Ласточка - Tree swallow
| Ласточка | |
|---|---|
 | |
| В заповеднике Страуд, недалеко от Западный Честер, Пенсильвания, НАС | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Авес |
| Заказ: | Воробьиные |
| Семья: | Hirundinidae |
| Род: | Тахицинета |
| Разновидность: | Т. биколор |
| Биномиальное имя | |
| Tachycineta bicolor (Vieillot, 1808) | |
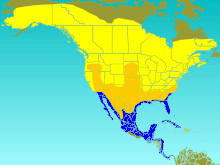 | |
| Диапазон Т. биколор Разведение дачника Посетитель миграции Зимний гость | |
| Синонимы | |
Хирундо биколор Вией, 1808 г. | |

В ласточка (Tachycineta bicolor) это перелетная птица семьи Hirundinidae. Найдено в Америка, ласточка была впервые описана в 1807 г. французским орнитолог Луи Виейо в качестве Хирундо биколор. С тех пор он был перемещен в свой нынешний род,Тахицинета, внутри которого обсуждается его филогенетическое размещение. Древесная ласточка имеет глянцевый сине-зеленый цвет. верхняя часть, за исключением черноватых крыльев и хвоста, и белого нижние части. Клюв черный, глаза темно-коричневые, а ноги и ступни светло-коричневые. Самка обычно тусклее самца, а у самки-первокурсницы верхняя часть тела в основном коричневая, с некоторыми голубыми перьями. У молодых особей коричневый верх и грудь серо-коричневого цвета. Древесная ласточка гнездится в США и Канаде. Зимует вдоль южного побережья США к югу, вдоль побережья Мексиканского залива, до Панамы и северо-западного побережья Южной Америки, а также в Вест-Индии.
Гнездится ласточка изолированными парами или отдельными группами, как в естественных, так и в искусственных полостях. Размножение может начаться уже в начале мая, хотя эта дата наступает раньше из-за изменение климата, а может закончиться уже в июле. Эта птица вообще социально моногамный (хотя около 8% мужчин полигинный ), с высоким уровнем внепарное отцовство. Это может принести пользу самцу, но, поскольку самка контролирует совокупление, отсутствие решения о том, как такое поведение приносит пользу самкам, ставит в тупик высокий уровень отцовства вне пары. Самка насиживает схватить от двух до восьми (но обычно от четырех до семи) чистых белых яиц в течение примерно 14-15 дней. Цыплята вылупляются немного асинхронно, что позволяет самке определять приоритетность кормления в периоды нехватки корма. Обычно они оперяются примерно через 18–22 дня после вылупления. Древесную ласточку иногда считают модельный организм, из-за большого количества проведенных исследований.
Антенна насекомоядное животное, дерево глотает корма как в одиночку, так и группами, питаясь в основном насекомые, в добавление к моллюски, пауки, и фрукты. Птенцы, как и взрослые особи, в основном питаются насекомыми, которых кормили им оба пола. Этот глотать уязвима для паразитов, но на птенцах они мало повреждают. Воздействие болезни может усиливаться по мере взросления ласточки, так как с возрастом некоторые части иммунной системы снижаются. Приобретенный Т-клетка -опосредованный иммунитет, например, с возрастом снижается, в то время как как врожденный, так и приобретенный гуморальный иммунитет не. Древесная ласточка из-за большого ареала и стабильной популяции считается наименьшие опасения посредством Международный союз охраны природы. В США он защищен Закон 1918 года о договоре о перелетных птицах, а в Канаде Закон о Конвенции о перелетных птицах. На эту ласточку негативно влияет деятельность человека, например, вырубка леса; закисленные озера может вынудить размножающуюся древесную ласточку преодолевать большие расстояния в поисках богатой кальцием пищи для своих птенцов.
Таксономия и этимология
Древесная ласточка описывалась какХирундо биколор к Луи Пьер Виейо в егоHistoire naturelle des oiseaux de l'Amerique Septentrionale, опубликовано в 1807 г.[2] Затем он был помещен в его текущий родТахицинета когда Жан Кабанис установил его в 1850 году.[3] В 1878 г. Эллиотт Коуз предложил поставить ласточку, по крайней мере, в собственное подрод, Иридопрокне, по оперению вместе с Белокрылая ласточка, Чилийская ласточка, белая ласточка, и мангровая ласточка.[4] К 1882 году он расширил его до полного рода.[5] Некоторые авторы продолжали использовать эту классификацию с добавлением Тумбес глотать; однако генетические данные подтверждают существование единственного рода,Тахицинета.[6] Древесную ласточку еще называют белобрюхой ласточкой из-за ее белого низа.[7]
Общепринятое название рода от Древнегреческий тахикинетос, "быстро движется" идвухцветный является латинский и означает «двухцветный».[8] Другое название рода,Иридопрокне, происходит из Греческий Ирис, что означает радуга, и Procne, фигура, которая якобы превратилась в ласточку.[5]
Как именно ласточка связана с другими членамиТахицинета не решена. В исследованиях на основе митохондриальная ДНК, он был размещен базальный (что означает, что это было первое ответвление в дереве видов) в Североамериканско-Карибском регионе клады состоящий из фиолетово-зеленая ласточка, золотая ласточка, и Багамская ласточка.[9][10] Хотя митохондриальная ДНК считается лучшим индикатором эволюционных изменений, поскольку она быстро эволюционирует, анализы, основанные на ней, могут пострадать, потому что она унаследована только от матери, что делает ее хуже ядерная ДНК из нескольких места при представлении филогении целой группы.[11] Исследование, основанное на такой ядерной ДНК, поместило ласточку в самое нижнее положение внутри. Тахицинета в целом (как сестринская группа к остальным представителям рода).[12]
Описание

Древесная ласточка имеет длину от 12 до 14 см (от 4,7 до 5,5 дюйма) и вес от 17 до 25,5 г (от 0,60 до 0,90 унции). Размах крыльев колеблется от 11,8 до 13,8 дюйма (30-35 см).[13] Самец преимущественно глянцевый сине-зеленый. верхняя часть, крылья и хвост черноватые. В нижние части[14] и щеки белые,[7] Хотя подкрылье кроющие серо-коричневые.[14] Клюв черный, глаза темно-коричневые, а ноги и ступни светло-коричневые.[7] Самка имеет более тусклый цвет, чем самец, и иногда имеет коричневый лоб.[14] Второй год[примечание 1] у самок также верхняя часть тела коричневая с переменным количеством синих перьев; некоторые самки третьего года жизни также сохраняют часть этого недозрелого оперения.[15] Согласно исследованию 1987 года, это, вероятно, позволяет более молодой самке исследовать места гнездования, поскольку постоянный самец обычно менее агрессивен по отношению к младшей самке.[16] Исследование 2013 года показало, что постоянные женщины были менее агрессивны по отношению к моделям второго года обучения, когда они были представлены отдельно от моделей старшего возраста. Неизвестно, почему самка в конечном итоге меняет свое оперение до взрослого; это может позволить самцам оценивать качество самок, как пары помощник ассортативно по яркости оперения.[17] Молодую древовидную ласточку можно отличить по коричневой верхней части и серо-коричневой груди.[14]
Голос

Песня ласточки состоит из трех частей: щебетание, нытье и бульканье. Эти разделы могут повторяться или опускаться, и все они могут стоять отдельно. Первый, как щебечущий зов (иногда разделенный на контактный и настойчивый), издается самкой во время совокупления.[18] и у обоих полов, чтобы стимулировать птенцов к попрошайничеству[19] или (в некоторых популяциях) когда их партнерша покидает или входит в следующую полость. Свист, обычно состоящий из сдвига частоты вниз, за которым следует сдвиг вверх, можно назвать одним лишь тревожным призывом.[18] иногда делается в ответ на определенных хищников.[20] Бульканье, которое появляется в конце песни, обычно произносится дважды. Вероятно, это связано с парными связями. Призыв болтовни используется для рекламы сайтов-гнезд (причина, по которой он также известен как "рекламный звонок-гнездо"), а также передается вторгшимся сородичам. Короткий высокий призыв к подчинению иногда произносится после агрессивной встречи с другой ласточкой дерева. Когда вы находитесь в физическом состоянии или испытываете боль, может быть подан сигнал бедствия. Самец часто издает тикающий (или грубый) агрессивный призыв во время совокупления, и оба пола используют его в конце моббинг-ныряния. Сигнал тревоги подается в ответ на хищников и других злоумышленников,[18] и может побуждать более старых птенцов приседать и прекращать попрошайничество, когда рядом хищник.[21]
Связь между родителями и потомством может быть нарушена шумом, создаваемым человеком. Например, исследование 2014 года показало, что выводки, для которых использовался белый шум, с меньшей вероятностью приседали или прекращали попрошайничать в ответ на сигнал тревоги. Родители не изменили своих требований, чтобы компенсировать ущерб, вероятно, увеличив риск нападения хищников.[22] Шум также может мешать реакции родителей на попрошайничество, но это может быть уравновешено более громкими криками, которые птенцы издают, когда им подвергают. Однако усиление попрошайничества может быть неэффективным или дорогостоящим из-за более высокого уровня шума.[23]
Распространение и среда обитания
Древесная ласточка размножается в Северной Америке. Его ареал простирается до северо-центральной Аляски и до линия дерева в Канаде. Он встречается так далеко на юг, как Теннесси в восточной части своего ареала, Калифорния и Нью-Мексико на западе и Канзас в центре. Иногда он гнездится дальше на юг в США,[7] и бродяги иногда встречаются в Полярный круг, северный Тихий океан, Гренландия, и Европа.[14] Ареал зимовки простирается от Калифорнии и юго-западной Аризоны на западе и юго-восточной Вирджинии на востоке и юге вдоль побережья Мексиканского залива до побережья Мексиканского залива. Вест-Индии, Панама и северо-западное побережье Южной Америки.[7] Во время миграции эта ласточка часто пользуется стоянками, проводя в них в среднем 57 дней в течение осени. Чтобы добраться до своего зимовья, он часто использует один из трех пролетных путей: западный пролетный путь, к западу от скалистые горы; Центральный пролет между Скалистыми горами и Великие озера, простираясь на юг в Восточную Мексику; и восточный пролетный путь от Великих озер на восток.[24] Когда ласточка возвращается в гнездо, она обычно не меняет места гнездования.[25]
Среда размножения этой птицы в основном на открытых и лесных участках, особенно у воды.[7] Он устраивается на ночлег каждую ночь в период отсутствия размножения, предпочитая отдыхать в трость или же тростниковые заросли над водой, но также встречается над сушей, деревьями и проводами. Обычно места гнездования находятся на расстоянии от 100 до 150 километров (от 62 до 93 миль).[26]
Поведение

Из-за большого количества исследований древовидной ласточки и того, как она охотно размножается в гнездах, биолог Джейсон Джонс рекомендовал считать ее одним из модельный организм.[27] Эта ласточка агрессивна в период размножения, но вне нее общительна, собирая стаи, иногда насчитывающие тысячи птиц.[7]
Разведение

Гнезда ласточек в конструкциях с уже существующими отверстиями, как естественными, так и искусственными. Когда-то они были найдены только в лесных районах, но строительство гнездовых ящиков позволило этой птице распространиться на открытые места обитания.[7] Эта ласточка обычно гнездится на том месте, где она разводилась годом ранее; только около 14% самок и 4% самцов расселяются для размножения на новом участке в год. Большинство из них не уезжают далеко, обычно гнездятся на участках менее чем в 10 километрах (6,2 мили) от их первоначальных земель. На рассредоточение влияет успех размножения; взрослых самок, у которых не опереться птенец, разбегается около 28% по сравнению с 5% успешных заводчиков.[25] Натальное расселение (когда птица не возвращается на то место, где она родилась, чтобы размножаться) является обычным явлением для древесной ласточки и происходит чаще, чем размножение.[28] Гнездится как отдельными группами, так и отдельными парами. При гнездовании отдельными группами гнезда обычно располагаются на расстоянии не менее 10-15 метров (33-49 футов) друг от друга.[14] а те, что расположены ближе друг к другу, обычно находятся дальше друг от друга с точки зрения даты кладки.[29] В естественных пустотах древовидная ласточка гнездится на расстоянии около 27 метров (89 футов) от своего соседа. Гнездо в таких ситуациях находится в среднем на высоте 3,4 метра (11 футов) над уровнем земли, хотя около 45% из них находятся на высоте менее 2 метров (6,6 футов) над землей. Более высокие гнезда, вероятно, предпочтительны, потому что они уменьшают хищничество, в то время как более низкие гнезда могут быть выбраны, чтобы избежать конкуренции. Ширина входа часто составляет от 4 до 5 сантиметров (от 1,6 до 2,0 дюймов), тогда как высота входа более разнообразна: исследование 1989 года показало, что проемы варьируются от 3,5 до 26 см (от 1,4 до 10,2 дюйма). Объем полости обычно составляет менее 1000 кубических сантиметров (61 кубический дюйм).[30] Найдя подходящее место для гнездования, самец садится возле него и часто звонит. Недостаток мест может вызвать драки между птицами, иногда приводящие к гибели птиц. Эта ласточка обычно защищает территорию вокруг гнезда радиусом около 4,6 метра (15 футов).[7] а также дополнительные гнезда внутри этого территория,[14] заблокировав вход в гнездо и преследуя злоумышленников.[7] Сама чаша для гнезда сделана из травы, мха, хвои и водных растений, собранных в основном самками.[14] и покрыт перьями, собранными в основном самцами в драках.[31] Перья могут служить изоляцией гнезда, сокращая время инкубации.[32] и вероятно предотвращение переохлаждение у цыплят. Помимо более быстрого роста цыплят,[31] в гнездах с перьями яйца охлаждаются медленнее, чем в гнездах без перьев.[33] Однако исследование, опубликованное в 2018 году, не обнаружило значительной корреляции между количеством перьев в гнездах, которые были искусственно утеплены, по сравнению с количеством перьев, которые не были. Кроме того, было обнаружено, что гнездится в Сен-Дени, Саскачеван использовало значительно меньше перьев, чем в Долина Аннаполис, несмотря на то, что первый находится дальше на север. Однако температуры в Новой Шотландии (где находится долина Аннаполис) обычно ниже, чем в Саскачеване, что, возможно, объясняет неожиданный результат.[34]

Во время ухаживания самец ласточки нападает на неизвестную самку. Это может быть вызвано взмахом крыльев самки, что может быть приглашением в суд. Затем самец может принять вертикальную позу, с приподнятым и слегка раскрытым хвостом, а крылья взмахом и слегка опущенными. Это побуждает самку попытаться приземлиться на спину самца, но он летит, чтобы предотвратить это; это повторяется. Ухаживая за самкой, самец летит к выбранному им месту гнездования, которое самка осматривает. Во время совокупления самец парит над самкой, а затем садится на нее, издавая тикающие крики. Затем он делает клоакальный контакт с самкой, держа перья на шее своим клювом и стоя на слегка вытянутых крыльях. Копуляция происходит многократно.[7]
Яйца откладывают с начала мая до середины июня (правда, это происходит раньше из-за изменение климата[35]) и птенцы оперяются с середины июня по июль.[14] Широта положительно коррелирует с датой закладки,[14] возраст самок и длина крыльев (более длинные крылья позволяют более эффективно добывать корм[36]) имеют отрицательную корреляцию.[37] Древесная ласточка скорее всего заводчик дохода, поскольку размножение зависит от обилия пищи и температуры во время сезона яйцекладки.[36] Этот вид вообще социально моногамный, но до 8% размножающихся самцов полигинный.[14] На полигинию влияет территория: самцы, у которых есть территории с гнездовыми ящиками на расстоянии не менее 5 метров (16 футов) друг от друга, с большей вероятностью будут полигинными.[38] Предполагается, что эта полигиния зависит от условий в сезон яйцекладки: лучшие условия, такие как изобилие пищи, позволяют самкам в полигинии, которые не получают помощи в поисках пищи, откладывать больше яиц.[39]

Древесная ласточка имеет высокие показатели отцовства от дополнительной пары, от 38% до 69% птенцов являются продуктом отцовства от дополнительной пары, и от 50% до 87% выводков, содержащих хотя бы одного птенца, являющегося результатом отцовства дополнительной пары. совокупление.[14] Одним из факторов, который может способствовать этому, является то, что самки контролируют совокупление, что делает защиту отцовства неэффективной.[40] Это может быть смягчено более частыми совокуплениями непосредственно перед кладкой яиц, согласно исследованию 2009 года, которое обнаружило, что попытки спаривания внутри пары достигают пика за три-один день до того, как было отложено первое яйцо, и что более успешные попытки в этот период увеличивают долю внутрипарных копуляций. -пара молодых самцов. Этот последний вывод противоречит результатам исследований 1993 и 1994 годов.[41] Отцовство вне пары не влияет на уровень родительской заботы, которую самец вносит в ласточку дерева.[42] Значительное количество отцов экстра-пары могут быть плавающими (присутствующими на нерестилищах, которые предположительно не размножаются). Исследование 2001 года показало, что из 35 дополнительных пар птенцов 25 были рождены местными жителями, три - жителями близлежащих участков и семь - самцами плавучих особей. Таким образом, у древесной ласточки плавание помогает самцам в хорошем состоянии произвести больше птенцов, а самцам в плохом состоянии добиться успеха, вкладывая средства в родительскую заботу.[43] Существует также значительная популяция плавающих самок; исследование 1985 г. показало, что от 23% до 27% самок были плавающими, из которых от 47% до 79% были малолетними.[44]
Почему самки вступают в экстра-парные совокупления и как они выбирают себе экстра-пар, остается спорным. Одна теория, называемая гипотезой генетической совместимости, утверждает, что повышение приспособленности потомства является результатом увеличения гетерозиготность, и, таким образом, эти самки древесных ласточек предпочли бы спариваться с самцами, которые генетически менее схожи с ними. Самки также могут выбирать сперму после совокупления, чтобы обеспечить себе совместимого партнера. В поддержку этой теории исследование 2007 года показало, что потомство вне пары было более гетерозиготным, чем потомство внутри пары.[45] Однако в статье 2005 года обнаружена небольшая отрицательная корреляция между генетическим сходством пары и долей детенышей дополнительных пар в их гнезде.[46] Теория хороших генов гласит, что самки выбирают самцов из дополнительных пар на основе качества их генов. Это объясняет, почему у некоторых древесных ласточек нет лишних парных детенышей, а у других они есть.[40] Однако большинство исследований не обнаружили фенотипический различия между самцами вне пары и внутри пары (хотя исследование 2007 года обнаружило, что более старые самцы с более ярким оперением имели больше шансов спариваться вне пары).[47] Вдобавок, согласно тезису 2017 года, у потомства вне пары шансы присоединиться к своей родной популяции не выше, чем у потомства внутри пары. Другая теория предполагает, что отцовство в дополнительных парах зависит от контекста, при этом молодые люди из дополнительных пар в определенных ситуациях превосходят молодых внутри пары и хуже в других условиях. В диссертации 2017 года, например, было обнаружено, что детеныши дополнительных пар были крупнее, тяжелее и длиннее крыльев, чем детеныши внутри пары, когда оба подвергались нападению хищников, в то время как детеныши внутри пары были тяжелее, чем детеныши других пар, когда они были показаны животные, не являющиеся хищниками. Этот тезис также показал, что детеныши внутри пары превзошли детенышей дополнительных пар с точки зрения жизненной пригодности, когда они выросли в менее изменчивой среде, что позволяет предположить, что потомство в дополнительных парах имеет меньшую пластичность в развитии, чем потомство внутри пары.[48] Исследование 2018 года слабо поддержало эту контекстно-зависимую гипотезу, обнаружив, что потомство в дополнительных парах с большей вероятностью оперется, чем потомство внутри пары в экспериментально увеличенных выводках; однако ни длина теломер (коррелят выживаемости и репродуктивного успеха), ни размер через 12 дней после вылупления у этих молодых особей существенно не различались, и не было обнаружено значительных различий между этими двумя типами в неувеличенных выводках.[49]
Исследования, пытающиеся доказать адаптивность экстра-парного отцовства для самок, подвергались критике за отсутствие положительного эффекта, который мог бы иметь повышение приспособленности потомства по сравнению с потенциальной ценой снижения приспособленности для самок.[50] например, усиление хищничества из-за поиска партнера.[51] Таким образом, были постулированы теории, основанные на неадаптивности внепарного отцовства для женщин. Эти теории основаны на генетическое ограничение, где аллель, приводящий к неадаптивному поведению, сохраняется, потому что он также способствует благоприятному фенотипу. Теория интерсексуалов антагонистическая плейотропия говорит, что сильный отбор по отцовству дополнительной пары у самцов (как видно на этой птице[50]) отменяет слабый отбор против экстра-парного отцовства у самок. Между тем гипотеза об интрасексуальной антагонистической плейотропии утверждает, что отцовство вне пары присутствует, потому что гены, регулирующие его, оказывают плейотропное влияние на аспекты женской приспособленности,[52] как скорость совокупления внутри пары.[50]

Древесная ласточка кладет кладку от двух до восьми, хотя обычно от четырех до семи,[14] чисто-белые и полупрозрачные при кладке яйца размером около 19 на 14 миллиметров (0,75 на 0,55 дюйма).[53] Эти яйца насиживает самка,[14] обычно после снесения предпоследнего яйца,[54] от 11 до 20 дней,[53] хотя большинство выводится через 14-15 дней. Около 88% гнезд дают по крайней мере одного птенца, но это может быть уменьшено плохой погодой и появлением более молодой самки.[14] Яйца обычно вылупляются в том порядке, в котором они были отложены. Они также вылупляются немного асинхронно, в среднем 28 часов между появлением первого и последнего птенцов. Это может привести к иерархии веса, при которой ранее вылупившиеся птенцы весят больше (особенно в начале периода птенцов), чем вылупившиеся позже, что позволяет самке определять приоритеты, каким птенцам давать корм во время нехватки корма. Это, вероятно, имеет наибольший эффект в начале периода птенцов, так как через 12 дней после вылупления, как правило, нет значительных различий в весе.[54] Детоубийство цыплят и яиц иногда происходит, когда самец заменяется другим самцом. Детоубийство обычно не происходит, когда кладка не завершена, так как самцы-заменители имеют шанс оплодотворить по крайней мере одно яйцо. Когда самец прибывает во время инкубации, он иногда совершает детоубийство, но в других случаях принимает яйца, так как есть вероятность, что некоторые яйца были получены от замещающего самца. Если заменяющий самец появляется после вылупления птенцов, обычно совершается детоубийство, хотя самка иногда предотвращает это.[55]
В гнездах, которые производят более благополучные самки, соотношение полов часто смещено в сторону самцов высокого качества. Исследование 2000 г. выдвинуло гипотезу о том, что репродуктивный успех самцов более разный, и, следовательно, качественный самец производит больше потомства, чем самка аналогичного качества.[56]
На рост и выживание птенцов ласточек влияет среда, в которой они живут. Как у более молодых, так и у более взрослых птенцов (от двух до четырех дней и от девяти до одиннадцати дней соответственно) на рост положительно влияет более высокая максимальная температура, особенно у первых. Более поздний срок вылупления отрицательно сказывается на росте, особенно молодых птенцов. Птенцы постарше растут несколько быстрее, когда насекомых много. У молодых птенцов рост увеличивается с возрастом, у старых - уменьшается по мере взросления.[57] Молодые древесные ласточки способны терморегулировать не менее 75% так же эффективно, как и взрослые особи, в среднем в 9,5 дней вне гнезда и в возрасте от четырех до восьми дней в гнезде (в зависимости от размера расплода).[58] Птенцы оперяются примерно через 18–22 дня с успехом оперения около 80%. Как и на успех вылупления, на это негативно влияют неблагоприятная погода и более молодая самка.[14] На цыплят могут охотиться змеи[59] и еноты.[60] Это хищничество может усугубиться просьбами о попрошайничестве.[61]
Кормление

Дерево глотает корм на высоте до 50 метров (160 футов) над землей поодиночке или группами. Его полет представляет собой смесь взмахов и скольжения. Во время сезона размножения это в основном в пределах 4 км (2,5 миль) от места гнездования. Однако, когда он добывает пищу для птенцов, он обычно уходит на расстояние до 200 метров (660 футов) от гнезда.[14] в основном оставаясь на виду, и добывает корм на высоте до 12 метров (39 футов).[62] Как быть пойманным в полете,[7] насекомых иногда добывают с земли, воды, растений и вертикальных поверхностей.[14]
Древесная ласточка питается в основном насекомыми, а также некоторыми моллюсками, пауками и фруктами. В Северной Америке мухи составляют около 40% рациона, дополненного жуками и муравьями. Иначе в рационе около 90% мух.[14] Взятые насекомые представляют собой смесь водных и наземных организмов; первый является важным источником омега-3 высоконенасыщенные жирные кислоты. Это потому, что, хотя ласточка дерева может преобразовать предшественник α-линоленовая кислота в высоконенасыщенные жирные кислоты, такие как докозагексаеновая кислота, он не может сделать это в необходимых количествах.[63] Корм из семян и ягод в основном из родаMyrica, который в основном снимается во все четыре сезона в Северном полушарии, кроме лета. Ракообразные также играют важную роль в зимнем рационе в исследовании Лонг-Айленд, Нью-Йорк.[14]

Оба пола кормят птенцов (хотя самец кормит птенцов меньше, чем самки), что дает от 10 до 20 кормлений в час.[14] Родители часто используют щебетание, чтобы побудить птенцов попрошайничать. Это чаще используется с молодыми цыплятами, поскольку они просят меньше, чем старшие цыплята, когда родитель приходит с едой, но не зовет.[19] Вероятность попрошайничества в отсутствие родителей также увеличивается с возрастом.[64] Порядок вылупления влияет на то, сколько цыпленка кормят; Птенцы, вылупившиеся последним (в случаях, когда вылупление происходит асинхронно), скорее всего, получают меньше птенцов, чем вылупившихся ранее.[65] Птенцов ближе ко входу в гнездо также больше шансов накормить, как и тех, кто клянчат первым.[66] и чаще. Общая скорость кормления выводка также увеличивается с увеличением попрошайничества.[67] Сама диета состоит в основном из насекомых, находящихся в отрядах. Двукрылые, Hemiptera, и Одоната составляя большую часть диеты.[14] Эти насекомые в основном имеют размер до 10 миллиметров (0,39 дюйма), но иногда и до 60 миллиметров (2,4 дюйма) в длину.[62] В гнездах у озер подкисленный от человека, добавки кальция, в первую очередь рыбьи кости, раки экзоскелеты, раковины моллюсков и скорлупы птичьих яиц труднее найти. Это заставляет взрослую древесную ласточку путешествовать дальше обычного - иногда до 650 метров (2130 футов) от гнезда - чтобы получить эти кальциевые добавки.[68]
Выживание
Средняя продолжительность жизни древесной ласточки составляет 2,7 года.[69] и максимум 12 лет. Около 79% людей не доживают до своего первого года, а у тех, у кого действительно есть риск смертности от 40% до 60%.[14] Большинство смертей, вероятно, является результатом холодной погоды, которая снижает доступность насекомых, что приводит к голоду.[7] Продолжительность жизни связана с длиной теломер: исследование 2005 года, в котором в качестве показателя выживаемости использовалось количество возвращающихся (к месту размножения в предыдущем году), показало, что прогнозируемая продолжительность жизни людей с самыми длинными теломерами в возрасте одного года составляет 3,5 года, по сравнению до 1,2 года для обладателей самых коротких теломер.[70] Неизвестно, вызывают ли короткие теломеры ухудшение физической формы или просто показатель этого. Тем не менее, тезис 2016 года показал, что показатели состояния положительно коррелировали с длиной теломер. Самцы также обычно имели более длинные теломеры, чем самки, как и птицы с меньшими крыльями. Люди с более короткими теломерами могут компенсировать потенциальную потерю приспособленности за счет увеличения репродуктивных усилий, тогда как люди с более длинными теломерами могут уменьшить свои вложения, о чем свидетельствует меньшая доля цыплят-самок с более длинными теломерами.[71] Длина теломер передается по наследству и особенно зависит от длины теломер.[72]
Хищничество
Древесная ласточка восприимчива к целому ряду хищников. Яйца, птенцы и взрослые особи в гнезде становятся жертвами черные крысиные змеи, Американские вороны, Американские пустельги, обыкновенная трава, северные мерцания, бурундуки, олень, домашние кошки, ласки,[73] Американские черные медведи,[74] и еноты,[75] В полете или сидя на дереве хищниками являются американские пустельги, черноклювые сороки,[73] совы,[76] большие рогатые совы, Merlins, сапсаны, и острозубые ястребы. В то время как уклончивый бегство - обычная реакция на хищников в свободно летающих ласточках, вокруг гнезда часто встречается моббинг.[73] и направлена не только на хищников, но и на конкурентов в местах гнездования, которых это может отпугнуть.[20] Такое поведение предполагает, что ласточка роется и ныряет к злоумышленнику (но не поражает его).[73] от 5 до 20 метров (от 16 до 66 футов) над землей, обычно издавая тихий тикающий звук ближе к концу и приближаясь к хищнику на расстоянии примерно 0,5–2 метра (1,6–6,6 футов).[20] Кажется, что он меняет интенсивность своих атак в зависимости от того, какой хищник приближается;[73] исследование 1992 года показало, что хорьки вызывают более сильную защиту, чем черные крысиные змеи,[20] и тезис 2019 года аналогичным образом обнаружил, что модели черной крысы, по крайней мере, ныряли, и восточный бурундук моделей больше всего.[77] Предполагается, что змея вызвала более слабую реакцию, потому что защитное поведение может быть менее эффективным.[20] и более опасно выступать против него.[77]
Паразиты

Древесная ласточка уязвима для различных паразитов, например, паразитов крови.Трипаносома. Также он восприимчив к блохам.Ceratophyllus idius и перьевые клещиPteronyssoides tyrrelli, Труссартия, и (вероятно)Гемиальгес. Также, вероятно, поражается вшами из родовБруэлия иMyrsidea. Существует корреляция между количеством блох на птице и количеством птенцов, за которыми она ухаживает. Предполагается, что эта взаимосвязь является причиной улучшения микроклимата для блох из-за более крупной кладки.[78] Птенцы тоже страдают паразитами, как и мухи родаПротокаллифора,[79] что приводит к потере крови птенцами.[80] Эти паразиты, однако, встречаются в большинстве гнезд и, похоже, не оказывают большого влияния на птенцов. Исследование, опубликованное в 1992 году, показало, что эффекты паразитизма мясных мух объясняют лишь около 5,5% вариации массы птенцов.[79]
Иммунология
В размножении самка древесная ласточка, гуморальная иммунокомпетентность (HIC) обратно коррелирует с датой кладки. Это означает, что в среднем птица, откладывающая яйца раньше, имеет более сильный антибиотический ответ на антиген чем птица, которая откладывает позже. У древесной ласточки, которая лишена возможности обрезать крылья, обычно более низкий HIC. Эти отношения можно интерпретировать как поддержку вывода о том, что самка, откладывающая раньше, приобретает более высокий HIC, но авторы исследования, обнаружившего корреляции, считали это маловероятным из-за более низких температур в начале сезона размножения. Вместо этого они думали, что HIC может быть критерием качества, и что более качественная самка способна откладывать яйца раньше. Авторы также предположили, что это показатель рабочей нагрузки, о чем свидетельствует более низкий HIC у птиц-инвалидов.[81]
Самки древесных ласточек более высокого качества (по дате кладки) способны поддерживать репродуктивную функцию, отвлекая ресурсы на борьбу с иммунной проблемой. Глотки более низкого качества менее способны на это; исследование 2005 года в Итаке, штат Нью-Йорк, показало, что поздно кладущие самки с искусственно увеличенным выводком, хотя и способны поддерживать качество потомства, имеют более низкий ответ на иммунный вызов, чем те, которые были более качественными или не имели увеличенного выводка.[82] Выбор женщиной приоритета качества потомства или иммунокомпетентности, вероятно, связан с вероятностью выживания; исследование 2005 года показало, что самки с увеличенным выводком на Аляске, где показатели выживаемости ниже, имели более слабые иммунные ответы, но сохраняли стабильную репродуктивную способность, тогда как у самок в Теннесси с более высокими показателями выживаемости был более сильный ответ, но более низкое качество потомства.[83]
У древесной ласточки с возрастом ухудшаются некоторые компоненты иммунной системы. Приобретенный Т-клетка -опосредованный иммунитет, например, у самок ласточек с возрастом снижается. Но, возраст самки не влияет ни на приобретенное, ни на врожденное. гуморальный иммунитет; отсутствие ухудшения первых контрастирует с исследованиями по ласточки и женщина ошейники мухоловки.[84] Из-за этого иммунного старения (снижение иммунной функции с возрастом) инфицированные заболеванием самки старшего возраста обычно реже посещают свое гнездо, в результате чего их птенцы растут медленнее. Они также могут похудеть из-за инфекции.[85]
Положение дел
Древесная ласточка считается наименее опасной. Международный союз охраны природы. Это происходит из-за большого ареала обитания птиц - около 834 000 квадратных километров (322 000 квадратных миль),[1] и его стабильная популяция, оцениваемая примерно в 20 000 000 особей.[14] Он защищен в США Закон 1918 года о договоре о перелетных птицах,[86] и в Канаде Закон о Конвенции о перелетных птицах.[87] В некоторых частях США ареал этой ласточки расширился на юг, вероятно, из-за изменений в землепользовании, реинтродукции бобров и установки гнездовых ящиков для синие птицы. На древесную ласточку негативно влияют расчистка лесов и сокращение болот, последнее сокращает место обитания, доступное для зимовки. Этой ласточке приходится конкурировать за места гнездования с обыкновенный скворец, домашний воробей (оба представлены Северная Америка ),[14] синие птицы[88] и домашний крапивник (который также разрушает гнезда, не занимая их).[89] Подкисление озер может заставить эту ласточку преодолевать относительно большие расстояния в поисках продуктов, богатых кальцием, и может привести к тому, что цыплята будут есть пластик.[68] Другие химические вещества, такие как пестициды и другие загрязнители, могут сильно концентрироваться в яйцах, и Печатные платы связаны с отказом от парного клатча.[14] Загрязнение месторождений нефтеносных песков может негативно сказаться на ласточках деревьев, увеличивая количество токсинов, что измеряется по активности этоксирезоруфина.о-деэтилаза (фермент детоксикации) у птенцов. Обычно это мало влияет на птенцов и оперение.[90] though extreme weather can reveal the effects: a 2006 study found that nestlings from wetlands most polluted by oil sands processing material were more than 10 times more likely to die than those from a control site during periods of synchronized cold temperatures and heavy rainfall, compared to the lack of difference in mortality between the groups when the weather was less extreme.[91] A 2019 paper, however, found that increased precipitation caused a similar decline in hatching and nestling success for nestlings both near and far from oil sands sites.[92] In another study, birds exposed to mercury fledged, on average, one less chick than those not, an effect amplified by warm weather.[93] In addition, cold weather events can rapidly reduce the availability of aerial insect prey,[94] and in some populations with advancing reproduction may result in reduced offspring survival.[95]
Рекомендации
Примечания
Цитаты
- ^ а б BirdLife International (2016). "Tachycineta bicolor". Красный список видов, находящихся под угрозой исчезновения МСОП. 2016. Получено 6 августа 2018.CS1 maint: ref = harv (связь)
- ^ Vieillot, Louis Pierre (1807). Histoire naturelle des oiseaux de l'Amérique Septentrionale: contenant un grand nombre d'espèces décrites ou figurées pour la première fois (На французском). 1. Paris: Crapelet. п. 61.
- ^ Кабанис, Жан (1850). Museum Heineanum : Verzeichniss der ornithologischen Sammlung des Oberamtmann Ferdinand Heine auf Gut St. Burchard vor Halberstatdt (на немецком). 1. Halberstadt: Independently commissioned by R. Frantz. п. 48.
- ^ Coues, Elliott (1878). Birds of the Colorado Valley: a repository of scientific and popular information concerning North American ornithology. Вашингтон, округ Колумбия: Государственная типография.
- ^ а б Coues, Elliott (1882). A check list of North American birds (2-е изд.). Бостон: Эстес и Лориат. п. 42.
- ^ Turner, Angela (2017). дель Ойо, Хосеп; Эллиотт, Эндрю; Саргатал, Хорди; Кристи, Дэвид А .; де Хуана, Эдуардо (ред.). "White-rumped Swallow (Tachycineta leucorrhoa)". Справочник живых птиц мира. Lynx Edicions. Получено 14 января 2017.
- ^ а б c d е ж грамм час я j k л м Turner, Angela (2010). A Handbook to the Swallows and Martins of the World. Лондон: A&C Black. pp. 97–100. ISBN 978-1-4081-3172-5.
- ^ Джоблинг, Джеймс А. (2010). Словарь научных названий птиц Helm. Лондон: Кристофер Хелм. pp. 72, 377. ISBN 978-1-4081-2501-4.
- ^ Cerasale, David J.; Дор, Рой; Винклер, Дэвид В .; Lovette, Irby J. (2012). "Phylogeny of the Тахицинета genus of New World swallows: Insights from complete mitochondrial genomes" (PDF). Energy balance in migratory birds: Insights from mitochondrial genomes and leptin receptor biology (Кандидат наук). Корнелл Университет. С. 91–122.
- ^ Whittingham, Linda A; Сликас, Бет; Winkler, David W; Sheldon, Frederick H (2002). "Phylogeny of the tree swallow genus, Тахицинета (Aves: Hirundinidae), by Bayesian analysis of mitochondrial DNA sequences". Молекулярная филогенетика и эволюция. 22 (3): 430–441. Дои:10.1006 / mpev.2001.1073. ISSN 1055-7903. PMID 11884168.
- ^ Moore, William S. (1995). "Inferring phylogenies from mtDNA variation: Mitochondrial-gene trees versus nuclear-gene trees". Эволюция. 49 (4): 718–726. Дои:10.2307/2410325. ISSN 0014-3820. JSTOR 2410325. PMID 28565131.
- ^ Дор, Рой; Carling, Matthew D.; Ловетт, Ирби Дж .; Шелдон, Фредерик Х .; Winkler, David W. (2012). "Species trees for the tree swallows (Genus Тахицинета): An alternative phylogenetic hypothesis to the mitochondrial gene tree". Молекулярная филогенетика и эволюция. 65 (1): 317–322. Дои:10.1016 / j.ympev.2012.06.020. ISSN 1055-7903. PMID 22750631.
- ^ "Tree Swallow Identification, All About Birds, Cornell Lab of Ornithology". www.allaboutbirds.org. Получено 28 сентября 2020.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у z Turner, A. (2017). дель Ойо, Хосеп; Эллиотт, Эндрю; Саргатал, Хорди; Кристи, Дэвид А .; де Хуана, Эдуардо (ред.). "Tree Swallow (Tachycineta bicolor)". Справочник живых птиц мира. Barcelona, Spain: Lynx Edicions. Получено 10 декабря 2017.
- ^ а б Hussell, David J. T. (1983). "Age and plumage color in female tree swallows". Журнал полевой орнитологии. 54 (3): 312–318.
- ^ Stutchbury, Bridget J.; Robertson, Raleigh J. (October 1987). "Signaling subordinate and female status: Two hypotheses for the adaptive significance of subadult plumage in female tree swallows" (PDF). Аук. 104 (4): 717–723. Дои:10.1093/auk/104.4.717. ISSN 0004-8038.
- ^ Coady, Chelsea D.; Dawson, Russell D. (2013). "Subadult plumage color of female tree swallows (Tachycineta bicolor) reduces conspecific aggression during the breeding season". Журнал орнитологии Уилсона. 125 (2): 348–357. Дои:10.1676/12-155.1. ISSN 1559-4491. S2CID 85030082.
- ^ а б c Sharman, M. Y.; Robertson, R. J.; Ratcliffe, L. M. (1994). "Vocalizations of the tree swallow (Tachycineta bicolor) during the pre-laying period: A structural and contextual analysis". Американский натуралист из Мидленда. 132 (2): 264–274. Дои:10.2307/2426582. ISSN 0003-0031. JSTOR 2426582.
- ^ а б Леонард, Марти Л .; Fernandez, Nicole; Brown, Glen (1997). "Parental calls and nestling behavior in tree swallows". Аук. 114 (4): 668–672. Дои:10.2307/4089286. ISSN 0004-8038. JSTOR 4089286.
- ^ а б c d е Winkler, David W. (1992). "Causes and consequences of variation in parental defense behavior by tree swallows". Кондор. 94 (2): 502–520. Дои:10.2307/1369222. ISSN 0010-5422. JSTOR 1369222. S2CID 45389696.
- ^ McIntyre, Emma; Horn, Andrew G.; Leonard, Marty L. (2014). "Do nestling tree swallows (Tachycineta bicolor) respond to parental alarm calls?". Аук. 131 (3): 314–320. Дои:10.1642/AUK-13-235.1. ISSN 0004-8038. S2CID 86754716.
- ^ McIntyre, Emma; Леонард, Марти Л .; Horn, Andrew G. (2014). "Ambient noise and parental communication of predation risk in tree swallows, Tachycineta bicolor". Поведение животных. 87: 85–89. Дои:10.1016/j.anbehav.2013.10.013. ISSN 0003-3472. S2CID 53173370.
- ^ Леонард, Марти Л .; Horn, Andrew G.; Oswald, Krista N.; McIntyre, Emma (2015). "Effect of ambient noise on parent–offspring interactions in tree swallows". Поведение животных. 109: 1–7. Дои:10.1016/j.anbehav.2015.07.036. ISSN 0003-3472. S2CID 53160897.
- ^ Knight, Samantha M.; Bradley, David W.; Clark, Robert G.; Gow, Elizabeth A.; Bélisle, Marc; Berzins, Lisha L.; Blake, Tricia; Мост, Эли С .; Burke, Lauren; Dawson, Russell D.; Данн, Питер О .; Garant, Dany; Holroyd, Geoffrey L.; Hussell, David J. T.; Lansdorp, Olga; Laughlin, Andrew J.; Леонард, Марти Л .; Pelletier, Fanie; Shutler, Dave; Siefferman, Lynn; Taylor, Caz M.; Trefry, Helen E.; Vleck, Carol M.; Vleck, David; Винклер, Дэвид В .; Whittingham, Linda A.; Norris, D. Ryan (2018). "Constructing and evaluating a continent-wide migratory songbird network across the annual cycle". Экологические монографии. 88 (3): 445–460. Дои:10.1002/ecm.1298. ISSN 0012-9615.
- ^ а б Винклер, Дэвид В .; Wrege, Peter H.; Allen, Paul E.; Kast, Tracey L.; Senesac, Pixie; Wasson, Matthew F.; Llambías, Paulo E.; Ferretti, Valentina; Sullivan, Patrick J. (2004). "Breeding dispersal and philopatry in the tree swallow". Кондор (Представлена рукопись). 106 (4): 768–776. Дои:10.1650/7634. ISSN 0010-5422. S2CID 42958338.
- ^ Winkler, David W. (2006). "Roosts and migrations of swallows". El Hornero. 21 (2): 85–97.
- ^ Jones, Jason (2003). "Tree swallows (Tachycineta bicolor): A new model organism?". Аук. 120 (3): 591. Дои:10.1642/0004-8038(2003)120[0591:TSTBAN]2.0.CO;2. ISSN 0004-8038.
- ^ Shutler, Dave; Clark, Robert G. (2003). "Causes and consequences of tree swallow (Tachycineta bicolor) dispersal in Saskatchewan". Аук. 120 (3): 619–631. Дои:10.1642/0004-8038(2003)120[0619:CACOTS]2.0.CO;2. ISSN 0004-8038.
- ^ Muldal, Alison; Гиббс, Х. Лайл; Robertson, Raleigh J. (1985). "Preferred nest spacing of an obligate cavity-nesting bird, the tree swallow". Кондор. 87 (3): 356–363. Дои:10.2307/1367216. ISSN 0010-5422. JSTOR 1367216.
- ^ Rendell, Wallace B.; Robertson, Raleigh J. (1989). "Nest-site characteristics, reproductive success and cavity availability for tree swallows breeding in natural cavities". Кондор. 91 (4): 875. Дои:10.2307/1368072. ISSN 0010-5422. JSTOR 1368072.
- ^ а б Winkler, David W. (1993). "Use and importance of feathers as nest lining in tree swallows (Tachycineta bicolor)". Аук. 110 (1): 29–36.
- ^ Lombardo, Michael P.; Bosman, Ruth M.; Faro, Christine A.; Houtteman, Stephen G.; Kluisza, Timothy S. (1995). "Effect of feathers as nest insulation on incubation behavior and reproductive performance of tree swallows (Tachycineta bicolor)". Аук. 112 (4): 973–981. Дои:10.2307/4089028. ISSN 0004-8038. JSTOR 4089028.
- ^ Windsor, Rebecca L.; Fegely, Jessica L.; Ardia, Daniel R. (2013). "The effects of nest size and insulation on thermal properties of tree swallow nests". Журнал биологии птиц. 44 (4): 305–310. Дои:10.1111/j.1600-048X.2013.05768.x. ISSN 0908-8857.
- ^ Holland, Erika R.; Shutler, Dave (2018). "Nest feathering responses by tree swallows (Tachycineta bicolor) to experimental warming". Журнал орнитологии. 159 (4): 991–998. Дои:10.1007/s10336-018-1568-6. ISSN 2193-7192. S2CID 46971093.
- ^ Dunn, P. O.; Winkler, D. W. (1999). "Climate change has affected the breeding date of tree swallows throughout North America". Труды Королевского общества B: биологические науки. 266 (1437): 2487–2490. Дои:10.1098/rspb.1999.0950. ISSN 0962-8452. ЧВК 1690485. PMID 10693819.
- ^ а б Nooker, Jacqueline K.; Данн, Питер О .; Whittingham, Linda A. (2005). "Effects of food abundance, weather, and female condition on reproduction in tree swallows (Tachycineta bicolor)". Аук. 122 (4): 1225. Дои:10.1642/0004-8038(2005)122[1225:EOFAWA]2.0.CO;2. ISSN 0004-8038.
- ^ Винклер, Дэвид В .; Allen, Paul E. (1996). "The seasonal decline in tree swallow clutch size: physiological constraint or strategic adjustment?". Экология. 77 (3): 922–932. Дои:10.2307/2265512. ISSN 0012-9658. JSTOR 2265512. S2CID 84919928.
- ^ Данн, Питер О .; Hannon, Susan J. (1991). "Intraspecific competition and the maintenance of monogamy in tree swallows". Поведенческая экология. 2 (3): 258–266. Дои:10.1093/beheco/2.3.258. ISSN 1045-2249.
- ^ Данн, Питер О .; Hannon, Susan J. (1992). "Effects of food abundance and male parental care on reproductive success and monogamy in tree swallows". Аук. 109 (3): 488–499. ISSN 0004-8038.
- ^ а б Lifjeld, Jan T.; Данн, Питер О .; Robertson, Raleigh J.; Boag, Peter T. (1993). "Extra-pair paternity in monogamous tree swallows". Поведение животных. 45 (2): 213–229. Дои:10.1006/anbe.1993.1028. ISSN 0003-3472. S2CID 53195131.
- ^ Crowe, Susan A.; Kleven, Oddmund; Delmore, Kira E.; Laskemoen, Terje; Nocera, Joseph J.; Lifjeld, Jan T.; Robertson, Raleigh J. (2009). "Paternity assurance through frequent copulations in a wild passerine with intense sperm competition". Поведение животных. 77 (1): 183–187. Дои:10.1016/j.anbehav.2008.09.024. ISSN 0003-3472. S2CID 13722818.
- ^ Kempenaers, Bart; Lanctot, Richard B.; Robertson, Raleigh J. (1998). "Certainty of paternity and paternal investment in eastern bluebirds and tree swallows". Поведение животных. 55 (4): 845–860. Дои:10.1006/anbe.1997.0667. ISSN 0003-3472. PMID 9632472. S2CID 24760456.
- ^ Kempenaers, Bart; Everding, Susie; Bishop, Cheryl; Boag, Peter; Robertson, Raleigh J. (2001). "Extra-pair paternity and the reproductive role of male floaters in the tree swallow (Tachycineta bicolor)". Поведенческая экология и социобиология. 49 (4): 251–259. Дои:10.1007/s002650000305. ISSN 0340-5443. S2CID 25483760.
- ^ Stutchbury, Bridget J.; Robertson, Raleigh J. (1985). "Floating populations of female tree swallows". Аук. 102 (3): 651–654. Дои:10.1093/auk/102.3.651. ISSN 0004-8038.
- ^ Stapleton, Mary K.; Kleven, Oddmund; Lifjeld, Jan T.; Robertson, Raleigh J. (2007). "Female tree swallows (Tachycineta bicolor) increase offspring heterozygosity through extrapair mating". Поведенческая экология и социобиология. 61 (11): 1725–1733. Дои:10.1007/s00265-007-0404-4. ISSN 0340-5443. S2CID 22513713.
- ^ Barber, Colleen A.; Edwards, Mandy J.; Robertson, Raleigh J. (2005). "A test of the genetic compatibility hypothesis with tree swallows, Tachycineta bicolor". Канадский журнал зоологии. 83 (7): 955–961. Дои:10.1139/z05-091. ISSN 0008-4301.
- ^ Bitton, Pierre-Paul; O'Brien, Erin L.; Dawson, Russell D. (2007). "Plumage brightness and age predict extrapair fertilization success of male tree swallows, Tachycineta bicolor". Поведение животных. 74 (6): 1777–1784. Дои:10.1016/j.anbehav.2007.03.018. ISSN 0003-3472. S2CID 53170228.
- ^ Hallinger, Kelly Kristen (2017). Context-dependent fitness consequences of extra-pair paternity in tree swallows (Tachycineta bicolor) (Кандидат наук). Корнелл Университет.
- ^ Belmaker, Amos; Hallinger, Kelly K.; Glynn, Rebecca A.; Haussmann, Mark F.; Winkler, David W. (2018). "Is there a context-dependent advantage of extra-pair mating in tree swallows?". Аук. 135 (4): 998–1008. Дои:10.1642/AUK-18-3.1. HDL:10150/631782. ISSN 0004-8038. S2CID 91253157.
- ^ а б c Арнквист, Горан; Kirkpatrick, Mark (2005). "The evolution of infidelity in socially monogamous passerines: The strength of direct and indirect selection on extrapair copulation behavior in females". Американский натуралист. 165 (S5): S26–S37. Дои:10.1086/429350. ISSN 0003-0147. PMID 15795859. S2CID 32810854.
- ^ Данн, Питер О .; Whittingham, Linda A. (2006). "Search costs influence the spatial distribution, but not the level, of extra-pair mating in tree swallows". Поведенческая экология и социобиология. 61 (3): 449–454. Дои:10.1007/s00265-006-0272-3. ISSN 0340-5443. S2CID 4975256.
- ^ Forstmeier, Wolfgang; Накагава, Шиничи; Griffith, Simon C.; Kempenaers, Bart (2014). "Female extra-pair mating: adaptation or genetic constraint?". Тенденции в экологии и эволюции. 29 (8): 456–464. Дои:10.1016/j.tree.2014.05.005. ISSN 0169-5347. PMID 24909948.
- ^ а б Хаубер, Марк Э. (1 августа 2014 г.). Книга яиц: Путеводитель по яйцам шести сотен видов птиц в натуральную величину. Чикаго: Издательство Чикагского университета. п. 448. ISBN 978-0-226-05781-1.
- ^ а б Clotfelter, Ethan D.; Whittingham, Linda A.; Dunn, Peter O. (2000). "Laying order, hatching asynchrony and nestling body mass in tree swallows Tachycineta bicolor". Журнал биологии птиц. 31 (3): 329–334. Дои:10.1034/j.1600-048X.2000.310308.x. ISSN 0908-8857.
- ^ Robertson, Raleigh J. (1990). "Tactics and Counter-Tactics of Sexually Selected Infanticide in Tree Swallows". Population Biology of Passerine Birds: An Integrated Approach. Серия НАТО ASI. pp. 381–390. Дои:10.1007/978-3-642-75110-3_32. ISBN 978-3-540-51759-7.
- ^ Whittingham, Linda A.; Dunn, Peter O. (2000). "Offspring sex ratios in tree swallows: females in better condition produce more sons". Молекулярная экология. 9 (8): 1123–1129. Дои:10.1046/j.1365-294x.2000.00980.x. ISSN 0962-1083. PMID 10964231. S2CID 3720108.
- ^ McCarty, John P.; Winkler, David W. (2008). "Relative importance of environmental variables in determining the growth of nestling tree swallows Tachycineta bicolor". Ибис. 141 (2): 286–296. Дои:10.1111/j.1474-919X.1999.tb07551.x. ISSN 0019-1019.
- ^ Dunn, Erica H. (1979). "Age of effective homeothermy in nestling tree swallows according to brood size". Бюллетень Уилсона. 91 (3): 455–457. ISSN 0043-5643.
- ^ De Steven, Diane (1980). "Clutch size, breeding success, and parental survival in the tree swallow (Iridoprocne bicolor)". Эволюция. 34 (2): 278–291. Дои:10.1111/j.1558-5646.1980.tb04816.x. HDL:2027.42/137562. ISSN 0014-3820. PMID 28563429. S2CID 19454000.
- ^ Robertson, Raleigh J.; Rendell, Wallace B. (1990). "A comparison of the breeding ecology of a secondary cavity nesting bird, the tree swallow (Tachycineta bicolor), in nest boxes and natural cavities". Канадский журнал зоологии. 68 (5): 1046–1052. Дои:10.1139/z90-152. ISSN 0008-4301.
- ^ Leech, Susan M.; Leonard, Marty L. (1997). "Begging and the risk of predation in nestling birds". Поведенческая экология. 8 (6): 644–646. Дои:10.1093/beheco/8.6.644. ISSN 1045-2249.
- ^ а б McCarty, John P.; Winkler, David W. (1999). "Foraging ecology and diet selectivity of tree swallows feeding nestlings". Кондор. 101 (2): 246–254. Дои:10.2307/1369987. ISSN 0010-5422. JSTOR 1369987.
- ^ Twining, Cornelia W.; Lawrence, Peter; Винклер, Дэвид В .; Flecker, Alexander S.; Brenna, J. Thomas (2018). "Conversion efficiency of α-linolenic acid to omega-3 highly unsaturated fatty acids in aerial insectivore chicks". Журнал экспериментальной биологии. 221 (3): jeb165373. Дои:10.1242/jeb.165373. ISSN 0022-0949. PMID 29217628.
- ^ Леонард, Марти Л .; Horn, Andrew G. (2001). "Begging in the absence of parents by nestling tree swallows". Поведенческая экология. 12 (4): 501–505. Дои:10.1093/beheco/12.4.501. ISSN 1465-7279.
- ^ Johnson, L. Scott; Wimmers, Larry E.; Campbell, Sara; Hamilton, Lucy (2003). "Growth rate, size, and sex ratio of last-laid, last-hatched offspring in the tree swallow Tachycineta bicolor". Журнал биологии птиц. 34 (1): 35–43. Дои:10.1034/j.1600-048X.2003.02950.x. ISSN 0908-8857.
- ^ Whittingham, Linda A; Dunn, Peter O; Clotfelter, Ethan D (2003). "Parental allocation of food to nestling tree swallows: the influence of nestling behaviour, sex and paternity". Поведение животных. 65 (6): 1203–1210. Дои:10.1006/anbe.2003.2178. ISSN 0003-3472. S2CID 4993059.
- ^ Leonard, M. L.; Horn, Andrew G. (2001). "Begging calls and parental feeding decisions in tree swallows (Tachycineta bicolor)". Поведенческая экология и социобиология. 49 (2–3): 170–175. Дои:10.1007/s002650000290. ISSN 0340-5443. S2CID 25507198.
- ^ а б St. Louis, Vincent L.; Breebaart, Loes (1991). "Calcium supplements in the diet of nestling tree swallows near acid sensitive lakes". Кондор. 93 (2): 286–294. Дои:10.2307/1368944. ISSN 0010-5422. JSTOR 1368944.
- ^ Vleck, Carol M.; Vleck, David; Palacios, Maria G. (2011). "Evolutionary ecology of senescence: a case study using tree swallows, Tachycineta bicolor". Журнал орнитологии. 152 (S1): 203–211. Дои:10.1007/s10336-010-0629-2. ISSN 0021-8375. S2CID 25321642.
- ^ Haussmann, Mark F.; Винклер, Дэвид В .; Vleck, Carol M. (22 June 2005). "Longer telomeres associated with higher survival in birds". Письма о биологии. 1 (2): 212–214. Дои:10.1098/rsbl.2005.0301. ISSN 1744-9561. ЧВК 1626238. PMID 17148169.
- ^ Belmaker, Amos (2016). "Patterns of co-variation of telomere length, condition, life history and fitness in a short-lived bird species". The Role of Telomere Length in Tree Swallow Behavior and Life History (Кандидат наук). Корнелл Университет.
- ^ Belmaker, Amos; Hallinger, Kelly K.; Glynn, Rebbeca A.; Винклер, Дэвид В .; Haussmann, Mark F. (2019). "The environmental and genetic determinants of chick telomere length in tree swallows (Tachycineta bicolor)". Экология и эволюция. 9 (14): 8175–8186. Дои:10.1002/ece3.5386. ISSN 2045-7758. ЧВК 6662556. PMID 31380080.
- ^ а б c d е Winkler, D. W.; Hallinger, K. K.; Ardia, D. R.; Robertson, R. J.; Stutchbury, B. J.; Chohen, R. R. (2011). Poole, A. F., ed. "Tree Swallow (Tachycineta bicolor)". Птицы Северной Америки. Итака, Нью-Йорк: Cornell Lab of Ornithology.
- ^ Zach, Reto; Mayoh, Keith R. (1984). "Gamma radiation effects on nestling tree swallows". Экология. 65 (5): 1641–1647. Дои:10.2307/1939142. ISSN 0012-9658. JSTOR 1939142.
- ^ Chapman, L. B. (1955). "Studies of a tree swallow colony". Кольцевание птиц. 6 (2): 45–70.
- ^ Errington, P. L. (1932). "Food habits of southern Wisconsin raptors. Part I. Owls". Кондор. 34 (4): 176–186.
- ^ а б Maass, Natalia May (2019). Perceived predation risk and the responses of adult and nestling tree swallows (Tachycineta bicolor) (РС ). Университет Восточного Кентукки.
- ^ Shutler, Dave; Mullie, Adele; Clark, Robert G (2004). "Tree swallow reproductive investment, stress, and parasites". Канадский журнал зоологии. 82 (3): 442–448. CiteSeerX 10.1.1.530.1736. Дои:10.1139/z04-016. ISSN 0008-4301.
- ^ а б Roby, Daniel D.; Brink, Karen L.; Wittmann, Karin (1992). "Effects of bird blowfly parasitism on eastern bluebird and tree swallow nestlings" (PDF). Бюллетень Уилсона. 104 (4): 630–643.
- ^ DeSimone, Joely G.; Clotfelter, Ethan D.; Black, Elizabeth C.; Knutie, Sarah A. (2018). "Avoidance, tolerance, and resistance to ectoparasites in nestling and adult tree swallows". Журнал биологии птиц. 49 (2): jav–01641. Дои:10.1111/jav.01641. ISSN 0908-8857.
- ^ Hasselquist, Dennis; Wasson, Matthew F.; Winkler, David W. (2001). "Humoral immunocompetence correlates with date of egg-laying and reflects work load in female tree swallows". Поведенческая экология. 12 (1): 93–97. Дои:10.1093/oxfordjournals.beheco.a000384. ISSN 1465-7279.
- ^ Ardia, Daniel R. (2005). "Individual quality mediates trade-offs between reproductive effort and immune function in tree swallows". Журнал экологии животных. 74 (3): 517–524. Дои:10.1111/j.1365-2656.2005.00950.x. ISSN 0021-8790.
- ^ Ardia, Daniel R. (2005). "Tree swallows trade off immune function and reproductive effort differently across their range". Экология. 86 (8): 2040–2046. Дои:10.1890/04-1619. ISSN 0012-9658. S2CID 33354318.
- ^ Palacios, Maria G; Cunnick, Joan E; Winkler, David W; Vleck, Carol M (2007). "Immunosenescence in some but not all immune components in a free-living vertebrate, the tree swallow". Труды Королевского общества B: биологические науки. 274 (1612): 951–957. Дои:10.1098/rspb.2006.0192. ISSN 0962-8452. ЧВК 2141670. PMID 17251097.
- ^ Palacios, Maria G.; Винклер, Дэвид В .; Klasing, Kirk C.; Hasselquist, Dennis; Vleck, Carol M. (2011). "Consequences of immune system aging in nature: a study of immunosenescence costs in free-living tree swallows". Экология (Представлена рукопись). 92 (4): 952–966. Дои:10.1890/10-0662.1. ISSN 0012-9658. PMID 21661557.
- ^ "Migratory Bird Treaty Act Protected Species (10.13 List)". Служба рыболовства и дикой природы США. 2013. Получено 4 июн 2018.
- ^ "Birds protected under the Migratory Birds Convention Act". Правительство Канады. 2017. Архивировано с оригинал 20 мая 2019 г.. Получено 21 июля 2018.
- ^ Wiebe, Karen L. (2016). "Interspecific competition for nests: Prior ownership trumps resource holding potential for mountain bluebird competing with tree swallow". Аук. 133 (3): 512–519. Дои:10.1642/AUK-16-25.1. ISSN 0004-8038.
- ^ Finch, Deborah M. (1990). "Effects of predation and competitor interference on nesting success of house wrens and tree swallows". Кондор. 92 (3): 674–687. Дои:10.2307/1368686. ISSN 0010-5422. JSTOR 1368686.
- ^ Smits, Judit E.; Wayland, Mark E.; Миллер, Майкл Дж .; Liber, Karsten; Trudeau, Suzanne (2000). "Reproductive, immune, and physiological end points in tree swallows on reclaimed oil sands mine sites". Экологическая токсикология и химия. 19 (12): 2951–2960. Дои:10.1002/etc.5620191216. ISSN 0730-7268.
- ^ Gentes, Marie-Line; Waldner, Cheryl; Papp, Zsuzsanna; Smits, Judit E.G. (2006). "Effects of oil sands tailings compounds and harsh weather on mortality rates, growth and detoxification efforts in nestling tree swallows (Tachycineta bicolor)". Загрязнение окружающей среды. 142 (1): 24–33. Дои:10.1016/j.envpol.2005.09.013. ISSN 0269-7491. PMID 16297515.
- ^ Godwin, Christine M.; Barclay, Robert M. R.; Smits, Judit E. G. (2019). "Tree swallow (Tachycineta bicolor) nest success and nestling growth near oil sands mining operations in northeastern Alberta, Canada". Канадский журнал зоологии. 97 (6): 547–557. Дои:10.1139/cjz-2018-0247. ISSN 0008-4301.
- ^ Hallinger, Kelly K.; Cristol, Daniel A. (2011). "The role of weather in mediating the effect of mercury exposure on reproductive success in tree swallows". Экотоксикология. 20 (6): 1368–1377. Дои:10.1007/s10646-011-0694-1. ISSN 0963-9292. PMID 21553259. S2CID 42492417.
- ^ Винклер, Дэвид В .; Luo, Miles K.; Rakhimberdiev, Eldar (September 2013). "Temperature effects on food supply and chick mortality in tree swallows (Tachycineta bicolor)". Oecologia. 173 (1): 129–138. Bibcode:2013Oecol.173..129W. Дои:10.1007/s00442-013-2605-z. ISSN 0029-8549. ЧВК 3751296. PMID 23468236.
- ^ Shipley, J. Ryan; Twining, Cornelia W.; Taff, Conor C.; Vitousek, Maren N.; Flack, Andrea; Winkler, David W. (28 September 2020). "Birds advancing lay dates with warming springs face greater risk of chick mortality". Труды Национальной академии наук. 117 (41): 25590–25594. Дои:10.1073/pnas.2009864117. ISSN 0027-8424. PMID 32989166. S2CID 222159036.
внешняя ссылка
| Авторитетный контроль |
|---|