Сексуальная мимикрия - Sexual mimicry
Сексуальная мимикрия возникает, когда один пол имитирует противоположный пол в своем поведении, внешности или химических сигналах. Это чаще встречается у видов беспозвоночных, хотя сексуальная мимикрия также наблюдается у позвоночных, таких как пятнистые гиены. Сексуальная мимикрия обычно используется как стратегия спаривания, чтобы получить доступ к партнеру, как защитный механизм, чтобы избежать большего. доминирующий люди или стратегия выживания. Это также может быть физическая характеристика, определяющая место человека в обществе. Сексуальная мимикрия используется по-разному у разных видов и является частью их стратегии выживания и воспроизводства. Примеры сексуального мимикрия в животных включают пятнистая гиена, некоторые виды рыб, воробьиные птицы и некоторые виды насекомых среди других. Это случаи внутривидовой сексуальной мимикрии, но межвидовая сексуальная мимикрия также может встречаться у некоторых видов растений, особенно у орхидей. У растений, использующих половую мимикрию, цветы имитируют брачные сигналы насекомых-опылителей. Эти насекомые привлекаются и опыляют цветы через псевдокопуляции или другое сексуальное поведение, совершаемое с цветком.
Социальные системы
Сексуальная мимикрия может влиять на социальную систему вида. Самый распространенный пример - пятнистые гиены, Crocuta crocuta. Самки гиены похожи на самцов гиен по своей сексуальной анатомии: у самок клитор в форме пениса, напоминающий пенис, и ложные мошоночные мешки. Эти характеристики, а также высокие андроген уровни в их крови делают самок агрессивными, что приводит к их преобладанию над самцами; женщина с самым низким рангом доминирует над мужчиной с самым высоким рангом. Внутри женского населения в каждом клане есть разные ранги: доминирующие женщины, которые размножаются в более раннем возрасте и получают больший доступ к пище, и недоминантные женщины. Их господство иерархично и передается от матери к дочери. Напротив, пятнистые гиены-самцы получают свой социальный статус с продолжительностью своего пребывания в клане;[1] он не предполагает агрессивных соревнований. Самцы покидают свой клан в возрасте от двух до шести лет. [2] и вступают в другой клан, где с возрастом получают статус. Мужчины также способствуют установлению дружеских отношений с женщинами, чтобы стабилизировать свое положение в социальной иерархии.[3]

Поскольку среди пятнистых гиен доминирующим полом являются самки, они пользуются наибольшим уважением. Подчиненные гиены начинают «приветствовать» доминирующих гиен в знак уважения и вынуждены делать это, если отказываются. Это приветствие, используемое гиенами, отражает асимметрию их ранжирования; приветствуемое животное (подчиненное лицо) вытягивает задние лапы, а человек, делающий приветствие (доминирующая гиена), облизывает или обнюхивает эрегированный клитор в форме пениса.[3] Поднимая задние лапы, приветствуемая гиена (подчиненная гиена) показывает свою самую уязвимую часть тела другому человеку, что отражает неполноценность. Кроме того, когда его задние лапы подняты, другой особь может определить запах. Подчиненные гиены обнаруживают свой запах чаще, чем высокопоставленные гиены. Однако это приветствие не часто встречается между мужчинами и взрослыми женщинами; когда это действительно происходит, это ограничивается мужчинами среднего или более высокого ранга, приветствующими доминирующих женщин.[3]
Системы спаривания
У пятнистых гиен единственный способ спаривания самцов с самками - это полное сотрудничество самки из-за ее полового клитора. Повышение статуса самцов дало им больше доступа к доминирующим самкам в клане. Доминирующие самки гиены не спариваются с несколькими самцами, возможно, из-за затрат на чистку гениталий, которые гиены делают после совокупления.[1] Поскольку они получат доступ к наиболее доминирующим и наиболее приспособленным самцам, им не нужно совокупляться с несколькими самцами, чтобы произвести потомство более приспособленных. Недоминантные самки чаще совокупляются с самцами более низкого ранга.[3] Самкам гиен приходится дорого рожать через длинный пенистый клитор. Длина пуповины составляет 12–18 см, а расстояние от матки до конца клитора составляет 40 см. Пуповина часто рвется еще до выхода детеныша, что приводит к смерти многих детенышей от аноксии. Это путешествие вредно не только для детенышей, но и для матери. Ткань клитора иногда разрывается при первых родах, что может быть фатальным для матери.[4]
Пятнистые гиены женского пола - разборчивый пол, потому что они вкладывают средства в родительскую заботу, а также являются доминирующим полом в клане. Однако самцы, вероятно, по-прежнему будут отдавать предпочтение определенной самке, как это наблюдается у других животных; самки с высоким рейтингом начинают размножаться в более молодом возрасте, и их потомство с большей вероятностью доживет до взрослой жизни, чем потомство самок с низким рейтингом. Самцы более тесно связаны с фертильными самками, и это состояние, скорее всего, можно заметить по обонятельным сигналам.[5] В то время как мужчины среднего / высокого ранга ассоциируются с женщинами высокого ранга, мужчины низкого ранга в равной степени ассоциируются с женщинами высокого и низкого ранга. Связь с самками с низким рейтингом может происходить из-за того, что самцы с низким рейтингом не могут распознать репродуктивный успех самок с высоким рейтингом или используют другой тип репродуктивной стратегии. Самцы, как правило, проводят много времени с самкой до зачатия, чтобы другие самцы не вступали с ней в тесный контакт.[5]
Сексуальная мимикрия также используется некоторыми видами в качестве стратегии защиты партнера. Охрана партнера - это процесс, в котором член одного вида не позволяет другому представителю того же вида спариваться со своим партнером. Охрана товарища видна в Cotesia rubecula, паразитическая оса из семейства Braconidae чья система спаривания полигинный. Самцов привлекают самки через феромоны, и они побуждают самок спариваться посредством вибраций, на которые самка реагирует, занимая определенное положение. Когда мужчина, совокуплявшийся с женщиной, видит, что другой мужчина пытается ухаживать за ней, он часто принимает женскую восприимчивую позицию. Пост-копулятивная женская мимика самца дает преимущество, так как действует как механизм защиты партнера. Если второй самец появляется достаточно скоро после совокупления самки с первым самцом, второй самец может вызвать второе совокупление, которое будет конкурировать с первым. Однако, если первый самец, совокуплявшийся с ней, подражает самке, это отвлекает второго самца на достаточно долгое время, и самка становится невосприимчивой.[6]
Подлый совокупление


Скрытное совокупление - это стратегия, используемая многими водными организмами, изображающими сексуальную мимикрию. Несколько исследований показали, что маленькие самцы рыб будут выглядеть и вести себя как самки своего вида, чтобы получить доступ к территории самок и совокупиться с ними.[7][8] В семье рыб Blenniidae, женщина Салария паво покажут определенный цветовой узор и движение, когда они хотят приблизиться к мужчине и совокупиться с ним. Самец охраняет территорию, и когда самка откладывает яйца, родительский самец защищает эту территорию, пока не вылупятся яйца. Второй тип самцов, самцы-тапки, паразитирует и напоминает самок блениид своим маленьким размером, цветом и характером движений. Это позволяет им вторгаться в гнездо, охраняемое родительскими самцами. Самцы-тапки подходят к гнездам с той же окраской и движениями, что и самки. В большинстве случаев самцы-тапки наблюдаются, когда самка уже находится внутри гнезда, хотя иногда рыба-тапка входит в гнездо вместе с самкой. Этот вид рыб выделяет сперму до того, как самка выпускает икру в воду.[9] позволяя рыбе-тапке оплодотворить яйцо, даже если самки нет в гнезде.[7]

В семье Сепиина Сепия апама, также известный как каракатица, есть несколько крупных самцов, способных охранять гнездо самки, в то время как другие самцы маленькие и напоминают самок, чтобы тайком пробираться во время совокуплений. У гигантских каракатиц самец ухаживает за самкой и переносит ее сперму в мешочек под клювом самки. Во время этого процесса самка демонстрирует узор из черных пятен на белом фоне. После того, как яйца отложены, самец охраняет гнездо от возможных женихов и противников. «Вторую женщину» иногда можно увидеть во время взаимодействия мужчины и женщины в непосредственной близости от пары. У этой похожей на самку каракатицы такие же черные пятна, как у настоящей самки. Если самец уходит, чтобы сразиться с другими самцами, эта особь приближается к самке и совокупляется с ней, как правило, успешно. Однако в отсутствие соперников эти «имитирующие самку» самцы проявляют фенотип зрелого самца.[8]
Сексуальная мимикрия против агрессии
Похожее явление с самцами-тапками наблюдается у расколотый плавник с темными краями, Girardinichthys multiradiatus. Молодые особи похожи на беременных самок этого вида тем, что имеют темное пятно возле отверстия. Однако в этом случае имитирующие самцы обладают способностью походить на самок или становиться морфологически зрелыми самцами на протяжении большей части своей взрослой жизни. Это темное пятно позволяет самцам, похожим на самок, избегать агрессии со стороны более доминирующих самцов, а также снижает вероятность того, что самка сбегает из-за упорных ухаживаний самцов. Взрослые самцы не нападают на подчиненную рыбу, а подчиненная рыба решает, когда начинать драку, что дает ей преимущество, поскольку зрелый самец этого не ожидает. Темное пятно также обеспечивает доступ подчиненных самцов к самкам, что является преимуществом, поскольку яйца самок могут быть оплодотворены только в течение пятидневного периода оплодотворения.[10]


Сексуальная мимика во избежание агрессии наблюдается и у птиц. У некоторых видов птиц у самцов на втором году жизни окраска оперения напоминает самку (самцы SY). Эти самцы SY являются половозрелыми и способны к размножению, но их морфология сильно отличается от более старых самцов после второго года жизни (ASY). Различные исследования изучали эту задержку созревания оперения (DPM) и обнаружили, что DPM у самцов SY снижает агрессию со стороны самцов ASY.[11][12][13] Самка мимикрии у птиц впервые была обнаружена у мухоловки-пеструшки, Ficedula hypoleuca. Когда поблизости находится тусклый самец, зрелые самцы снижают свою агрессивность и ведут себя так, как будто злоумышленник - самка. Тусклое оперение наблюдается в основном у молодых самцов, вероятно, из-за того, что они родились позже, прошлой весной. Сходство с самками приносит пользу этим молодым самцам при попытке занять территорию, на которой уже присутствует много самцов, потому что молодые самцы могут получить информацию и доступ к территории, которая в противном случае была бы для них недоступна.[12]
Когда дело доходит до защиты территории или привлечения партнера, то не выглядеть как мужчина - это большая цена. Самки проявляют агрессию по отношению к самцам тусклого окраса, из-за чего им сложнее спариваться.[12] Однако DPM имеет некоторые преимущества: как упоминалось выше, он снижает агрессию со стороны пожилых мужчин.[12] Кроме того, эти похожие на самок птицы могут получить доступ к территориям, партнерам и еде, которые в противном случае могут быть им недоступны. Другое преимущество состоит в том, что DPM обеспечивает птицам SY более продолжительную продолжительность жизни; поскольку им не приходится соревноваться с другими самцами, их смертность ниже. Это преимущество, однако, приносит пользу только тем видам, которые имеют более длительную потенциальную продолжительность жизни, и, следовательно, DPM не принесет пользу короткоживущим видам.[11] Это известно как гипотеза порога размножения и гласит, что самцы SY должны откладывать размножение только в том случае, если существует большая разница в смертности между самцами SY, которые пытаются размножаться, и теми, кто этого не делает.[11]

Большинство исследований рассматривали DPM как тип сексуальной мимикрии,[11][12] что делается путем обмана: самцы птиц ASY не должны отличать самок или самцов SY. Однако Muheter et al. (1997) обнаружили, что территориальные самцы воспринимают тусклых самцов как самцов, но проявляют меньше агрессии, потому что их тусклое оперение способствует низкой конкурентоспособности. Они назвали это честная сигнализация а не сексуальная мимикрия.[13]
Другой пример сексуальной мимикрии встречается в «Плоской ящерице» Бродли, Platysaurus broadleyi, где некоторые самцы имитируют самок. Самцы плоских ящериц обычно территориальный и агрессивен по отношению к другим мужчинам. Поэтому некоторым самцам выгодно подражать самкам, чтобы избегать агрессивных столкновений и свободно перемещаться по территории самцов в поисках партнеров. В этой популяции есть два типа мужчин; самцы, имитирующие женщин, и самцы, похожие на мужчин. Самцы могут визуально обмануть самцов, заставив их поверить в то, что они женщины, из-за их женской морфологии. Однако самцы не могут обмануть самцов через запах, поскольку самцы могут заметить разницу. Таким образом, наиболее успешными самцами являются те, кто избегает тесного контакта с другими самцами, тем самым снижая вероятность обнаружения с помощью химических сигналов.[14]
Молекулярный контроль над сексуальной мимикрией
Половая имитация самок гиен и самцов является частью их анатомии и, как полагают, эволюционировала благодаря высокому уровню андрогенов. Хотя предки-самки были меньше самцов, отбор должен был воздействовать на уровни андрогенов и размер женского тела, чтобы увеличить и то, и другое, что привело к дальнейшему отбору и к более крупным самкам, чем самцам.[15] Высокие уровни андрогенов отсутствуют в женских яичниках, как когда-то считалось;[16] стромальная ткань яичников содержит более низкий уровень тестостерона, чем мужские семенники. Однако уровни андрогенов у женщин в крови такие же высокие, как и у мужчин, что имеет эффект морфологически похожих на мужчин женщин.[17]
Ерши также могут демонстрировать сексуальную мимикрию благодаря сочетанию генетики и гормонов. В популяции ерши, Philomachus pugnax, есть три типа мужских морфов: независимые самцы и самцы-спутники, оба из которых являются репродуктивными конкурентами, и фейдерные ерши, которые по своему оперению напоминают самок. Первые две морфы контролируются доминантным аллелем в одном аутосомном локусе, в то время как третья морфа, вероятно, возникла из комбинации третьего аллеля и отсутствия тестостерон.[18] Когда тестостерон вводят в рифы (женские ерши), ухаживание самцов и окраска перьев самцов выражаются в рифах. В этом случае тестостерон проявляет ограниченные по полу характеристики, воздействуя на единственный аутосомный ген.[19] Точно так же, хотя это еще не было проверено, вполне вероятно, что недостаток тестостерона является причиной сходства фейдерных ерш с самками.

Другой пример наблюдается у половозрелых самок плодовых мух, Drosophila melanogaster, которые очень привлекательны, но их уровень привлекательности снижается наполовину или более после трех минут спаривания.[20] Во время ухаживания самцы выделяют в самку соединение 7-трикосин, которое снижает привлекательность самок. Однако исследователь обнаружил, что самки также выделяют это соединение через шесть часов после спаривания. Это соединение снижает уровень привлекательности самки как во время ухаживания самца, так и во время спаривания. Таким образом, самка имитирует самца, и с помощью этого соединения она снижает уровень своей привлекательности.[21]
Генетический контроль над сексуальной мимикрией
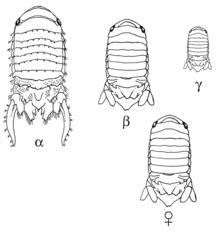
Сексуальная мимикрия некоторых организмов генетически определяется конкретными аллелями. В отличие от сексуальной мимикрии, которая возникает из-за молекулярных соединений или гормонов и иногда может быть вызвана этими молекулами, эта сексуальная мимикрия возникает из генетического материала организма. Помимо половой анатомии самок гиен, которая является частью их генетики, у некоторых других организмов в популяции есть только несколько самцов / самок, которые выглядят как противоположный пол, и это определяется определенными аллелями.
В популяции морских изопод Paracerceis sculpta, существует три различных морфологии самцов: альфа-самец является самой крупной морфой, он созревает последним и именно он получает привилегированный доступ к самкам. Бета-самец среднего размера, и он имитирует самку, чтобы получить доступ к самкам. Наконец, гамма-самец является самым маленьким морфом и вторгается в гаремы, где самки идут спариваться с альфа-самцами для возможности спаривания. Эта морфология связана с одним аутосомным геном и тремя разными аллелями. Бета - это самый доминирующий аллель, за ним следует гамма, за которой следует альфа. Отбор по этим аллелям действует в соответствии с равновесием Харди-Вайнберга, и успех спаривания одинаков для всех трех морфов.[22]
Альфа-самцы, которые гомозиготный для альфа-аллеля спариваться со многими самками в гареме. Самки предпочитают объединяться с другими самками в гареме, что дает альфа-самцу больший выбор партнеров для спаривания. Шустер (1992) изучил поведение и взаимосвязь каждой морфы по отношению к гарему и обнаружил, что бета- и гамма-самцы могут определять местонахождение гаремов, у которых есть сексуально восприимчивые самки. Они также смогли различить гарем с сексуально восприимчивой самкой, то есть той, которая способна спариваться, и несексуально восприимчивой самкой, то есть той, которая уже поместила эмбрион в свою сумку и больше не может спариваться. Хотя до сих пор неясно, как бета-самцы делают это или как работают их стратегии спаривания, альфа-самцы не преследуют их из-за того, что они подражают самкам: бета-самцы могут привлекать других самок в гарем, поскольку самки любят идти туда, где другие самки. есть, и это дает альфа-самцам больше партнеров.[23]
Другой отряд организмов, чья половая мимикрия определяется их ДНК, - это Одоната, плотоядные насекомые, известные как стрекозы и стрекозы. У этих видов самка иногда подражает самцу. Внутри вида группы самок различаются по цвету: одна группа имитирует цвет самцов, и они известны как андрохромы. У других групп будет собственная женская окраска, и они известны как гинохромы. В Ischnura elegans, андрохромы составляют 6-30% женского населения, и их цвет обычно синий, как и у мужчин; в некоторых популяциях андрохромы крупнее гинохромов. Этот полиморфизм контролируется аутосомным аллелем, и в некоторых исследованиях была изучена причина сохранения полиморфизма.[24]


Наиболее вероятной теорией сохранения полиморфизма Odonata является теория зависимости от плотности. [25] в котором говорится, что при высокой плотности мужчин андрохромы не беспокоят мужчин и их существованию не угрожают домогательства со стороны мужчин. Эта гипотеза также предполагает, что самцы не могут отличить андрохромов от других самцов. Этому преимуществу, однако, противодействует тот факт, что они не получат много возможностей для спаривания (если таковые имеются), а их воспроизводство ограничено. Эта теория является наиболее вероятным объяснением сохранения полиморфизма, поскольку исследования показали, что андрохромы имеют преимущество в популяциях с высокой мужской плотностью.[26]
Самоконтроль сексуальной мимикрии
Хотя, как было замечено ранее, большинство организмов, изображающих сексуальную мимикрию, рождаются с этой морфологией / поведением, это не всегда так. Гигантская каракатица, Сепия апама, упомянутый выше в разделе «скрытые совокупления», рождается со способностью выбирать, изменять ли свою морфологию, чтобы выглядеть как женщина или зрелый мужчина. Когда поблизости не видно конкурентов, каракатица будет выглядеть как взрослый самец и спариться с самкой. Однако, когда зрелый самец и самка совокупляются, гигантская каракатица будет напоминать самку и оставаться на близком расстоянии от пары, надеясь на шанс спариться с самкой, если зрелый самец уйдет, чтобы сразиться с другими самцами.[8] Другой пример организма, который может оставаться маленьким и выглядеть как самка или становиться морфологически зрелым самцом, - это тёмный двуногий двуногий плавник, Girardinichthys multiradiatus. Цель их женской мимикрии была замечена ранее в разделе «Сексуальная мимикрия против агрессии», где самцы женского вида избегают агрессии со стороны доминирующих самцов и избегают бегства самок из их компании из-за упорных ухаживаний за самцами.[10]
Межвидовая обманчивая мимикрия

Межвидовая половая мимикрия также может встречаться у некоторых видов растений. Самый распространенный пример этого - опыление с обманом полов, которое встречается у некоторых орхидей.[27] Орхидея имитирует самок своего опылителя, обычно перепончатокрылые такие как осы и пчелы, привлекая к цветку самцов. Цветки орхидей имитируют половые феромоны и до некоторой степени внешний вид самок насекомых своего вида-опылителя. Примат обоняния над визуальными сигналами был продемонстрирован во многих случаях, например, в европейском роду орхидей. Офрис а также многие австралийские сексуально обманчивые орхидеи. В некоторых других случаях, например, южноафриканская ромашка Gorteria diffusa, визуальные сигналы имеют первостепенное значение.[28] Визуальные сигналы также повышают привлекательность цветков некоторых видов Ophrys для опылителей.[29][30] Какой-то мужчина чешуйчатые осы Такие как Campsoscolia ciliata больше привлекают Офрис запахов цветов, чем запахов самок ос, хотя оба они привлекают самцов одними и теми же соединениями. Скорее всего, это результат более сильного запаха цветов орхидеи; самки ос, как правило, издают меньше запаха, чтобы не привлекать хищников.[31] Независимо от того, используют ли орхидеи внешний вид, аромат или и то, и другое, они имитируют самку-опылителя для собственной выгоды.
Смотрите также
Рекомендации
- ^ а б Восточный ML (1 сентября 2001 г.). «Пятнистые гиены-самцы (Crocuta crocuta) в очереди за статусом в социальных группах, где преобладают самки». Поведенческая экология. 12 (5): 558–568. Дои:10.1093 / beheco / 12.5.558.
- ^ Смейл Л., Нуньес С., Холекамп К.Е. (1997). «Половой диморфизм у млекопитающих: закономерности, причины и последствия». В Slater PJ, Rosenblatt JS, Snowdon CT, Milinski M (ред.). Достижения в учебном поведении. 26. Академическая пресса. С. 181–250. ISBN 9780080582870.
- ^ а б c d Восток М.Л., Берк Т., Вильгельм К., Крейг С., Хофер Х (2003). «Сексуальные конфликты у пятнистых гиен: тактика спаривания самцов и самок и их репродуктивный результат в зависимости от возраста, социального статуса и владения». Труды Королевского общества B. 270 (1521): 1247–54. Дои:10.1098 / rspb.2003.2363. ЧВК 1691369. PMID 12816637.
- ^ Фрэнк Л.Г. (февраль 1997 г.). «Эволюция генитальной маскулинизации: почему у самок гиен такой большой« пенис »?». Тенденции в экологии и эволюции. 12 (2): 58–62. Дои:10.1016 / S0169-5347 (96) 10063-X. PMID 21237973.
- ^ а б Шикман М., Энг А.Л., Хорн Р.К., Функ С.М., Скрибнер К.Т., Холекамп К.Е. (2001). «Модели ассоциации между самцами и самками пятнистых гиен (Crocuta crocuta) отражают выбор самца». Поведенческая экология и социобиология. 50 (3): 231–238. Дои:10.1007 / s002650100356. JSTOR 4601958.
- ^ Филд С.А., Келлер М.А. (декабрь 1993 г.). «Альтернативная тактика спаривания и мимикрия самок как пост-копулятивное поведение по охране спаривания у паразитической осы Cotesia rubecula». Поведение животных. 46 (6): 1183–1189. Дои:10.1006 / anbe.1993.1308.
- ^ а б Гонсалвеш Э.Дж., Алмада В.К., Оливейра РФ, Сантос А.Дж. (11 мая 2009 г.). "Женская мимикрия как тактика спаривания у самцов Blenniid Fish Salaria Pavo". Журнал Морской биологической ассоциации Соединенного Королевства. 76 (2): 529. Дои:10.1017 / S0025315400030721.
- ^ а б c Norman MD, Finn J, Tregenza T. (7 июля 1999 г.). «Имитация самки как альтернативная репродуктивная стратегия гигантских каракатиц». Труды Королевского общества B. 266 (1426): 1347–1349. Дои:10.1098 / rspb.1999.0786. ЧВК 1690068.
- ^ Пацнер Р.А. (1984). "Воспроизведение Blennius pavo (Teleostei Bleniidae). II. Структура поверхности зрелого яйца ». Zoologischer Anzeiger. 213: 44–50.
- ^ а б Масиас-Гарсия К., Валеро А. (19 мая 2010 г.). «Контекстно-зависимая сексуальная мимикрия у живородящих рыб». Этология, экология и эволюция. 13 (4): 331–339. Дои:10.1080/08927014.2001.9522764.
- ^ а б c d Стадд М.В., Робертсон Р.Дж. (1985). «Продолжительность жизни, конкуренция и задержка созревания оперения у самцов воробьиных: гипотеза порога размножения». Американский натуралист. 126 (1): 101–115. Дои:10.1086/284399. JSTOR 2461565.
- ^ а б c d е Slagsvold T, Saetre G (июнь 1991 г.). "Эволюция окраски оперения у самцов мухоловок-пеструшек (Ficedula Hyopleuca): Доказательства женской мимикрии ». Эволюция. 45 (4): 910–917. Дои:10.1111 / j.1558-5646.1991.tb04359.x. PMID 28564056.
- ^ а б Мюхтер В.Р., Грин Э., Рэтклифф Л. (27 октября 1997 г.). «Задержка созревания оперения у лазуритовых овсянок: тесты на мимикрию самок и гипотезы о статусных сигналах». Поведенческая экология и социобиология. 41 (4): 281–290. Дои:10.1007 / s002650050389.
- ^ Уайтинг MJ, Уэбб JK, Keogh JS (25 февраля 2009 г.). «Подражатели плоских ящериц используют сексуальный обман в визуальных, но не химических сигналах». Труды Королевского общества B. 276 (1662): 1585–1591. Дои:10.1098 / rspb.2008.1822. ЧВК 2660994. PMID 19324828.
- ^ Гамильтон В.Дж., 3-е место, Тилсон Р.Л., Фрэнк Л.Г. (26 апреля 2010 г.). «Половой мономорфизм у пятнистых гиен, Crocuta crocuta». Этология. 71 (1): 63–73. Дои:10.1111 / j.1439-0310.1986.tb00570.x.
- ^ Мэтьюз LH (5 июля 1939). «Размножение гиены пятнистой, Crocuta crocuta (Erxleben)». Философские труды Королевского общества B. 230 (565): 1–78. Дои:10.1098 / рстб.1939.0004.
- ^ Рейси PA, Скиннер JD (20 августа 2009 г.). «Эндокринные аспекты половой мимикрии у пятнистых гиен Crocuta crocuta». Журнал зоологии. 187 (3): 315–326. Дои:10.1111 / j.1469-7998.1979.tb03372.x.
- ^ Ланк ДБ, Фаррелл Л.Л., Берк Т., Пирсма Т., Макрей С.Б. (6 ноября 2013 г.). «Доминантный аллель контролирует развитие женского пола, имитирующего мужские и миниатюрные женские ерши». Письма о биологии. 9 (6): 20130653. Дои:10.1098 / rsbl.2013.0653. ЧВК 3871350. PMID 24196515.
- ^ Lank DB, Coupe M, Wynne-Edwards KE (22 ноября 1999 г.). «Вызванные тестостероном мужские черты у самок ершей (Philomachus pugnax): аутосомное наследование и гендерная дифференциация». Труды Королевского общества B. 266 (1435): 2323–2330. Дои:10.1098 / rspb.1999.0926. ЧВК 1690456.
- ^ Томпкинс Л., Холл Дж. К. (январь 1981 г.). «Различное воздействие на ухаживание летучих соединений от спарившихся и девственных самок дрозофилы». Журнал физиологии насекомых. 27 (1): 17–21. Дои:10.1016/0022-1910(81)90026-3.
- ^ Скотт Д. (ноябрь 1986 г.). «Сексуальная мимикрия регулирует привлекательность спарившихся самок Drosophila melanogaster» (PDF). Труды Национальной академии наук Соединенных Штатов Америки. 83 (21): 8429–33. Дои:10.1073 / pnas.83.21.8429. ЧВК 386942. PMID 3095835.
- ^ Шустер С.М., Уэйд М.Дж. (18 апреля 1991 г.). «Равный успех спаривания среди мужских репродуктивных стратегий у морских изопод». Природа. 350 (6319): 608–610. Дои:10.1038 / 350608a0.
- ^ Шустер С.М. (1992). «Репродуктивное поведение морфов α-, β- и γ-самцов у Paracerceis sculpta, морских ракообразных изопод» (PDF). Поведение. 121 (3/4): 231–258. Дои:10.1163 / 156853992X00381. JSTOR 4535029.
- ^ Кордеро А., Карбоне СС, Утзери С. (январь 1998 г.). «Возможности спаривания и затраты на спаривание снижаются у самок андрохромных стрекоз Ischnura elegans (Odonata)». Поведение животных. 55 (1): 185–197. Дои:10.1006 / anbe.1997.0603. PMID 9480685.
- ^ Hinnekint BO. 1987 г. Динамика численности населения Ишнуры e. Elegans (Vnader Linden) (Insecta: Odonata) с особым упором на морфологические изменения окраски, женский полиморфизм, многолетние циклы и их влияние на поведение. Hydobiologia. 146: 3-31.
- ^ Кордеро А., Андрес Дж. А. (1996). «Цветовой полиморфизм у стрекоз: самки, имитирующие самцов?». Журнал Британского общества стрекоз . 12 (2): 50–60.
- ^ Schiestl FP (1 июня 2005 г.). «Об успехе мошенничества: опыление обманом орхидей». Naturwissenschaften. 92 (6): 255–264. Дои:10.1007 / s00114-005-0636-у. HDL:20.500.11850/32223. PMID 15931514.
- ^ Ellis AG, Johnson SD (ноябрь 2010 г.). «Цветочная мимикрия увеличивает экспорт пыльцы: эволюция опыления путем сексуального обмана за пределами орхидных». Американский натуралист. 176 (5): E143 – E151. Дои:10.1086/656487. PMID 20843263.
- ^ Gaskett AC, Herberstein ME (2 октября 2009 г.). «Цветовая мимикрия и сексуальный обман со стороны языковых орхидей (Cryptostylis)». Naturwissenschaften. 97 (1): 97–102. Дои:10.1007 / s00114-009-0611-0. PMID 19798479.
- ^ Певец RB, Flach A, Koehler S, Marsaioli AJ, Amaral Mdo C (июнь 2004 г.). «Сексуальная мимикрия у Mormolyca ringens (Lindl.) Schltr. (Orchidaceae: Maxillariinae)». Анналы ботаники. 93 (6): 755–62. Дои:10.1093 / aob / mch091. ЧВК 4242296. PMID 15051623.
- ^ Ayasse M, Schiestl FP, Paulus HF, Ibarra F, Francke W. (7 марта 2003 г.). «Привлечение опылителей к орхидеям, обманывающим сексуальность, с помощью нетрадиционных химикатов». Труды Королевского общества B. 270 (1514): 517–522. Дои:10.1098 / rspb.2002.2271. ЧВК 1691269. PMID 12641907.
