Разобщающий белок - Uncoupling protein

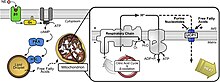
An разобщающий белок (UCP) представляет собой белок внутренней мембраны митохондрий, который является регулируемым протонным каналом или переносчиком. Таким образом, разобщающий белок способен рассеивать протон градиент создано НАДН -силовая перекачка протонов из митохондриального матрикса в митохондриальное межмембранное пространство. Энергия, теряемая при рассеянии протонного градиента через UCP, не используется для биохимической работы. Вместо этого выделяется тепло. Это то, что связывает UCP с термогенезом. Однако не все типы UCP связаны с термогенезом. Хотя UCP2 и UCP3 тесно связаны с UCP1, UCP2 и UCP3 не влияют на терморегуляторные способности позвоночных.[1] UCP расположены на той же мембране, что и АТФ-синтаза, который также является протонным каналом. Таким образом, два белка работают параллельно: один генерирует тепло, а другой генерирует АТФ из АДФ и неорганического фосфата, что является последним этапом в окислительного фосфорилирования.[2] Дыхание митохондрий связано с синтезом АТФ (фосфорилирование АДФ), но регулируется UCP.[3] UCP принадлежат к митохондриальный носитель (SLC25) семейство.
Расщепление белков играет роль в нормальной физиологии, например, при воздействии холода или спячка, потому что энергия используется для выработки тепла (см. термогенез ) вместо того, чтобы производить АТФ. Некоторые виды растений используют тепло, выделяемое при расщеплении белков, для специальных целей. Восточная скунсовая капуста, например, поддерживает температуру своих шипов на 20 ° C выше, чем температура окружающей среды, распространяя запах и привлекая насекомых, удобряющих цветы.[4] Однако другие вещества, такие как 2,4-динитрофенол и карбонилцианид м-хлорфенилгидразон, также выполняют ту же функцию разъединения. Салициловая кислота также является разобщающим агентом (главным образом в растениях) и снижает выработку АТФ и повышает температуру тела, если принимать его в чрезмерном избытке.[5] Расщепляющие белки увеличиваются гормон щитовидной железы, норэпинефрин, адреналин, и лептин.[6]
История
Ученые наблюдали термогенную активность в коричневая жировая ткань, что в конечном итоге привело к открытию UCP1, первоначально известного как «Uncoupling Protein».[3] Коричневая ткань показала повышенные уровни дыхания митохондрий и другого дыхания, не связанного с синтезом АТФ, что символизировало сильную термогенную активность.[3] UCP1 был обнаруженным белком, ответственным за активацию протонного пути, который не был связан с фосфорилированием АДФ (обычно осуществляется через АТФ-синтаза ).[3]
У млекопитающих
У млекопитающих известно пять гомологов UCP. Хотя каждый из них выполняет уникальные функции, определенные функции выполняются несколькими гомологами. Гомологи следующие:
- UCP1, также известный как термогенин или SLC25A7
- UCP2, также известный как SLC25A8
- UCP3, также известный как SLC25A9
- UCP4, также известный как SLC25A27
- UCP5, также известный как SLC25A14
Поддержание температуры тела
Первый обнаруженный разобщающий белок, UCP1, был обнаружен в коричневая жировая ткань из спящие и мелких грызунов, которые обеспечивают этим животным не дрожащее тепло.[3] Эти коричневые жировые ткани необходимы для поддержания температуры тела мелких грызунов, и исследования с (UCP1) -нокаутные мыши показывают, что эти ткани не функционируют правильно без функционирования разобщающих белков.[3] Фактически, эти исследования показали, что холодовая акклиматизация невозможна для этих мышей с нокаутом, что указывает на то, что UCP1 является важным фактором производства тепла в этих коричневых жировых тканях.[7][8]
Известно, что в других частях тела разобщающая активность белков влияет на температуру в микросредах.[9][10] Считается, что это влияет на активность других белков в этих областях, хотя все еще требуется работа, чтобы определить истинные последствия градиентов температуры, вызванных разобщением внутри клеток.[9]
Роль в концентрациях АТФ
Влияние UCP2 и UCP3 на концентрацию АТФ варьируется в зависимости от типа клеток.[9] Например, бета-клетки поджелудочной железы испытывать уменьшение АТФ концентрация с повышенной активностью UCP2.[9] Это связано с дегенерацией клеток, уменьшением инсулин секреция и диабет II типа.[9][11] И наоборот, UCP2 в гиппокамп клетки и UCP3 в мышечных клетках стимулируют выработку митохондрии.[9][12] Большее количество митохондрий увеличивает общую концентрацию ADP и АТФ, что фактически приводит к чистому увеличению концентрации АТФ, когда эти разобщающие белки становятся связанными (то есть механизм, позволяющий протекать протонам, ингибируется).[9][12]
Поддержание концентрации активных форм кислорода
Полный перечень функций UCP2 и UCP3 неизвестен.[13] Однако исследования показывают, что эти белки участвуют в петля отрицательной обратной связи ограничение концентрации активные формы кислорода (ROS).[14] Текущий научный консенсус утверждает, что UCP2 и UCP3 осуществляют перенос протонов только тогда, когда активация виды присутствуют.[15] Среди этих активаторов - жирные кислоты, АФК и некоторые побочные продукты АФК, которые также являются реактивными.[14][15] Следовательно, более высокие уровни ROS прямо или косвенно вызывают повышение активности UCP2 и UCP3.[14] Это, в свою очередь, увеличивает утечку протонов из митохондрий, снижая протонодвижущую силу через митохондриальные мембраны, активируя цепь переноса электронов.[13][14][15] Ограничение движущей силы протона посредством этого процесса приводит к отрицательной обратной связи, которая ограничивает производство АФК.[14] В частности, UCP2 снижает трансмембранный потенциал митохондрий, тем самым уменьшая продукцию ROS. Таким образом, раковые клетки могут увеличивать продукцию UCP2 в митохондриях.[16] Эта теория подтверждается независимыми исследованиями, которые показывают повышенную продукцию ROS у мышей с нокаутом UCP2 и UCP3.[15]
Этот процесс важен для здоровья человека, поскольку считается, что высокие концентрации АФК участвуют в развитии дегенеративных заболеваний.[15]
Функции нейронов
Обнаружив связанный мРНК, UCP2, UCP4 и UCP5, как было показано, располагаются в нейронах центральной нервной системы человека.[17] Эти белки играют ключевую роль в функции нейронов.[9] Хотя результаты многих исследований остаются противоречивыми, некоторые из них получили широкое признание.[9]
Например, UCP изменяют концентрацию свободного кальция в нейроне.[9] Митохондрии являются основным местом хранения кальция в нейронах, и емкость накопления увеличивается с потенциалом через митохондриальные мембраны.[9][18] Следовательно, когда разобщающие белки снижают потенциал через эти мембраны, ионы кальция высвобождаются в окружающую среду нейрона.[9] Из-за высокой концентрации митохондрий вблизи терминалы аксонов, это означает, что UCP играют роль в регулировании концентрации кальция в этой области.[9] Учитывая, что ионы кальция играют большую роль в нейротрансмиссии, ученые предсказывают, что эти UCP напрямую влияют на нейротрансмиссию.[9]
Как обсуждалось выше, нейроны в гиппокампе испытывают повышенные концентрации АТФ в присутствии этих разобщающих белков.[9][12] Это заставляет ученых предположить, что UCP улучшают синаптическую пластичность и передачу.[9]
Рекомендации
- ^ Gaudry MJ, Jastroch M (март 2019). «Молекулярная эволюция разобщающих белков и последствия для функции мозга». Письма о неврологии. 696: 140–145. Дои:10.1016 / j.neulet.2018.12.027. PMID 30582970.
- ^ Nedergaard J, Ricquier D, Kozak LP (октябрь 2005 г.). «Разобщение белков: современное состояние и терапевтические перспективы». EMBO отчеты. 6 (10): 917–21. Дои:10.1038 / sj.embor.7400532. ЧВК 1369193. PMID 16179945.
- ^ а б c d е ж Rousset S, Alves-Guerra MC, Mozo J, Miroux B, Cassard-Doulcier AM, Bouillaud F, Ricquier D (февраль 2004 г.). «Биология митохондриальных разобщающих белков». Сахарный диабет. 53 (приложение 1): S130-5. Дои:10.2337 / диабет.53.2007.S130. PMID 14749278.
- ^ Гарретт Р.Х., Гришем К.М. (2013). Биохимия (Пятое издание, международное издание). Китай: Мэри Финч. п. 668. ISBN 978-1-133-10879-5.
- ^ «Калифорнийская система борьбы с отравлениями: салицилаты». Архивировано из оригинал на 2014-08-02.
- ^ Гонг Д.В., Хе И, Карас М., Рейтман М. (сентябрь 1997 г.). «Разобщающий белок-3 является медиатором термогенеза, регулируемого тироидным гормоном, бета3-адренергическими агонистами и лептином».. Журнал биологической химии. 272 (39): 24129–32. Дои:10.1074 / jbc.272.39.24129. PMID 9305858.
- ^ Хаген Т., Видаль-Пуч А. (февраль 2002 г.). «Митохондриальные разобщающие белки в физиологии и болезни человека». Минерва Медика. 93 (1): 41–57. PMID 11850613.
- ^ Feldmann HM, Golozoubova V, Cannon B, Nedergaard J (февраль 2009 г.). «Абляция UCP1 вызывает ожирение и устраняет термогенез, вызванный диетой, у мышей, освобожденных от теплового стресса за счет термонейтральности». Клеточный метаболизм. 9 (2): 203–9. Дои:10.1016 / j.cmet.2008.12.014. PMID 19187776.
- ^ а б c d е ж грамм час я j k л м п о п Эндрюс З. Б., Диано С., Хорват Т. Л. (ноябрь 2005 г.). «Митохондриальные разобщающие белки в ЦНС: в поддержку функции и выживания». Обзоры природы. Неврология. 6 (11): 829–40. Дои:10.1038 / №1767. PMID 16224498.
- ^ Хорват Т.Л., надзиратель С.Х., Хаджос М., Ломбарди А., Гоглиа Ф., Диано С. (декабрь 1999 г.). «Разобщающий белок 2 мозга: несвязанные митохондрии нейронов предсказывают тепловые синапсы в гомеостатических центрах». Журнал неврологии. 19 (23): 10417–27. PMID 10575039.
- ^ Zhang CY, Baffy G, Perret P, Krauss S, Peroni O, Grujic D, et al. (Июнь 2001 г.). «Несвязанный белок-2 отрицательно регулирует секрецию инсулина и является основным звеном между ожирением, дисфункцией бета-клеток и диабетом 2 типа». Клетка. 105 (6): 745–55. Дои:10.1016 / s0092-8674 (01) 00378-6. PMID 11440717.
- ^ а б c Диано С., Мэтьюз Р. Т., Патрило П., Ян Л., Бил М. Ф., Барнстейбл С. Дж., Хорват Т. Л. (ноябрь 2003 г.). «Разобщающий белок 2 предотвращает гибель нейронов, в том числе происходящую во время припадков: механизм предварительного кондиционирования». Эндокринология. 144 (11): 5014–21. Дои:10.1210 / en.2003-0667. PMID 12960023.
- ^ а б Ястроч М., Дивакаруни А.С., Мукерджи С., Треберг-младший, бренд-MD (14.06.2010). «Митохондриальные протоны и утечки электронов». Очерки биохимии. 47: 53–67. Дои:10.1042 / bse0470053. ЧВК 3122475. PMID 20533900.
- ^ а б c d е Mailloux RJ, Harper ME (сентябрь 2011 г.). «Разобщение белков и контроль митохондриальной продукции активных форм кислорода». Свободная радикальная биология и медицина. 51 (6): 1106–15. Дои:10.1016 / j.freeradbiomed.2011.06.022. PMID 21762777.
- ^ а б c d е Бренд MD, Esteves TC (август 2005 г.). «Физиологические функции митохондриальных разобщающих белков UCP2 и UCP3». Клеточный метаболизм. 2 (2): 85–93. Дои:10.1016 / j.cmet.2005.06.002. PMID 16098826.
- ^ Сридхар А., Чжао Ю. (май 2017 г.). «Разъединение белка 2 и метаболические заболевания». Митохондрия. 34: 135–140. Дои:10.1016 / j.mito.2017.03.005. ЧВК 5477468. PMID 28351676.
- ^ Ричард Д., Клавель С., Хуанг К., Санчис Д., Рикье Д. (ноябрь 2001 г.). «Разобщающий белок 2 в головном мозге: распределение и функции». Сделки Биохимического Общества. 29 (Пт 6): 812–7. Дои:10.1042 / bst0290812. PMID 11709080.
- ^ Николс Д.Г., Ward MW (апрель 2000 г.). «Митохондриальный мембранный потенциал и нейрональная эксайтотоксичность глутамата: смертность и милливольты». Тенденции в неврологии. 23 (4): 166–74. Дои:10.1016 / s0166-2236 (99) 01534-9. PMID 10717676.