Гетерозис - Heterosis
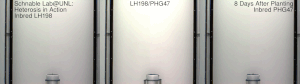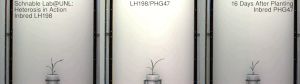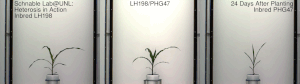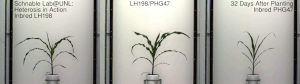
Гетерозис, гибридная сила, или улучшение аутбридинга улучшенная или усиленная функция любого биологического качества в гибридный потомство. Потомство гетеротично, если его черты усиливаются в результате смешения генетического вклада его родителей. Эти эффекты могут быть вызваны Менделевский или неменделирующее наследование.
Определения
Предлагая термин гетерозис заменить старый термин гетерозиготный, G.H. Шулл направлена на то, чтобы избежать ограничения этого термина эффектами, которые можно объяснить гетерозиготностью в менделевском наследовании.[1]
Физиологическая жизнеспособность организма, проявляющаяся в его скорости роста, его высоте и общей устойчивости, положительно коррелирует со степенью несходства гамет, объединением которых был образован организм ... Чем больше различий между объединяющими гаметами - при по крайней мере в определенных пределах - тем больше в целом степень стимуляции ... Эти различия не обязательно должны быть менделевскими по своей наследственности ... Чтобы избежать вывода о том, что все генотипические различия, которые стимулируют деление клеток, рост и другие физиологические действия организма, являются Менделирующие по своей наследственности, а также для краткости выражения я предлагаю ... принять слово «гетерозис».
Гетерозис часто рассматривается как противоположность инбридинговая депрессия хотя различия в этих двух концепциях можно увидеть в эволюционных соображениях, таких как роль генетическая вариация или эффекты генетический дрейф в небольших популяциях по этим понятиям. Инбридинговая депрессия возникает, когда у родственных родителей есть дети с черты которые негативно влияют на их фитнес во многом благодаря гомозиготность. В таких случаях ауткроссинг должен привести к гетерозису.
Не все скрещивания приводят к гетерозису. Например, когда гибрид наследует признаки от своих родителей, которые не полностью совместимы, приспособленность может снизиться. Это форма аутбредная депрессия.
Доминирование против чрезмерного доминирования
Доминирование против чрезмерного доминирования это научная полемика в области генетика это сохраняется более века.[2] Эти две альтернативные гипотезы были впервые высказаны в 1908 году.
Генетическая основа
Когда популяция небольшая или инбредная, она имеет тенденцию терять генетическое разнообразие. Инбридинговая депрессия потеря приспособленности из-за потери генетического разнообразия. Инбредные штаммы обычно гомозиготный для рецессивный аллели которые являются умеренно вредными (или производят признак, нежелательный с точки зрения заводчика). С другой стороны, гетерозис или гибридная сила - это тенденция беспородных пород превосходить обоих инбредных родителей по пригодности.
Селективное разведение растений и животных, включая гибридизацию, началось задолго до того, как появилось понимание основных научных принципов. В начале 20 века после Законы Менделя Чтобы понять и принять, генетики взялись объяснить превосходную силу многих гибридов растений. Были разработаны две конкурирующие гипотезы, которые не исключают друг друга:[3]
- Гипотеза доминирования. Гипотеза доминирования приписывает превосходство гибридов подавлению нежелательных рецессивных аллелей одного родителя доминантными аллелями другого. Он объясняет низкую производительность инбредных штаммов потерей генетического разнообразия, когда штаммы становятся чисто гомозиготными по многим локусам. Гипотеза доминирования была впервые высказана в 1908 году генетиком. Чарльз Давенпорт.[4] Согласно гипотезе доминантности вредные аллели, как ожидается, будут поддерживаться в популяции случайного спаривания при балансе выбора-мутации, который будет зависеть от скорости мутации, эффекта аллелей и степени экспрессии аллелей в гетерозиготах.[5]
- Гипотеза сверхдоминирования. Определенные комбинации аллелей, которые могут быть получены при скрещивании двух инбредных линий: выгодно в гетерозиготе. Гипотеза сверхдоминирования приписывает преимущество гетерозигот выживанию многих аллелей, которые являются рецессивными и вредными для гомозигот. Он объясняет низкую производительность инбредных штаммов высоким процентом этих вредных рецессивов. Гипотеза сверхдоминирования была независимо разработана Эдвард М. Ист (1908)[6] и Джордж Шулль (1908).[7] Ожидается, что генетическая изменчивость в преобладающем локусе будет поддерживаться балансирующим отбором. Высокая пригодность гетерозиготных генотипов способствует сохранению аллельного полиморфизма в популяции.[5]
Доминирование и сверхдоминирование имеют разные последствия для профиль экспрессии гена лиц. Если преобладание является основной причиной преимуществ гетерозиса по пригодности, то у гетерозиготного потомка должна быть сверхэкспрессия определенных генов по сравнению с гомозиготными родителями. С другой стороны, если причиной является доминирование, у гетерозиготного потомства должно быть меньше экспрессии генов по сравнению с родителями. Кроме того, для любого данного гена экспрессия должна быть сравнима с экспрессией, наблюдаемой у установщика двух родителей.
Историческая ретроспектива
Популяционный генетик Джеймс Кроу (1916–2012) в молодости считал, что чрезмерное доминирование является основным фактором жизнеспособности гибридов. В 1998 году он опубликовал ретроспективный обзор развивающейся науки.[8] По словам Кроу, демонстрация нескольких случаев преимущество гетерозиготы у Drosophila и других организмов теория сверхдоминирования впервые вызвала большой энтузиазм среди ученых, изучающих гибридизацию растений. Но чрезмерное доминирование подразумевает, что урожайность инбредной линии должна снижаться по мере того, как инбредные линии отбираются для работы их гибридных скрещиваний, поскольку доля вредных рецессивов в инбредной популяции возрастает. На протяжении многих лет эксперименты в области генетики растений доказали, что происходит обратное, что приводит к увеличению урожайности как инбредных линий, так и гибридов, предполагая, что одного доминирования может быть достаточно для объяснения более высокой урожайности гибридов. Сообщалось лишь о нескольких убедительных случаях чрезмерного доминирования во всей генетике. С 1980-х годов, когда появились экспериментальные данные, теория доминирования вернулась.
Ворона писал:
Текущая точка зрения ... заключается в том, что гипотеза доминирования является основным объяснением снижения инбридинга и высокой урожайности гибридов. Статистических свидетельств влияния чрезмерного доминирования и эпистаз. Но остается открытым вопрос, получают ли лучшие гибриды дополнительный импульс от чрезмерного доминирования или благоприятного эпистатического вклада.[8]
Полемика
Термин гетерозис часто вызывает путаницу и даже споры, особенно при селективном разведении домашние животные, потому что иногда (ошибочно) утверждается, что все помесь растения и животные «генетически превосходят» своих родителей из-за гетерозиса[нужна цитата ]. Однако у этого утверждения есть две проблемы:
- Во-первых, согласно статье, опубликованной в журнале Геномная биология, "генетическое превосходство "- это плохо определенный термин и не является общепринятой терминологией в научной области генетики.[9] Родственный термин фитнес хорошо определен, но его редко можно измерить напрямую. Вместо этого ученые используют объективные измеримые величины, такие как количество семян, которые производит растение, скорость прорастания семян или процент организмов, доживающих до репродуктивного возраста.[10] С этой точки зрения помесь растения и животные, демонстрирующие гетерозис, могут обладать «превосходными» чертами, но это не обязательно приравнивается к какому-либо доказательству явного «генетического превосходства». Использование термина «превосходство» является обычным явлением, например, в селекции сельскохозяйственных культур, где хорошо понимают, что он означает более урожайное и более устойчивое растение для сельского хозяйства. Такое растение может давать более высокие урожаи на ферме, но, вероятно, будет бороться за выживание в дикой природе, что делает его использование открытым для неправильного толкования. В генетике человека любой вопрос о «генетическом превосходстве» становится еще более проблематичным из-за исторических и политических последствий любого такого утверждения. Некоторые могут даже пойти дальше и назвать это сомнительным оценочным суждением в области политики, а не науки.[9]
- Во-вторых, не все гибриды проявляют гетерозис (см. аутбредная депрессия ).
Примером неоднозначных оценочных суждений, налагаемых на гибриды и гибридную силу, является мул. Хотя мулы почти всегда бесплодны, их ценят за сочетание выносливости и темперамента, которые отличаются от их родителей - лошадей или ослов. Хотя эти качества могут сделать их «превосходными» для конкретных целей использования людьми, проблема бесплодия подразумевает, что эти животные, скорее всего, вымрут без вмешательства человека через животноводство, что делает их «низшими» с точки зрения естественный отбор.
Генетические и эпигенетические основы
С начала 1900-х годов были разработаны две конкурирующие генетические гипотезы, не обязательно взаимоисключающие, для объяснения жизнеспособности гибридов. Совсем недавно также был установлен эпигенетический компонент жизнеспособности гибридов.[11][12]
Генетический гипотеза доминирования объясняет превосходство гибридов маскировкой экспрессии нежелательных (вредных) рецессивных аллелей от одного родителя доминантными (обычно дикого типа) аллелями от другого (см. Комплементация (генетика) ). Он приписывает низкую производительность инбредных штаммов экспрессии гомозиготных вредных рецессивных аллелей. Генетический гипотеза сверхдоминирования утверждает, что некоторые комбинации аллелей (которые могут быть получены путем скрещивания двух инбредных штаммов) особенно выгодно в паре с гетерозиготной особью. Эта гипотеза обычно используется для объяснения устойчивости некоторых аллелей (наиболее известный из них: Серповидно-клеточная характеристика аллель), которые вредны для гомозигот. В нормальных условиях такие вредные аллели будут удалены из популяции в процессе естественного отбора. Подобно гипотезе доминирования, он приписывает плохие характеристики инбредных линий экспрессии таких вредных рецессивных аллелей. В любом случае ауткроссное скрещивание дает преимущество маскировки вредных рецессивных аллелей в потомстве. Было предложено, чтобы это преимущество было основным фактором в поддержании полового размножения среди эукариот, как обобщено в статье. Эволюция полового размножения.
Установлен эпигенетический вклад в гетерозис у растений,[12] и это также было зарегистрировано у животных.[13] МикроРНК (miRNA), обнаруженные в 1993 году, представляют собой класс некодирующих малых РНК, которые подавляют трансляцию информационных РНК (мРНК) или вызывают деградацию мРНК.[14] В гибридных растениях большинство miRNA имеют неаддитивную экспрессию (она может быть выше или ниже, чем уровни у родителей).[12] Это говорит о том, что малые РНК участвуют в росте, силе и адаптации гибридов.[12]
Влияние «гетерозиса без гибридности» на размер растений было продемонстрировано на генетически изогенных триплоидных (автополиплоидных) растениях F1, где избыток триплоидов F1 в отцовском геноме показывает положительный гетерозис, тогда как избыток F1 в материнском геноме демонстрирует отрицательные эффекты гетерозиса.[15] Такие данные демонстрируют, что эффекты гетерозиса с зависимой от дозировки геномной эпигенетической основой могут возникать в потомстве F1, которое является генетически изогенным (то есть не имеет гетерозиготности).[15][16] Было показано[11] эта гибридная сила в аллополиплоидном гибриде двух Арабидопсис Этот вид был обусловлен эпигенетическим контролем в вышележащих областях двух генов, что вызвало серьезные последующие изменения в накоплении хлорофилла и крахмала. Механизм включает ацетилирование и / или метилирование определенных аминокислот в гистон H3, белок, тесно связанный с ДНК, который может либо активировать, либо репрессировать связанные гены.
Главный комплекс гистосовместимости у животных
Одним из примеров того, где определенные гены могут быть важны для гетерозиса у позвоночных животных, является главный комплекс гистосовместимости (MHC). Позвоночные животные наследуют несколько копий обоих MHC класс I и MHC класс II от каждого родителя, которые используются в презентация антигена как часть адаптивной иммунной системы. Каждая отдельная копия генов способна связывать и представлять различный набор потенциальных пептидов для Т-лимфоциты. Эти гены очень полиморфны во всех популяциях, но будут более похожими в меньших, более близких популяциях. Селекция между более генетически далекими индивидуумами снизит вероятность наследования двух одинаковых или похожих аллелей, что позволит представить более разнообразный диапазон пептидов. Таким образом, это снижает вероятность того, что какой-либо конкретный патоген не будет распознаваться, и означает, что больше антигенных белков любого патогена, вероятно, будет распознаваться, что дает больший диапазон активации Т-клеток и, следовательно, больший ответ. Это также будет означать, что иммунитет, приобретенный против патогена, будет направлен против более широкого спектра антигенов, а это означает, что патоген должен мутировать больше до потери иммунитета. Таким образом, гибриды с меньшей вероятностью будут подвержены патогенным заболеваниям и будут более способны бороться с инфекцией.
Растения
Кресты между инбредами от разных гетеротические группы приводят к сильным гибридам F1 со значительно большим гетерозисом, чем гибриды F1 от инбредов в пределах одной и той же гетеротической группы или образца. Селекционеры растений создают гетеротические группы для классификации инбредных линий, и их можно постепенно улучшать путем реципрокного рекурсивного отбора.
Гетерозис используется для повышения урожайности, однородности и урожайности. Методы гибридного разведения используются в кукуруза, сорго, рис, сахарная свекла, лук, шпинат, подсолнухи, брокколи и создать более психоактивный каннабис.
Кукуруза (кукуруза)
Почти вся полевая кукуруза (кукуруза ) выращивается в большинстве развитые страны проявляет гетерозис. Современные гибриды кукурузы значительно превосходят традиционные сорта и лучше реагируют на удобрение.
Гетерозис кукурузы был хорошо продемонстрирован в начале 20 века Джордж Х. Шулль и Эдвард М. Ист после того, как гибридная кукуруза была изобретена доктором Уильям Джеймс Бил из Университет штата Мичиган на основе работы, начатой в 1879 г. по настоянию Чарльз Дарвин. Работа доктора Била привела к первому опубликованному отчету о полевой эксперимент демонстрируя энергию гибрида кукурузы, Юджин Давенпорт и Перри Холден, 1881. Эти различные пионеры ботаники и смежных областей показали, что скрещивания инбредных линий, полученные из южной вмятины и северного кремня, соответственно, показали значительный гетерозис и превзошли урожай традиционных сортов той эпохи. Однако в то время такие гибриды не могли быть экономически выгодны для массового использования фермерами. Дональд Ф. Джонс на Коннектикут Сельскохозяйственная опытная станция, Новый рай изобрел первый практический метод выращивания высокоурожайной гибридной кукурузы в 1914–1917 гг. Метод Джонса привел к получению гибрида двойного скрещивания, который требует двух этапов скрещивания, основанных на четырех различных исходных инбредных линиях. Более поздняя работа селекционеров кукурузы позволила получить инбредные линии, обладающие достаточной энергией для практического получения коммерческого гибрида за один этап, - гибриды одинарного скрещивания. Гибриды одиночного кросса производятся всего от двух исходных родительских инбредов. Как правило, они более сильны и однороднее, чем более ранние гибриды двойного скрещивания. Процесс создания этих гибридов часто включает удаление метелок.
Гибриды кукурузы умеренного пояса происходят от двух основных гетеротических групп: Айова с жестким стеблем и синтетическим стеблем.[нужна цитата ]
Рис (Oryza sativa)
Производство риса в Китае резко выросло из-за интенсивного использования гибридного риса. В Китае в результате усилий был получен супергибридный сорт риса (LYP9) с производительностью ~ 15 тонн с гектара. В Индии также несколько сортов показали высокую жизнеспособность, в том числе RH-10 и Suruchi 5401.
Гибридный скот
Концепция гетерозиса также применяется в производстве коммерческих домашний скот. У крупного рогатого скота скрещивает Блэк Ангус и Херефорд произвести крест, известный как "Черный лысый ". В свинья, "синие окурки" производятся крестом Хэмпшир и Йоркшир. Другие, более экзотические гибриды, такие как "гибрид коровы и бизона "также используются для специализированных рынков.
Домашняя птица
В пределах домашняя птица, связанный с полом гены были использованы для создания гибридов, в которых самцы и самки могут быть отсортированы по цвету однодневного возраста. Конкретные гены, используемые для этого, - это гены, отвечающие за рост оперения и роста крыльев. Кресты такого типа создают то, что продается как черные секс-ссылки, красные секс-ссылки и различные другие кресты, известные под торговыми названиями.
Коммерческий бройлеры производятся путем скрещивания различных сортов White Rocks и White Cornish, Cornish обеспечивает большую структуру, а Rocks обеспечивает высокую скорость увеличения. Полученная сила гибрида позволяет получать однородных птиц с товарной тушкой в возрасте 6–9 недель.
Аналогичным образом, гибриды между различными сортами белого леггорна используются для получения стад-несушек, которые обеспечивают большую часть белых растений. яйца для продажи в США.
Собаки
В 2013 году исследование показало, что смешанные породы живут в среднем на 1,2 года дольше, чем чистопородные.[17]
Джон Скотт и Джон Л. Фуллер провели детальное исследование породистых кокер-спаниелей, породистых басенджи и их гибридов.[18]Они обнаружили, что гибриды работают быстрее, чем любой из родителей, возможно, из-за гетерозиса. Другие характеристики, такие как базальная частота сердечных сокращений, не демонстрировали какого-либо гетерозиса - базальная частота сердечных сокращений собаки была близка к средней для ее родителей - возможно, из-за аддитивных эффектов нескольких генов.[19]
Иногда люди, работающие над программой разведения собак, не находят полезного гетерозиса.[20]
Птицы
В 2014 году исследование, проведенное Центром интегративной экологии в г. Университет Дикина в Джилонге Виктория пришла к выводу, что внутривидовые гибриды между подвидами Flaveolus и elegans из Малиновая розелла (Platycercus elegans) были более склонны бороться с болезнями, чем их чистые аналоги.[21]
Люди
Все люди чрезвычайно генетически похожи друг на друга.[22][23][24] Майкл Мингрони предложил гетерозис в форме гибридной силы, связанной с историческим сокращением уровней инбридинга, как объяснение Эффект Флинна, устойчивый рост показателей IQ во всем мире в течение двадцатого века.
Смотрите также
использованная литература
- ^ Джордж Харрисон Шулл (1948). "Что такое" гетерозис "?". Генетика. 33 (5): 439–446. ЧВК 1209417. PMID 17247290.
- ^ Birchler J.A .; Auger D.L .; Загадка Н.С. (2003). «В поисках молекулярных основ гетерозиса». Растительная клетка. 15 (10): 2236–2239. Дои:10.1105 / tpc.151030. ЧВК 540269. PMID 14523245.
- ^ Ворона, Джеймс Ф. (1948). «Альтернативные гипотезы гибридной силы». Генетика. 33 (5): 477–487. ЧВК 1209419. PMID 17247292.
- ^ Давенпорт CB (1908). «Дегенерация, альбинизм и инбридинг». Наука. 28 (718): 454–5. Bibcode:1908Sci .... 28..454D. Дои:10.1126 / science.28.718.454-b. PMID 17771943.
- ^ а б Карр, Дэвид Э .; Дудаш, Мишель Р. (29.06.2003). «Современные подходы к генетическим основам инбридинговой депрессии у растений». Философские труды Королевского общества B: биологические науки. 358 (1434): 1071–1084. Дои:10.1098 / rstb.2003.1295. ISSN 0962-8436. ЧВК 1693197. PMID 12831473.
- ^ Восток EM (1908). «Инбридинг в кукурузе». Отчеты сельскохозяйственной экспериментальной станции Коннектикута за 1907 год: 419–428.
- ^ Шулль Г. Х. (1908). «Состав кукурузного поля». Отчеты Американской ассоциации заводчиков: 296–301.
- ^ а б Ворона, Джеймс Ф. (1998). «90 лет назад: начало гибридной кукурузы». Генетика. 148 (3): 923–928. ЧВК 1460037. PMID 9539413.
- ^ а б Риш Н., Бурчард Э, Зив Э, Тан Х (июль 2002 г.). «Категоризация людей в биомедицинских исследованиях: гены, раса и болезни». Геном Биол. 3 (7): comment2007. Дои:10.1186 / gb-2002-3-7-comment2007. ЧВК 139378. PMID 12184798.
- ^ Веллер С.Г., Сакаи А.К., Тайский Д.А., Том Дж., Ранкин А.Е. (ноябрь 2005 г.). «Инбридинговая депрессия и гетерозис в популяциях Schiedea viscosa, высоко самоопыляющегося вида». J. Evol. Биол. 18 (6): 1434–44. Дои:10.1111 / j.1420-9101.2005.00965.x. PMID 16313456.
- ^ а б Ni Z, Kim ED, Ha M, et al. (Январь 2009 г.). «Измененные циркадные ритмы регулируют силу роста у гибридов и аллополиплоидов». Природа. 457 (7227): 327–31. Bibcode:2009Натура.457..327Н. Дои:10.1038 / природа07523. ЧВК 2679702. PMID 19029881.
- ^ а б c d Баранвал В.К., Миккилинени В., Зер У.Б., Тяги А.К., Капур С. (ноябрь 2012 г.). «Гетерозис: новые идеи о гибридной силе». J. Exp. Бот. 63 (18): 6309–14. Дои:10.1093 / jxb / ers291. PMID 23095992.
- ^ Хан З., Мтанго Н. Р., Патель Б. Г., Сапиенца С., Латам К. Э. (октябрь 2008 г.). «Гибридная сила и трансгенерационные эпигенетические эффекты на фенотип ранних эмбрионов мыши». Биол. Репрод. 79 (4): 638–48. Дои:10.1095 / биолрепрод.108.069096. ЧВК 2844494. PMID 18562704.
- ^ Чжоу Й., Фергюсон Дж., Чанг Дж. Т., Клугер Й. (2007). «Меж- и внутрикомбинаторная регуляция факторами транскрипции и микроРНК». BMC Genomics. 8: 396. Дои:10.1186/1471-2164-8-396. ЧВК 2206040. PMID 17971223.
- ^ а б Форт, Антуан; Райдер, Питер; Маккеун, Питер С .; Wijnen, Cris; Аартс, Марк Г .; Сюльпис, Ронан; Спиллейн, Чарльз (01.01.2016). «Дезагрегация полиплоидии, дозировка родительского генома и вклад гибридности в гетерозис у Arabidopsis thaliana». Новый Фитолог. 209 (2): 590–599. Дои:10.1111 / nph.13650. ISSN 1469-8137. PMID 26395035.
- ^ Душинская, Дорота; Маккеун, Питер С .; Juenger, Thomas E .; Пьетрашевская-Богель, Анна; Гилен, Дэнни; Спиллейн, Чарльз (2013-04-01). «Плодовитость гамет и изменение числа семяпочек в самоопыленных реципрокных триплоидных растениях F1 являются наследственными и проявляют эпигенетические эффекты родительского происхождения». Новый Фитолог. 198 (1): 71–81. Дои:10.1111 / nph.12147. ISSN 1469-8137. PMID 23368793.
- ^ O’Neill, D.G .; Чёрч, Д. Б .; McGreevy, P.D .; Thomson, P.C .; Бродбельт, Д. К. (2013). «Долголетие и смертность домашних собак в Англии» (PDF). Ветеринарный журнал. 198 (3): 638–43. Дои:10.1016 / j.tvjl.2013.09.020. PMID 24206631.
- ^ Спеди, Тайрон С .; Острандер, Элейн А. (2008). «Поведенческая генетика собак: определение фенотипов и накопление генов». Американский журнал генетики человека. 82: 10–8. Дои:10.1016 / j.ajhg.2007.12.001. ЧВК 2253978. PMID 18179880.
- ^ Джон Пол Скотт и Джон Л. Фуллер. «Генетика и социальное поведение собаки». 1965. с. 307 и стр. 313.
- ^ Пер Дженсен. «Поведенческая биология собак». 2007. с. 179
- ^ Австралийская география (сентябрь 2014 г.). «Гибридные птицы лучше борются с болезнями, чем чистокровные».
- ^ Ястребы, Джон (2013). Значение геномов неандертальцев и денисовцев в эволюции человека. Ежегодный обзор антропологии. 42. Ежегодные обзоры. С. 433–449, 438. Дои:10.1146 / annurev-anthro-092412-155548. ISBN 978-0-8243-1942-7. ISSN 0084-6570.
Общая эволюционная история живых людей привела к высокой степени родства среди всех живых людей, на что указывает, например, очень низкий индекс фиксации (FST) среди живых человеческих популяций.
- ^ Барбужани, Гвидо; Колонна, Винченца (15 сентября 2011 г.). «Глава 6: Генетические основы биоразнообразия человека: обновленная информация». В Zachos, Frank E .; Хабель, Ян Кристиан (ред.). Горячие точки биоразнообразия: распределение и защита приоритетных территорий сохранения. Springer. С. 97–119. Дои:10.1007/978-3-642-20992-5_6. ISBN 978-3-642-20992-5. Получено 23 ноября 2013.
Огромные усилия по детальному изучению генома человека привели к получению огромного количества генетических данных. Хотя нам все еще не удается понять молекулярные основы большинства сложных признаков, включая многие распространенные заболевания, теперь у нас есть более четкое представление о степени генетического сходства между людьми и другими видами приматов. Мы также знаем, что люди генетически очень близки друг к другу, на самом деле больше, чем любые другие приматы, что большая часть нашего генетического разнообразия объясняется индивидуальными различиями внутри популяций, и что только небольшая часть генетической дисперсии видов приходится на разные популяции. и их географические группы.
- ^ Рамачандран, Сохини; Тан, Хуа; Gutenkunst, Ryan N .; Бустаманте, Карлос Д. (2010). «Глава 20: Генетика и геномика структуры человеческой популяции» (PDF). В Speicher, Michael R .; Антонаракис, Стилианос Э .; Мотульский, Арно Г. (ред.). Генетика человека Фогеля и Мотульского: проблемы и подходы. Гейдельберг: Springer Scientific. С. 589–615. Дои:10.1007/978-3-540-37654-5. ISBN 978-3-540-37653-8. Архивировано из оригинал (PDF) 3 декабря 2013 г.. Получено 29 октября 2013. Сложить резюме (4 сентября 2010 г.).
Большинство исследований популяционной генетики человека начинается со ссылки на основополагающую статью Ричарда Левонтина 1972 года, названную этим подразделом [29]. Учитывая центральную роль, которую эта работа сыграла в нашей области, мы начнем с ее краткого обсуждения и вернемся к ее выводам на протяжении всей главы. ... Ключевой вывод статьи состоит в том, что 85,4% от общей наблюдаемой генетической изменчивости произошло внутри каждой группы. То есть он сообщил, что подавляющее большинство генетических различий обнаруживается внутри популяций, а не между ними. ... Его открытие воспроизводилось в исследовании за исследованием вплоть до настоящего времени: два случайных человека из любой одной группы (которая может быть континентом или даже местной популяцией) почти так же различаются, как любые два случайных человека со всего мира.
дальнейшее чтение
- Баккер, Винфридус (2006). «Повышенная энергия гибрида приносит пользу селекционерам и бройлерам» (PDF). Кобб Фокус (2). Архивировано из оригинал (PDF) 17 декабря 2008 г.
- Бирчлер Дж. А., Оже Д. Л., Риддл, Северная Каролина (октябрь 2003 г.). «В поисках молекулярной основы гетерозиса». Растительная клетка. 15 (10): 2236–9. Дои:10.1105 / tpc.151030. ЧВК 540269. PMID 14523245.
- Техническая записка NOAA NMFS NWFSC-30: Генетические эффекты заблудших инкубаторных рыб в естественные популяции: депрессия инбридинга и депрессия аутбридинга
- Мак Грегор, С. Э. (2009) [1976]. «Введение: Гибридная сила растений и ее связь с опылением насекомыми» (PDF). Опыление насекомыми культурных растений. Справочник по сельскому хозяйству № 496. Служба сельскохозяйственных исследований, США. OCLC 243509268.
- «Гибриды и фамильные реликвии» - статья из издательства Иллинойсского университета. Подсказки Home Hort
- Ройбал, Дж. (1 июля 1998 г.). "Ranchstar ". Говядина (beefmagazine.com).
- "Секс-ссылки" —В отношении домашней птицы; на FeatherSite