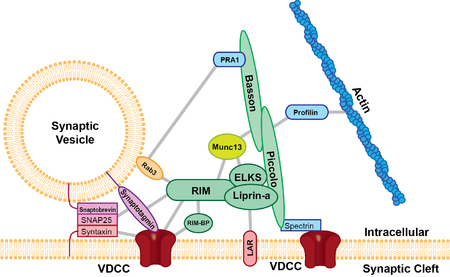
Активная зона присутствует во всех химические синапсы изучен до сих пор и присутствует у всех видов животных. Активные зоны, исследованные до сих пор, имеют по крайней мере две общие черты: все они содержат белковый плотный материал, который выступает из мембраны и связывает синаптические везикулы рядом с мембраной, и они имеют длинные нитевидные выступы, берущие начало на мембране и оканчивающиеся на везикулах немного дальше от нее. пресинаптическая мембрана. Плотные белковые выступы различаются по размеру и форме в зависимости от типа исследуемого синапса. Одним из ярких примеров плотных выступов является ленточный синапс (см. Ниже), который содержит «ленту» из плотного белкового материала, окруженного ореолом синаптических везикул и простирающегося перпендикулярно пресинаптической мембране, и его длина может достигать 500 нм.[3] Глутаматный синапс содержит более мелкие пирамидоподобные структуры, которые отходят от мембраны примерно на 50 нм.[4] Нервно-мышечный синапс содержит два ряда везикул с длинной белковой полосой между ними, которая соединяется с регулярно расположенными горизонтальными ребрами, идущими перпендикулярно полосе и параллельно мембране. Затем эти ребра соединяются с везикулами, каждая из которых расположена над выступом в мембране (предположительно, с кальциевым каналом).[5] Предыдущие исследования показали, что активная зона глутаматергический нейроны содержали очень регулярный массив плотного белкового материала в форме пирамиды, что указывало на то, что эти пирамиды были связаны нитями. Эта структура напоминала геометрическую решетку, в которой везикулы направлялись в отверстия решетки.[4] Эта привлекательная модель была поставлена под сомнение в результате недавних экспериментов. Недавние данные показывают, что глутаматергическая активная зона действительно содержит выступы плотного белкового материала, но эти выступы не были регулярными и содержали длинные волокна, выступающие в цитоплазму примерно на 80 нм.[6]
Есть по крайней мере пять основных белков каркаса, которые обогащены в активной зоне; UNC13B / Munc13, RIMS1 (Rab3-взаимодействующая молекула), Фагот, Пикколо / акзонин, ELKS, и липрины-α. Считается, что эти каркасные белки являются составными частями плотных пирамидоподобных структур активной зоны и, как полагают, приводят синаптические везикулы в непосредственную близость к пресинаптической мембране и кальциевым каналам. Белок ELKS связывается с клеточная адгезия белок β-нейрексин, и другие белки в составе комплекса, такие как Piccolo и Bassoon.[7] β-нейрексин затем связывается с молекулой клеточной адгезии, нейролигин расположен на постсинаптической мембране. Затем нейролигин взаимодействует с белками, которые связываются с постсинаптическими рецепторами. Белковые взаимодействия, подобные тем, которые наблюдаются между Piccolo / ELKS / β-нейрексином / нейролигином, гарантируют, что механизмы, которые опосредуют слияние пузырьков находится в непосредственной близости к кальциевым каналам, и это слияние пузырьков находится рядом с постсинаптическими рецепторами. Это слияние везикул в непосредственной близости от постсинаптических рецепторов обеспечивает небольшую задержку между активацией постсинаптических рецепторов и высвобождением нейротрансмиттеров.
Механизм высвобождения нейротрансмиттера

Высвобождение нейротрансмиттера достигается за счет слияния везикулы нейротрансмиттеров к пресинаптической мембране. Хотя детали этого механизма все еще изучаются, существует консенсус по некоторым деталям процесса. Известно, что слияние синаптических везикул с пресинаптической мембраной требует местного увеличения кальция.[9] от всего лишь одного, тесно связанных кальциевых каналов[10] и образование высокостабильных SNARE комплексы. Одна преобладающая модель слияния синаптических везикул заключается в том, что образование комплекса SNARE катализируется белками активной зоны, такими как Munc18, Munc13 и RIM. Считается, что образование этого комплекса «запускает» везикулу, чтобы она была готова к слиянию везикул и высвобождению нейромедиатора (см. Ниже: высвобождаемый пул). После того, как везикула загрунтована, тогда комплексин связывается с комплексом SNARE, это называется суперпраймированным. Везикулы с суперпраймированием находятся в легко высвобождаемом пуле (см. Ниже) и готовы к быстрому высвобождению. Прибытие потенциала действия открывает управляемые напряжением кальциевые каналы рядом с комплексом SNARE / комплексин. Затем кальций связывается, чтобы изменить конформацию синаптотагмин. Это изменение конформации позволяет синаптотагмину затем вытеснять комплексин, связываться с комплексом SNARE и связываться с мембраной-мишенью. Когда синаптотагмин связывается как с комплексом SNARE, так и с мембраной, это вызывает механическое воздействие на мембрану, заставляя мембрану везикул и пресинаптическую мембрану слиться. Это слияние открывает мембранную пору, которая высвобождает нейромедиатор. Пора увеличивается в размерах до тех пор, пока вся мембрана везикул не становится неотличимой от пресинаптической мембраны.[11][12][13]
Цикл синаптических везикул

Пресинаптический бутон имеет эффективно организованный процесс слияния везикул с пресинаптической мембраной для высвобождения нейротрансмиттеров и регенерации везикул нейромедиатора. Этот процесс, называемый циклом синаптических пузырьков, поддерживает количество пузырьков в пресинаптическом бутоне и позволяет синаптическому окончанию быть автономной единицей. Цикл начинается с (1) регион аппарат Гольджи отщипывается, образуя синаптический пузырек, и этот пузырек транспортируется к синаптическому окончанию. На терминале (2) пузырек заполнен нейромедиатором. (3) Везикула транспортируется в активную зону и стыкуется в непосредственной близости от плазматической мембраны. (4) Во время потенциала действия везикула сливается с мембраной, высвобождает нейромедиатор и позволяет мембранным белкам, ранее находившимся на везикуле, диффундировать в периактивную зону. (5) В периактивной зоне мембранные белки изолированы и эндоцитозированный формирование клатрин покрытый оболочкой пузырек. (6) Затем везикула наполняется нейротрансмиттером и затем транспортируется обратно в активную зону.
Механизм эндоцитоза медленнее, чем у экзоцитоз механизм. Это означает, что при интенсивной активности везикула в терминале может истощиться и больше не будет доступна для высвобождения. Чтобы предотвратить истощение синаптических пузырьков, повышение уровня кальция во время интенсивной активности может активировать кальциневрин который дефосфорилат белки, участвующие в клатрин-опосредованном эндоцитозе.[14]
Пузыри везикул
Синапс содержит по крайней мере два кластера синаптических везикул: легко высвобождаемый пул и резервный пул. Легко высвобождаемый пул расположен в активной зоне и напрямую связан с пресинаптической мембраной, в то время как резервный пул сгруппирован цитоскелетом и не связан напрямую с активной зоной.
Выдвижной бассейн
Высвобождаемый пул расположен в активной зоне и связан непосредственно с пресинаптической мембраной. Он стабилизируется белками в активной зоне и связывается с пресинаптической мембраной посредством SNARE белки. Эти везикулы готовы к высвобождению с помощью единственного потенциала действия и пополняются пузырьками из резервного пула. Выполняемый пул иногда подразделяется на выпускаемый пул и выпускаемый пул.
Резервный пул
Резервный пул не связан напрямую с активной зоной. Повышение пресинаптической концентрации кальция активирует кальций-кальмодулин-зависимая протеинкиназа (CaMK). CaMK фосфорилирует белок, синапсин, который обеспечивает кластеризацию везикул резервного пула и прикрепление к цитоскелету. Фосфорилирование синапсина мобилизует пузырьки в резервном пуле и позволяет им мигрировать в активную зону и пополнять легко высвобождаемый пул.[15][16]
Периактивная зона
Периактивная зона окружает активную зону и является местом эндоцитоза пресинаптического терминала. В периактивной зоне белки каркаса, такие как интерсектин 1 рекрутировать белки, которые опосредуют эндоцитоз, такие как динамин, клатрин и эндофилин.[17] В Дрозофилия гомолог интерсектина, Dap160, расположен в периактивной зоне нервно-мышечного соединения, а мутантный Dap160 истощает синаптические пузырьки во время высокочастотной стимуляции.[18]
Активная зона ленточного синапса
Ленточный синапс - это особый тип синапса, обнаруженный в сенсорные нейроны Такие как фоторецепторные клетки, биполярные клетки сетчатки, и волосковые клетки. Ленточные синапсы содержат плотную белковую структуру, которая связывает множество везикул, перпендикулярных пресинаптической мембране. В электронная микрофотография он выглядит как ленточная структура, перпендикулярная мембране. В отличие от «традиционных» синапсов, ленточные синапсы могут поддерживать постепенное высвобождение пузырьков. Другими словами, чем более деполяризован нейрон, тем выше скорость слияния везикул. Активная зона ленточного синапса разделена на две области: дугообразную плотность и ленту. Архиформная плотность является местом слияния везикул, а лента хранит высвобождаемый пул везикул. Ленточная структура состоит в основном из белка RIBEYE, примерно 64–69% объема ленты, и связана с аркообразной плотностью с помощью таких белков, как фагот.[19]
Белки
| Протеин | Структура / Функция |
| Структурные белки | |
| Пикколо | |
| Фагот | |
| Диски | |
| ЛОСЬ (ERC или CAST) | |
| КАСКА | |
| Мята | |
| Липрин-альфа-1 | |
| Стыковка и заливка | |
| Munc-13 | |
| Munc-18 | |
| SNAREs | |
| SNAP25 | |
| ВАМП2 | |
| синтаксин | Расположен на синаптической мембране и связывается с SNAP-25 и синаптобревином, опосредуя слияние везикул. |
| Цитоскелетные белки | |
| Актин | |
| Тубулин | |
| миозин Множественные молекулы миозина II создают силу в скелетных мышцах за счет механизма силового удара, подпитываемого энергией, высвобождаемой при гидролизе АТФ. | |
| спектрин | |
| β-катенин | |
| Кальциевый канал | |
| Напряжение зависимый кальциевый канал (VDCC) | Обеспечивает быстрый приток кальция во время потенциала действия. |
Измерение высвобождения нейромедиатора

Высвобождение нейротрансмиттера можно измерить, определив амплитуду постсинаптический потенциал после запуска потенциала действия в пресинаптическом нейроне. Измерение высвобождения нейротрансмиттера таким способом может быть проблематичным, потому что влияние постсинаптического нейрона на то же количество высвобожденного нейротрансмиттера может со временем измениться. Другой способ - измерить слияние везикул с пресинаптической мембраной непосредственно с помощью патч пипетка. Клеточную мембрану можно рассматривать как конденсатор в том, что положительные и отрицательные ионы накапливаются с обеих сторон мембраны. Чем больше площадь мембраны, тем больше ионов необходимо для удержания мембраны при определенном потенциале. В электрофизиологии это означает, что введение тока в терминал потребует меньше времени для зарядки мембраны до заданного потенциала до слияния везикул, чем после слияния везикул. Измеряется время зарядки мембраны до потенциала и сопротивление мембраны, и с этими значениями емкость мембраны может быть рассчитана по уравнению Тау / Сопротивление = Емкость. С помощью этого метода исследователи могут измерять высвобождение синаптических везикул напрямую, измеряя увеличение мембранной емкости пресинаптического терминала.[20]
Смотрите также
Рекомендации
- ^ Мисслер М, Зюдхоф ТК, Бидерер Т (2012). «Адгезия синаптических клеток». Холодная весна Харб Perspect Biol. 4 (4): a005694. Дои:10.1101 / cshperspect.a005694. ЧВК 3312681. PMID 22278667.
- ^ Крейг С. Гарнер и Кан Шен. Структура и функция активных зон позвоночных и беспозвоночных. Структура и функциональная организация синапса. Эд: Йоханнес Ад и Майкл Элерс. Спрингер, 2008.
- ^ Чжай Р. Грейс; Беллен Хьюго Дж. (2004). «Архитектура активной зоны в пресинаптическом нервном окончании». Физиология. 19 (5): 262–270. Дои:10.1152 / Physiol.00014.2004. PMID 15381754. S2CID 9609266.
- ^ а б Филлипс GR; и другие. (2001). «Пресинаптическая сеть частиц: ультраструктура, состав, растворение и воссоздание». Нейрон. 32 (1): 63–77. Дои:10.1016 / s0896-6273 (01) 00450-0. PMID 11604139. S2CID 5996555.
- ^ Mark L .; и другие. "Харлоу и другие. Архитектура материала активной зоны у лягушки. нервномышечное соединение". Природа. 409: 2001.
- ^ Сиксоу; и другие. (2007). «Трехмерная архитектура пресинаптической терминальной цитоматрицы». Журнал неврологии. 27 (26): 6868–6877. Дои:10.1523 / jneurosci.1773-07.2007. ЧВК 6672225. PMID 17596435.
- ^ Зив, Гарнер (2004). «Клеточные и молекулярные механизмы пресинаптической сборки». Клеточные и молекулярные механизмы пресинаптической сборки. 5 (5): 385–399. Дои:10.1038 / номер 1370. PMID 15100721. S2CID 21516580.
- ^ Георгиев, Данко Д .; Джеймс Ф. Глейзбрук (2007). «Субнейронная обработка информации уединенными волнами и случайными процессами». В Лышевском, Сергей Эдуард (ред.). Справочник по нано- и молекулярной электронике. Серия нано- и микротехники. CRC Press. С. 17-1–17-41. Дои:10.1201/9781315221670-17. ISBN 978-0-8493-8528-5.
- ^ Хайдельбергер; и другие. (1994). «Кальциевая зависимость скорости экзоцитоза в синаптическом окончании». Природа. 371 (6497): 513–515. Bibcode:1994Натура.371..513H. Дои:10.1038 / 371513a0. PMID 7935764. S2CID 4316464.
- ^ Стэнли Э. Ф. (1993). «Отдельные кальциевые каналы и высвобождение ацетилхолина на пресинаптическом нервном окончании». Нейрон. 11 (6): 1007–1011. Дои:10.1016 / 0896-6273 (93) 90214-с. PMID 8274272. S2CID 7311805.
- ^ Атасой и Кавалали. Машины высвобождения нейротрансмиттеров: компоненты нейронального комплекса SNARE и их функции. Структурно-функциональная организация синапса Ад и Элерс (ред.) 2008
- ^ Pang Z .; Судхоф Т. (2010). "Клеточная биология экзоцитоза, запускаемого Ca2 +". Текущее мнение в области клеточной биологии. 22 (4): 496–505. Дои:10.1016 / j.ceb.2010.05.001. ЧВК 2963628. PMID 20561775.
- ^ Carr C .; Мансон М. (2007). «Действие команды тегов в синапсе». EMBO отчеты. 8 (9): 834–838. Дои:10.1038 / sj.embor.7401051. ЧВК 1973957. PMID 17767192.
- ^ Юнг Наджа; Хаук Фолькер (2007). «Клатрин-опосредованный эндоцитоз в синапсах». Трафик. 8 (9): 1129–1136. Дои:10.1111 / j.1600-0854.2007.00595.x. PMID 17547698. S2CID 11320827.
- ^ Пинг Чи; Пол Грингард; Тимоти Райан (10 апреля 2003 г.). «Мобилизация синаптических пузырьков регулируется различными путями фосфорилирования синапсина I с разной частотой». Нейрон. 38 (1): 69–78. Дои:10.1016 / S0896-6273 (03) 00151-X. PMID 12691665. S2CID 17405359.
- ^ Cesca et al. (2010) Синапсины: ключевые участники функции и пластичности синапсов. Прогресс нейробиологии. Vol. 91. 313-348.
- ^ Дергай; и другие. (2010). «Интерсектин 1 образует комплексы с SGIP1 и Reps1 в ямках, покрытых клатрином». Сообщения о биохимических и биофизических исследованиях. 402 (2): 408–413. Дои:10.1016 / j.bbrc.2010.10.045. PMID 20946875.
- ^ Мари; и другие. (2004). «Dap160 / Intersectin Scaffolds Periactive Zone для достижения высокоточного эндоцитоза и нормального синаптического роста». Нейрон. 43 (2): 207–219. Дои:10.1016 / j.neuron.2004.07.001. PMID 15260957. S2CID 16296285.
- ^ Джордж Дзанацци и Гэри Мэтьюз. Молекулярная архитектура ленточных пресинаптических терминалов. Мол Нейробиол (2009) 39: 130-148.
- ^ Герсдорф Х. и Мэтьюз Г. (1994) Динамика слияния синаптических везикул и восстановления мембран в синаптических окончаниях. Природа. Том 367. 735-739
| Авторитетный контроль |
|---|