Стимулятор генов интерферона - Stimulator of interferon genes





Стимулятор генов интерферона (STING), также известный как трансмембранный белок 173 (TMEM173) и MPYS/MITA/ЭРИС это белок что у человека кодируется STING1 ген.[5]
STING играет важную роль в врожденный иммунитет. STING вызывает интерферон I типа продукция, когда клетки инфицированы внутриклеточными патогенами, такими как вирусы, микобактерии и внутриклеточные паразиты.[6] Интерферон I типа, опосредованный STING, защищает инфицированные и близлежащие клетки от местной инфекции, связываясь с той же клеткой, которая его секретирует (автокринный сигнализация) и близлежащие клетки (паракринный сигнализация.)
СТИНГ работает как прямой цитозольный датчик ДНК (CDS) и адаптерный белок в Интерферон I типа передача сигналов через различные молекулярные механизмы. Было показано, что он активирует нижестоящие факторы транскрипции. STAT6 и IRF3 через TBK1, которые отвечают за противовирусный ответ и врожденный иммунный ответ против внутриклеточный патоген.[7]
Структура

Аминокислоты 1–379 человеческого STING включают 4 трансмембранные области (TM) и С-концевой домен. В С-концевой домен (CTD: аминокислоты 138–379) содержит домен димеризации (DD) и карбокси-концевой хвост (СТТ: аминокислоты 340–379).[7]
СТИНГ образует в клетке симметричный димер. Димер STING напоминает бабочку с глубокой щелью между двумя протомерами. В гидрофобный остатки от каждой формы протомера STING гидрофобные взаимодействия между собой на интерфейсе.[7][8]
Выражение
STING выражается в кроветворный клетки в периферические лимфоидные ткани, включая Т-лимфоциты, NK-клетки, миелоидные клетки и моноциты. Также было показано, что STING сильно экспрессируется в легких, яичник, сердце, гладкая мышца, сетчатка, Костный мозг и влагалище.[9][10]
Локализация
Субклеточная локализация STING была выяснена как эндоплазматический ретикулум белок. Кроме того, вполне вероятно, что СТИНГ ассоциируется в непосредственной близости с митохондриально-ассоциированная мембрана ER (MAM) -поверхность между митохондрией и ER.[11] Во время внутриклеточной инфекции STING может перемещаться из эндоплазматический ретикулум к перинуклеарные пузырьки потенциально участвует в экзоциста опосредованный транспорт.[11] Также было показано, что STING колокализируется с белками аутофагии, ассоциированный с микротрубочками белок 1 легкая цепь 3 (LC3) и белок 9A, связанный с аутофагией, после стимуляции двухцепочечной ДНК, что позволяет предположить его присутствие в аутофагосома.[12]
Функция
STING выступает посредником интерферон I типа продукция в ответ на внутриклеточную ДНК и различные внутриклеточные патогены, включая вирусы, внутриклеточные бактерии и внутриклеточные паразиты.[13] При заражении СИНГ от инфицированных клеток может определять присутствие нуклеиновые кислоты от внутриклеточных патогенов, а затем вызвать интерферон β и более 10 форм интерферон α производство. Интерферон I типа продуцируемые инфицированными клетками, могут находить и связываться с Рецептор интерферона-альфа / бета ближайших клеток, чтобы защитить клетки от местной инфекции.
Противовирусный иммунитет
STING вызывает мощный интерфероновый иммунитет I типа против вирусной инфекции. После вирусное проникновение, популярный нуклеиновые кислоты будет присутствовать в цитозоле инфицированных клеток. Несколько датчиков ДНК, таких как DAI, РНК-полимераза III, IFI16, DDX41 и cGAS, может обнаруживать посторонние нуклеиновые кислоты. После распознавания вирусной ДНК сенсоры ДНК инициируют нижестоящие сигнальные пути, активируя STING-опосредованный ответ интерферона.[14]
Аденовирус, Вирус простого герпеса, HSV-1 и HSV-2, а также вирус с отрицательной цепью РНК -вирус везикулярного стоматита (VSV) было показано, что они могут активировать зависящие от СИНГА врожденный иммунный ответ.[13]
Дефицит STING у мышей приводил к летальной восприимчивости к инфекции HSV-1 из-за отсутствия успешного ответа интерфероном типа I.[15]
Точечная мутация серина S358 подавляет активацию STING-IFN у летучих мышей и, как предполагается, дает летучим мышам их способность в качестве резервуарных хозяев.[16]
Против внутриклеточных бактерий
Внутриклеточные бактерии, Listeria monocytogenes, как было показано, стимулируют иммунный ответ хозяина через STING.[17] STING может играть важную роль в производстве МКП-1 и CCL7 хемокины. Моноциты с дефицитом STING по своей природе не могут мигрировать в печень во время Listeria monocytogenes инфекционное заболевание. Таким образом, STING защищает хост от Listeria monocytogenes инфекция путем регулирования моноцит миграция. Активация STING, вероятно, будет опосредована: циклический ди-АМФ секретируются внутриклеточными бактериями.[17][18]
Другой
STING может быть важной молекулой защитного иммунитета против инфекционных организмов. Например, животные, которые не могут экспрессировать СТИНГ, более восприимчивы к инфекции от VSV, HSV-1 и Listeria monocytogenes, предполагая его потенциальную связь с инфекционными заболеваниями человека.[19]
Роль в иммунитете хозяина
Несмотря на то что IFN типа I абсолютно критичен для устойчивости к вирусам, появляется все больше литературы о негативной роли интерферон I типа в иммунитете хозяина, опосредованном STING. Богатый AT мотив ДНК стволовой петли в Плазмодий falciparum и Плазмодий бергей геном и внеклеточная ДНК из Микобактерии туберкулеза было показано, чтобы активировать интерферон I типа через СТИНГ.[20][21] Перфорация мембраны фагосомы опосредована ESX1 система секреции позволяет внеклеточной микобактериальной ДНК получать доступ к сенсорам цитозольной ДНК хозяина, тем самым вызывая выработку интерферон I типа в макрофагах. Высоко интерферон I типа подпись ведет к М. туберкулез патогенез и длительное инфицирование.[21] STING-TBK1-IRF опосредованный интерферон I типа ответ является центральным в патогенезе экспериментальной церебральной малярии у лабораторных животных, инфицированных Плазмодий бергей. Лабораторные мыши с дефицитом интерферон I типа ответ устойчивы к экспериментальной церебральной малярии.[20]
Механизмы передачи сигналов STING
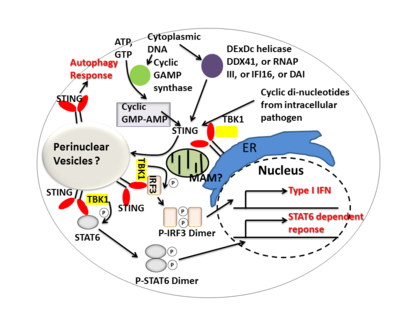
STING выступает посредником интерферон I типа иммунный ответ, действуя как прямой датчик ДНК и сигнальный адаптерный белок. При активации СТИНГ стимулирует TBK1 активность по фосфорилированию IRF3 или же STAT6. Фосфорилированные IRF3 и STAT6 димеризуются, а затем проникают в ядро, чтобы стимулировать экспрессию генов, участвующих в иммунном ответе хозяина, таких как IFNB, CCL2, CCL20, так далее.[7][22]
Несколько сообщений предположили, что STING связан с активацией избирательной аутофагии.[12] Микобактерии туберкулеза было показано, что они продуцируют цитозольные лиганды ДНК, которые активируют STING, что приводит к убиквитинирование бактерий и последующее пополнение аутофагия родственные белки, все из которых необходимы для «селективного» нацеливания на аутофагию и врожденной защиты от М. туберкулез.[23]
Таким образом, STING координирует множественные иммунные ответы на инфекцию, включая индукцию интерферонов и STAT6-зависимый ответ, а также избирательный ответ аутофагии.[7]
Как цитозольный датчик ДНК
Циклические динуклеотиды -вторичные сигнальные молекулы, продуцируемые различными видами бактерий, были обнаружены в цитозоле клеток млекопитающих во время внутриклеточной патогенной инфекции; это приводит к активации TBK1 -IRF3 и последующее производство интерферон I типа.[7][24]Было показано, что STING напрямую связывается с циклический ди-GMP, и это признание приводит к производству цитокины, Такие как интерферон I типа, которые необходимы для успешного устранения патогенов.[25]
В качестве сигнального адаптера
DDX41, член семейства геликаз DEXDc, в миелоидных дендритных клетках распознает внутриклеточную ДНК и опосредует врожденный иммунный ответ через прямую ассоциацию с STING.[26] Другие датчики ДНК- DAI, РНК-полимераза III, IFI16, также было показано, что они активируют СТИНГ посредством прямого или косвенного взаимодействия.[14]
Циклическая GMP-AMP-синтаза (cGAS), который принадлежит к семейству нуклеотидилтрансфераз, способен распознавать содержимое цитозольной ДНК и вызывать STING-зависимый интерфероновый ответ, продуцируя вторичный мессенджер. циклический гуанозинмонофосфат-аденозинмонофосфат (циклический GMP-AMP или cGAMP). После циклический GMP-AMP связанный СТИНГ активируется, усиливает TBK1 активность по фосфорилированию IRF3 и STAT6 для нисходящего потока интерферон I типа отклик.[27][28]
Было высказано предположение, что внутриклеточный кальций играет важную роль в ответе пути STING.[29]
Рекомендации
- ^ а б c ENSG00000288243 GRCh38: Ensembl release 89: ENSG00000184584, ENSG00000288243 - Ансамбль, Май 2017
- ^ а б c GRCm38: выпуск Ensembl 89: ENSMUSG00000024349 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ "Ссылка на Mouse PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ "STING1, стимулятор интерферонового ответа cGAMP interctor 1 [Homo sapiens (человек)]".
- ^ Nakhaei P, Hiscott J, Lin R (июнь 2010 г.). «Укус противовирусного пути». Журнал молекулярной клеточной биологии. 2 (3): 110–2. Дои:10.1093 / jmcb / mjp048. PMID 20022884.
- ^ а б c d е ж Burdette DL, Vance RE (январь 2013 г.). «STING и врожденный иммунный ответ на нуклеиновые кислоты в цитозоле». Иммунология природы. 14 (1): 19–26. Дои:10.1038 / ni.2491. PMID 23238760. S2CID 7968532.
- ^ Шу Ц., Йи Г, Уоттс Т., Као С.Ч., Ли П (июль 2012 г.). «Структура STING, связанного с циклическим ди-GMP, раскрывает механизм распознавания циклических динуклеотидов иммунной системой». Структурная и молекулярная биология природы. 19 (7): 722–4. Дои:10.1038 / nsmb.2331. ЧВК 3392545. PMID 22728658.
- ^ «Профиль экспрессии EST TMEM173». biogps org. biogps.org.
- ^ "NCBI TMEM173 выражение GEOprofile". NCBI. www.ncbi.nlm.nih.gov/geoprofiles.
- ^ а б Исикава Х., Парикмахер Г.Н. (октябрь 2008 г.). «STING - это адаптер эндоплазматического ретикулума, который способствует передаче сигналов врожденного иммунитета». Природа. 455 (7213): 674–8. Дои:10.1038 / природа07317. ЧВК 2804933. PMID 18724357.
- ^ а б Сайто Т., Фудзита Н., Хаяси Т., Такахара К., Сато Т., Ли Х, Мацунага К., Кагеяма С., Омори Х, Нода Т., Ямамото Н., Кавай Т., Исии К., Такеучи О, Йошимори Т., Акира С. (декабрь 2009 г. ). «Atg9a контролирует управляемую дцДНК динамическую транслокацию STING и врожденный иммунный ответ». Труды Национальной академии наук Соединенных Штатов Америки. 106 (49): 20842–6. Дои:10.1073 / pnas.0911267106. ЧВК 2791563. PMID 19926846.
- ^ а б Парикмахер Г.Н. (февраль 2011 г.). «Врожденные иммунные пути восприятия ДНК: STING, AIMII и регуляция выработки интерферона и воспалительных реакций». Текущее мнение в иммунологии. 23 (1): 10–20. Дои:10.1016 / j.coi.2010.12.015. ЧВК 3881186. PMID 21239155.
- ^ а б Китинг С.Е., Баран М., Боуи АГ (декабрь 2011 г.). «Цитозольные ДНК-сенсоры, регулирующие индукцию интерферона I типа» (PDF). Тенденции в иммунологии. 32 (12): 574–81. Дои:10.1016 / j.it.2011.08.004. HDL:2262/68041. PMID 21940216.
- ^ Ma Z, Damania B (февраль 2016 г.). «Путь защиты cGAS-STING и противодействие ему вирусами». Клеточный хозяин и микроб. 19 (2): 150–8. Дои:10.1016 / j.chom.2016.01.010. ЧВК 4755325. PMID 26867174.
- ^ Xie J, Li Y, Shen X, Got G, Zhu Y, Cui J, Wang L, Shi Z, Zhou P (март 2018 г.). "Снижение активации STING-зависимого интерферона у летучих мышей". Клеточный хозяин и микроб. 23 (3): 297–301.e4. Дои:10.1016 / j.chom.2018.01.006. ЧВК 7104992. PMID 29478775.
- ^ а б Джин Л., Гетахун А., Ноулз Х.М., Моган Дж., Акерлунд Л.Дж., Паккард Т.А., Перро А.Л., Камбье Дж.С. (март 2013 г.). «STING / MPYS опосредует защиту хозяина против инфекции Listeria monocytogenes, регулируя миграцию моноцитов Ly6C (hi)». Журнал иммунологии. 190 (6): 2835–43. Дои:10.4049 / jimmunol.1201788. ЧВК 3593745. PMID 23378430.
- ^ Вудвард Дж. Дж., Явароне А. Т., Портной Д. А. (июнь 2010 г.). «c-ди-АМФ, секретируемый внутриклеточными Listeria monocytogenes, активирует реакцию хозяина на интерферон I типа». Наука. 328 (5986): 1703–5. Дои:10.1126 / science.1189801. ЧВК 3156580. PMID 20508090.
- ^ Исикава Х., Ма З., Парикмахер Г.Н. (октябрь 2009 г.). "STING регулирует внутриклеточный ДНК-опосредованный интерферон-зависимый врожденный иммунитет типа I". Природа. 461 (7265): 788–92. Дои:10.1038 / природа08476. ЧВК 4664154. PMID 19776740.
- ^ а б Sharma S, DeOliveira RB, Kalantari P, Parroche P, Goutagny N, Jiang Z, Chan J, Bartholomeu DC, Lauw F, Hall JP, Barber GN, Gazzinelli RT, Fitzgerald KA, Golenbock DT (август 2011 г.). «Врожденное иммунное распознавание АТ-богатого мотива ДНК« стебель-петля »в геноме Plasmodium falciparum». Иммунитет. 35 (2): 194–207. Дои:10.1016 / j.immuni.2011.05.016. ЧВК 3162998. PMID 21820332.
- ^ а б Мансанильо П.С., Шайло М.Ю., Портной Д.А., Cox JS (май 2012 г.). «Mycobacterium tuberculosis активирует ДНК-зависимый путь цитозольного наблюдения в макрофагах». Клеточный хозяин и микроб. 11 (5): 469–80. Дои:10.1016 / j.chom.2012.03.007. ЧВК 3662372. PMID 22607800.
- ^ Чен Х, Сун Х, Ю Ф, Сунь В, Чжоу Х, Чен Л, Ян Дж, Ван И, Тан Х, Гуань И, Ся В, Гу Дж, Исикава Х, Гутман Д., Парикмахер Г, Цинь З, Цзян З. (Октябрь 2011 г.). «Активация STAT6 с помощью STING имеет решающее значение для противовирусного врожденного иммунитета». Клетка. 147 (2): 436–46. Дои:10.1016 / j.cell.2011.09.022. PMID 22000020.
- ^ Уотсон Р.О., Мансанильо П.С., Кокс Д.С. (август 2012 г.). «Внеклеточная ДНК M. tuberculosis нацелена на бактерии для аутофагии, активируя путь определения ДНК хозяина». Клетка. 150 (4): 803–15. Дои:10.1016 / j.cell.2012.06.040. ЧВК 3708656. PMID 22901810.
- ^ McWhirter SM, Barbalat R, Monroe KM, Fontana MF, Hyodo M, Joncker NT, Ishii KJ, Akira S, Colonna M, Chen ZJ, Fitzgerald KA, Hayakawa Y, Vance RE (август 2009 г.). «Реакция хозяина на интерферон I типа индуцируется цитозольным зондированием бактериального вторичного мессенджера cyclic-di-GMP». Журнал экспериментальной медицины. 206 (9): 1899–911. Дои:10.1084 / jem.20082874. ЧВК 2737161. PMID 19652017.
- ^ Burdette DL, Monroe KM, Sotelo-Troha K, Iwig JS, Eckert B., Hyodo M, Hayakawa Y, Vance RE (октябрь 2011 г.). «STING - это датчик прямого врожденного иммунитета циклического ди-GMP». Природа. 478 (7370): 515–8. Дои:10.1038 / природа10429. ЧВК 3203314. PMID 21947006.
- ^ Чжан З., Юань Б., Бао М., Лу Н, Ким Т., Лю Ю.Дж. (октябрь 2011 г.). «Хеликаза DDX41 воспринимает внутриклеточную ДНК, опосредованную адаптером STING в дендритных клетках». Иммунология природы. 12 (10): 959–65. Дои:10.1038 / ni.2091. ЧВК 3671854. PMID 21892174.
- ^ Ву Дж, Сун Л., Чен Х, Ду Ф, Ши Х, Чен С., Чен З. Дж. (Февраль 2013 г.). «Циклический GMP-AMP является вторичным эндогенным посредником в передаче сигналов врожденного иммунитета цитозольной ДНК». Наука. 339 (6121): 826–30. Дои:10.1126 / science.1229963. ЧВК 3855410. PMID 23258412.
- ^ Сунь Л., Ву Дж, Ду Ф, Чен Х, Чен ЗДж (февраль 2013 г.). «Циклическая GMP-AMP-синтаза представляет собой цитозольный ДНК-сенсор, который активирует путь интерферона I типа». Наука. 339 (6121): 786–91. Дои:10.1126 / science.1232458. ЧВК 3863629. PMID 23258413.
- ^ Ким С., Кох П., Ли Л., Пешкин Л., Митчисон Т. Дж. (4 июня 2017 г.). «Доказательства роли кальция в передаче сигналов STING». bioRxiv 10.1101/145854.
дальнейшее чтение
- Ван И, Тонг Х, Омореги Э.С., Лю В., Мэн С., Е Икс (октябрь 2012 г.). «Тетраспанин 6 (TSPAN6) отрицательно регулирует иммунную сигнализацию, индуцируемую ретиноевой кислотой, опосредованную I-подобным рецептором геном, зависимым от убиквитинирования образом». Журнал биологической химии. 287 (41): 34626–34. Дои:10.1074 / jbc.M112.390401. ЧВК 3464568. PMID 22908223.
- Инь Цюй, Тиан И, Кабалисваран В, Цзян Х, Ту Д, Эк MJ, Чен Ц.Дж., Ву Х (июнь 2012 г.). «Циклическое определение ди-GMP с помощью сигнального белка врожденного иммунитета STING». Молекулярная клетка. 46 (6): 735–45. Дои:10.1016 / j.molcel.2012.05.029. ЧВК 3697849. PMID 22705373.
- Агирре С., Маэстре А.М., Пагни С., Патель Дж. Р., Сэвидж Т., Гутман Д., Маринджер К., Бернал-Рубио Д., Шабман Р. С., Саймон В., Родригес-Мадос Дж. Р., Малдер Л. К., Парикмахер Г. Н., Фернандес-Сесма А. (2012) . «DENV подавляет продукцию IFN типа I в инфицированных клетках путем расщепления STING человека». Патогены PLOS. 8 (10): e1002934. Дои:10.1371 / journal.ppat.1002934. ЧВК 3464218. PMID 23055924.
- Ли И, Ли С, Сюэ П, Чжун Б., Мао А.П., Ран И, Чен Х, Ван ИЙ, Ян Ф, Шу Х.В. (май 2009 г.). «ISG56 является регулятором отрицательной обратной связи вирусной передачи сигналов и клеточного противовирусного ответа». Труды Национальной академии наук Соединенных Штатов Америки. 106 (19): 7945–50. Дои:10.1073 / pnas.0900818106. ЧВК 2683125. PMID 19416887.
- Конлон Дж., Бёрдетт Д.Л., Шарма С., Бхат Н., Томпсон М., Цзян З., Ратинам В.А., Монахи Б., Джин Т., Сяо Т.С., Фогель С.Н., Вэнс Р.Э., Фицджеральд К.А. (май 2013 г.). «Мышь, но не человек STING, связывается и передает сигнал в ответ на агент, разрушающий сосуды, 5,6-диметилксантенон-4-уксусную кислоту». Журнал иммунологии. 190 (10): 5216–25. Дои:10.4049 / jimmunol.1300097. ЧВК 3647383. PMID 23585680.
- Абэ Т., Харашима А., Ся Т., Конно Х, Конно К., Моралес А., Ан Дж., Гутман Д., Барбер Г. Н. (апрель 2013 г.). «Распознавание STING цитоплазматической ДНК запускает клеточную защиту». Молекулярная клетка. 50 (1): 5–15. Дои:10.1016 / j.molcel.2013.01.039. ЧВК 3881179. PMID 23478444.
- Назми А., Мукхопадхьяй Р., Датта К., Басу А. (2012). «STING опосредует врожденный иммунный ответ нейронов после инфицирования вирусом японского энцефалита». Научные отчеты. 2: 347. Дои:10.1038 / srep00347. ЧВК 3317237. PMID 22470840.
- Чжан Дж., Ху ММ, Ван Й.Й., Шу HB (август 2012 г.). «Белок TRIM32 модулирует индукцию интерферона I типа и клеточный противовирусный ответ, воздействуя на белок MITA / STING для K63-связанного убиквитинирования». Журнал биологической химии. 287 (34): 28646–55. Дои:10.1074 / jbc.M112.362608. ЧВК 3436586. PMID 22745133.
- Исикава Х., Парикмахер Г.Н. (октябрь 2008 г.). «STING - это адаптер эндоплазматического ретикулума, который способствует передаче сигналов врожденного иммунитета». Природа. 455 (7213): 674–8. Дои:10.1038 / природа07317. ЧВК 2804933. PMID 18724357.