Островная карликовость - Википедия - Insular dwarfism

Островная карликовость, форма филетическая карликовость,[1] это процесс и состояние крупных животных, которые развиваются или имеют уменьшенный размер тела[а] когда ареал их популяции ограничен небольшой средой, в основном островами. Этот естественный процесс отличается от намеренного создания карликовых пород, называемых карликовый. Этот процесс происходил много раз на протяжении эволюционной истории, с примерами, включая динозавры, подобно Европазавр, и современные животные, такие как слоны и их родственники. Этот процесс и другие »островная генетика "артефакты могут встречаться не только на островах, но и в других ситуациях, когда экосистема изолирована от внешних ресурсов и размножения. Это может включать пещеры, пустыня оазисы, изолированные долины и изолированные горы ("небесные острова "). Островная карликовость - один из аспектов более общего "эффект острова" или "правило Фостера", который утверждает, что, когда материковые животные колонизируют острова, мелкие виды, как правило, развивают более крупные тела (островной гигантизм ), а у крупных видов развиваются тела меньшего размера.
Возможные причины
Есть несколько предложенных объяснений механизма, вызывающего такую карликовость.[3][4]
Один из них - это селективный процесс, при котором выживают только мелкие животные, пойманные в ловушку на острове, поскольку пища периодически снижается до пограничного уровня. Более мелкие животные нуждаются в меньших ресурсах и меньших территориях, и поэтому с большей вероятностью преодолеют критическую точку, когда сокращение популяции позволяет источникам пищи пополняться, чтобы выжившие могли процветать. Меньший размер также выгоден с точки зрения репродукции, поскольку влечет за собой более короткие периоды беременности и времена поколения.[3]
В тропиках небольшой размер должен составлять терморегуляция Полегче.[3]
Среди травоядных большой размер дает преимущества в борьбе как с конкурентами, так и с хищниками, поэтому сокращение или отсутствие любого из них будет способствовать карликованию; конкуренция кажется более важным фактором.[4]
Считается, что среди хищников главным фактором является размер и доступность ресурсов добычи, а конкуренция считается менее важной.[4] В тигровые змеи островная карликовость встречается на островах, где доступная добыча ограничена меньшими размерами, чем обычно добывают материковые змеи. Поскольку предпочтение размера жертвы у змей обычно пропорционально размеру тела, маленькие змеи могут быть лучше приспособлены к добыче мелкого размера.[5]
Карликовость против гигантизма
Обратный процесс, при котором мелкие животные, размножающиеся на изолированных островах, лишенных хищников больших массивов суши, могут стать намного крупнее обычного, называется островной гигантизм. Прекрасным примером является додо, предки которых были нормального размера голуби. Также есть несколько видов гигантские крысы, один до сих пор существует, который сосуществовал с обоими Homo floresiensis и карлик стегодонты на Флоресе.
Процесс островного карликования может происходить относительно быстро по эволюционным стандартам. Это контрастирует с увеличением максимального размера тела, которое происходит гораздо более постепенно. При нормировании на длину поколения было обнаружено, что максимальная скорость уменьшения массы тела во время островковой карликовости более чем в 30 раз превышает максимальную скорость увеличения массы тела для десятикратного изменения у млекопитающих.[6] Считается, что это несоответствие отражает тот факт, что педоморфизм предлагает относительно простой путь к уменьшению размеров взрослого тела; с другой стороны, эволюция увеличения максимального размера тела, вероятно, будет прервана появлением ряда ограничений, которые должны быть преодолены эволюционными инновациями, прежде чем процесс может продолжиться.[6]
Факторы, влияющие на степень карликовости
Как для травоядных, так и для плотоядных животных размер острова, степень изолированности острова и размер предковых континентальных видов, по-видимому, не имеют большого прямого значения для степени карликовости.[4] Однако при рассмотрении только массы тела недавних высших травоядных и плотоядных животных и с учетом данных как по континентальным, так и по островным массивам суши было обнаружено, что массы тела самых крупных видов на суше соответствуют размеру суши, причем уклоны примерно 0,5 log (масса тела / кг) на log (площадь суши / км2).[7] Были отдельные линии регрессии за эндотермический высшие хищники, экзотермический высшие хищники, эндотермические высшие травоядные и (на основании ограниченных данных) высшие травоядные с эктотермией, так что потребление пищи было в 7-24 раза выше для высших травоядных, чем для высших хищников, и примерно одинаково для эндотермов и эктотерм тех же трофический уровень (это приводит к тому, что эктотермы в 5–16 раз тяжелее соответствующих эндотермов).[7]
Примеры
Нептичьи динозавры
Признание того, что островная карликовость может относиться к динозаврам, возникло благодаря работе Ференц Нопца, венгерский аристократ, авантюрист, ученый и палеонтолог. Нопса тщательно изучил трансильванских динозавров, заметив, что они меньше своих кузенов в других частях света. Например, он раскопал шестиметровую зауроподы, группа динозавров, которая в других местах обычно вырастала до 30 метров и более. Нопса пришел к выводу, что место, где были обнаружены останки, было островом, Остров Хацег (ныне бассейн Хацег или Хацег в Румыния ) вовремя Мезозойский эпоха.[8][9] Предложение Нопса о карликовости динозавров на острове Хацег сегодня широко принято после того, как дальнейшие исследования подтвердили, что найденные останки принадлежат не молодым особям.[10]
Зауроподы
| Пример | Разновидность | Классифицировать | Временное ограничение | Континентальный родственник |
|---|---|---|---|---|
Ампелозавр | A. atacis | Иберо -Armorican Остров | Поздний мел / Маастрихтский |  Немегтозавриды |
 Европазавр | Э. холджери | Нижняя Саксония | Поздняя юра / Середина Кимериджский |  Брахиозавры |
 Мадьярозавр | М. дакус | Остров Хатег | Поздний мел / маастрихт |  Рапетозавр |
 Лираинозавр[11] | L. astibiae | Иберо-Армориканский остров | Поздний мел | |
 Палудититан | P. nalatzensis | Остров Хатег | Поздний мел / маастрихт |  Эпахтозавр |
Другой
| Пример | Разновидность | Классифицировать | Временное ограничение | Континентальный родственник |
|---|---|---|---|---|
Лангенбергский карьер торвозавр (синий) | Безымянный | Нижняя Саксония | Поздняя юра / Середина Кимериджский | Торвозавр |
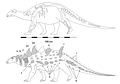 Струтиозавр[12] | S. austriacus S. transylvanicus С. langedocensis | Иберо-Армориканский, Австралоальпийский и Хатегский острова | Поздний мел |  Эдмонтония |
 Телматозавр | Т. transsylvanicus | Остров Хатег | Поздний мел | Гадрозавриды |
 Тетисадрос | Т. insularis | Провинция Триест | Поздний мел | |
 Текодонтозавр[9] | T. antiquus | Южная Англия | Поздний триас / Ретиан |  Платеозавры |
 Залмокс[9] (фиолетовый) | Z. robustus Z. shqiperorum | Остров Хатег | Поздний мел |  Тенонтозавр |
Кроме того, род Балаур изначально был описан как Велоцираптор размер дромеозаврид (и, как следствие, сомнительный пример островной карликовости), но с тех пор была реклассифицирована как вторично нелетающая стволовая птица, более близкая к современным птицам, чем Jeholornis (таким образом, фактически пример островной гигантизм ).
Птицы
| Пример | Биномиальное имя | Родной диапазон | Положение дел | Континентальный родственник | Островной / материк соотношение длины или массы |
|---|---|---|---|---|---|
 Гавайский нелетающий ибисы | Аптерибис Glenos | Молокаи | Вымерший (Поздно Четвертичный ) |  Американские ибисы | |
| Apteribis brevis | Мауи | ||||
| Косумель курасов[13] | Crax rubra griscomi | Косумель | Неизвестный |  Великий курасов | |
 Остров кенгуру эму[14] | Dromaius novaehollandiae baudinianus | Остров Кенгуру, Южная Австралия | Вымерший (ок. 1827 г.) |  Эму | |
 Эму[15] (чернить) | Dromaius novaehollandiae minor | King Island, Тасмания | Вымерший (1822 г. н.э.) | LR ≈ 0,48 [b] | |
 Косумель трэшер[13] | Toxostoma gluttatum | Косумель | Находящихся под угрозой исчезновения |  Другие трэшеры |
Скваматы
| Пример | Биномиальное имя | Родной диапазон | Положение дел | Континентальный родственник | Островной / материк соотношение длины или массы |
|---|---|---|---|---|---|
 Мадагаскарский карликовый хамелеон | Brookesia minima | Нуси-Бе остров, Мадагаскар | Находящихся под угрозой исчезновения |  Мадагаскарские листовые хамелеоны | |
 Нуси-хара-хамелеон[16] | Brookesia Micra | Нуси Хара остров, Мадагаскар | Уязвимый | ||
| Тигровая змея с острова Роксби[5] | Notechis scutatus | Остров Роксби, Южная Австралия | Неизвестный |  Тигровая змея | |
| Карликовый бирманский питон | Python bivittatus progschai | Ява, Бали, Сумбава и Сулавеси, Индонезия | Неизвестный |  Бирманский питон | LR ≈ 0,44 [c] |
| Сетчатый питон Танахьямпеа[19] | Питон ретикулатус jampeanus | Танахьямпеа, между Сулавеси и Флоресом | Неизвестный |  Сетчатый питон | LR ≈ 0,41, самцы LR ≈ 0,49, самки [d] |
Млекопитающие
Пилосаны
| Пример | Биномиальное имя | Родной диапазон | Положение дел | Континентальный родственник |
|---|---|---|---|---|
 Карликовый трехпалый ленивец | Bradypus pygmaeus | Исла-Эскудо-де-Верагуас, Панама | Находящихся под угрозой исчезновения |  Ленивец с коричневым горлом |
 Акратокнус | A. antillensis А. odontrigonus A. ye | Куба, Hispaniola и Пуэрто-Рико | Вымерший (ок. 3000 г. до н.э.) |  Континентальный наземные ленивцы |
| Имагокнус | I. zazae | Куба | Вымерший (Ранний миоцен ) | |
 Мегалокнус | М. роденс М. зиле | Куба и Hispaniola | Вымерший (ок. 2700 г. до н.э.) | |
 Neocnus | Neocnus виды | Куба и Hispaniola | Вымерший (ок. 3000 г. до н.э.) |
Хоботки
| Пример | Биномиальное имя | Родной диапазон | Положение дел | Континентальный родственник |
|---|---|---|---|---|
| Карликовый слон Сулавеси | Elephas Celebensis | Сулавеси | Вымерший (Ранний плейстоцен) |  Азиатский слон |
 Критский карликовый мамонт | Mammuthus creticus | Крит | Вымерший | Маммутус |
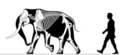 Мамонт Нормандских островов | Mammuthus exilis | Санта-Розае остров | Вымерший (Поздний плейстоцен ) | Колумбийский мамонт |
| Сардинский мамонт | Mammuthus lamarmorai | Сардиния | Вымерший (Поздний плейстоцен) |  Степной мамонт |
| Шерстистый мамонт острова Святого Павла[22][23] | Mammuthus primigenius | Остров Святого Павла, Аляска | Вымерший (ок. 3750 г. до н.э.) | Шерстистый мамонт |
 Сикуло-мальтийские слоны | Palaeoloxodon antiquus leonardi П. mnaidriensis P. melitensis P. falconeri | Сицилия и Мальта | Вымерший |  Слон с прямыми бивнями (оставили) |
| Критские слоны | Palaeoloxodon chaniensis P. creutzburgi | Крит | Вымерший | |
 Кипрский карликовый слон | Палеолоксодон киприоты | Кипр | Вымерший (ок. 9000 г. до н.э.) | |
| Карликовый слон Наксоса | Палеолоксодон sp. | Наксос | Вымерший | |
| Карликовый слон Родоса и Тилоса | Палеолоксодон тилиенсис | Родос и Тилос | Вымерший | |
| Бумиаю карликовый синомастодонт[24] | Sinomastodon bumiajuensis | Остров Бумиаю (ныне часть Ява ) | Вымерший (Ранний плейстоцен) |  Sinomastodon |
 Японский стегодонт[25] | Stegodon aurorae | Япония и Тайвань[26] | Вымерший (Ранний плейстоцен ) |  Китайский Стегодон |
| Карликовый стгодонт Большого Флореса[3] | Stegodon florensis | Флорес | Вымерший (Поздний плейстоцен) |  Сундаленд Стегодон |
| Яванские карликовые стегодонты | Стегодон гипсилофус[24] С. семедоенсис[27] С. sp.[24] | Ява | Вымерший (Четвертичный ) | |
| Минданао карликовый стгодонт[28] | Стегодон mindanensis | Минданао и Сулавеси | Вымерший (Средний плейстоцен) | |
| Сулавесский карликовый стгодонт[24] | Stegodon sompoensis | Сулавеси | Вымерший | |
| Карликовый стгодонт Малый Флорес[3] | Stegodon sondaari | Флорес | Вымерший (Средний плейстоцен) | |
| Сумба карликовый стгодонт[29] | Stegodon sumbaensis | Сумба, Индонезия | Вымерший (Средний плейстоцен) | |
| Тимор карликовый стгодонт[24] | Стгодон тиморский | Тимор | Вымерший | |
| Карликовый стеголофодонт[30] | Стеголофодон псевдолатиденс | Япония | Вымерший (Миоцен ) |  Стеголофодон |
Приматы
| Пример | Биномиальное имя | Родной диапазон | Положение дел | Континентальный родственник |
|---|---|---|---|---|
| Карликовый лемур Нуси Хара[31] | Cheirogaleus sp. | Нуси Хара остров у Мадагаскара | Неизвестный |  Карликовые лемуры |
 Флорес Мэн[32] | Homo floresiensis | Флорес | Вымерший (Поздний плейстоцен) |  человек прямоходящий |
 Мужчина Кальяо | Homo luzonensis[33][34] | Лусон, Филиппины | Вымерший (Поздний плейстоцен) | |
| Современные пигмеи Флореса[35] | Homo sapiens | Флорес | Сохранившийся |  Homo sapiens |
| Ранние современные люди Палау (оспариваются)[36] | Homo sapiens | Палау | Вымерший (?) | |
 Андаманский[37][38] | Homo sapiens | Андаманские острова | Сохранившийся | |
 Сардинская макака[39] | Macaca majori | Сардиния | Вымерший (Плейстоцен) |  Берберийская макака |
 Занзибарский красный колобус | Piliocolobus kirkii | Унгуджа | Находящихся под угрозой исчезновения |  Удзунгва красный колобус |
Плотоядные
| Пример | Биномиальное имя | Родной диапазон | Положение дел | Континентальный родственник | Островной / материк соотношение длины или массы |
|---|---|---|---|---|---|
 Японский волк | Canis lupus hodophilax | Япония (без учета Хоккайдо ) | Вымерший (1905 г. н.э.) |  серый волк | |
 Сардинский волк (вперед) | Цинотерий сардус | Корсика и Сардиния | Вымерший (ок. 8300 г. до н.э.) | 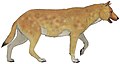 Ксеноцион | |
| Тринил собака | Mececyon trinilensis | Ява | Вымерший (Плейстоцен) | ||
| Коати острова Косумель[13] | Насуа Нарика Нельсони | Косумель | Находящихся под угрозой исчезновения |  Юкатан белоносое пальто | |
 Занзибарский леопард | Panthera pardus pardus | Унгуджа | Находящиеся под угрозой исчезновения или вымершие |  Африканский леопард | |
 Балийский тигр | Panthera tigris sondaica | Бали | Вымерший (ок. 1940 г.) |  Суматранский тигр | |
 Яванский тигр | Ява | Вымерший (ок. 1975 г.) | |||
 Косумельский енот | Procyon pygmaeus | Косумель | Находящихся под угрозой исчезновения |  Обыкновенный енот | |
 Островная лиса | Urocyon littoralis | Шесть из Нормандские острова Калифорнии | Под угрозой |  Серая лиса | LR ≈ 0,84 [e] LR ≈ 0,75 [f] |
| Косумельская лиса | Уроцион sp. | Косумель | Находящиеся под угрозой исчезновения или вымершие |
Нежвачные животные копытные
| Пример | Биномиальное имя | Родной диапазон | Положение дел | Континентальный родственник |
|---|---|---|---|---|
 Малагасийские карликовые бегемоты | Гиппопотам лалумена H. lemerlei H. madagascariensis | Мадагаскар | Вымерший (ок. 1000 г. н.э.) |  Обыкновенный бегемот |
| Бумиаю карликовый бегемот[24] | Гексапротодон симплекс | Остров Бумиаю (ныне Ява ) | Вымерший (Ранний плейстоцен ) |  Азиатские бегемоты |
 Критский карликовый бегемот | Бегемот Creutzburgi | Крит | Вымерший (Средний плейстоцен ) |  Европейский бегемот |
 Мальтийский карликовый бегемот | Гиппопотам мелитенсис | Мальта | Вымерший (Плейстоцен) | |
 Кипрский карликовый бегемот | Малый бегемот | Кипр | Вымерший (ок. 8000 г. до н.э.) | |
 Сицилийский карликовый бегемот | Бегемот пентланди | Сицилия | Вымерший (Плейстоцен) | |
| Косумель ошейник пекари[13] | Pecari tajacu nanus | Косумель | Неизвестный |  Пекари с ошейником |
| Филиппинский носорог[42] | Филиппинский носорог | Лусон | Вымерший (Средний плейстоцен) |  Яванский носорог |
Bovids
| Пример | Биномиальное имя | Родной диапазон | Положение дел | Континентальный родственник |
|---|---|---|---|---|
| Сицилийский бизон[25] | Бизон priscus siciliae | Сицилия | Вымерший (Поздний плейстоцен) |  Степной зубр |
| Сицилийские зубры[43] | Bos primigenius siciliae[25] | Сицилия | Вымерший (Поздний плейстоцен) |  Евразийские зубры |
| Себу тамарау | Bubalus cebuensis | Себу, Филиппины | Вымерший |  Дикий буйвол |
 Низменные аноа | Bubalus depressicornis | Сулавеси и Но на, Индонезия | Находящихся под угрозой исчезновения | |
 Тамарау | Bubalus mindorensis | Миндоро, Филиппины | Находящихся под угрозой исчезновения | |
 Горное аноа | Bubalus Quarlesi | Сулавеси и Но на, Индонезия | Находящихся под угрозой исчезновения | |
 Пещерный козел Балеарских островов | Myotragus balearicus | Майорка и Менорка | Вымерший (после 3000 г. до н.э.) | Галлогорал |
| Несогорал[44] | Несогорал виды | Сардиния | Вымерший | |
| Дахлак Кебир газель[45] | Nanger Soemmerringi ssp. | Дахлак Кебир остров, Эритрея | Уязвимый |  Газель Земмерринга |
Cervids и родственники
| Пример | Биномиальное имя | Родной диапазон | Положение дел | Континентальный родственник |
|---|---|---|---|---|
 Критские карликовые мегацерины[грамм] | Candiacervus виды | Крит | Вымерший (Плейстоцен) |  Praemegaceros вертикорнис[9] |
 Сардинский мегацерин[9] (второй слева) | Praemegaceros cazioti | Сардиния | Вымерший (ок. 5500 г. до н.э.) | |
 Рюкю карликовый олень[48] | Цервус астилодон | Острова Рюкю | Вымерший |  Пятнистый олень (?) Cervus praenipponicus (?) |
| Популяция благородного оленя Джерси[49] | Cervus elaphus jerseyensis | Джерси | Вымерший (Плейстоцен) |  Красный олень |
 Корсиканский благородный олень | Cervus elaphus corsicanus | Корсика и Сардиния | Под угрозой | |
| Плейстоценовый сицилийский олень[25] | Cervus siciliae | Сицилия | Вымерший (Поздний плейстоцен) | |
 Гоплитомерикс[час] | Гоплитомерикс виды | Остров Гаргано | Вымерший (Ранний плиоцен ) |  Pecorans |
| Сицилийский мегацерин[25] | Megaloceros carburangelensis | Сицилия | Вымерший (Поздний плейстоцен) |  Ирландский лось |
 Флорида ключевой олень | Odocoileus virginianus clavium | Флорида-Кис | Находящихся под угрозой исчезновения |  Вирджиния олень |
 Шпицберген северный олень | Rangifer tarandus platyrhynchus | Свальбард | Неизвестный |  Северный олень |
 Филиппинский олень | Руса Марианна | Филиппины | Уязвимый |  Самбарский олень |
Растения
| Возможный пример | Биномиальное имя | Родной диапазон | Положение дел | Континентальный родственник |
|---|---|---|---|---|
 Островные кактусы слона[50][51] | Pachycereus pringlei | Удаленные острова в Море Кортеса (например. Санта Круз, Сан-Педро-Мартир ) | Не оценивается |  Материковый слон кактусы |
Смотрите также
- Островной гигантизм
- Правило Фостера
- Карликовость
- Франц Нопца
- Островная покорность
- Плейстоценовые вымирания
Примечания
- ^ Примером неостровного филетического карликовизма является эволюция карликовости. мартышки и тамарины среди обезьян Нового Света, кульминацией которого стало появление самого маленького экземпляра, Cebuella pygmaea.[2]
- ^ Исходя из высот Heupink на рис. и другие., 2011[15]
- ^ На основе максимальной длины 2,5 м для карликовой формы[17] и 5,74 м для материковой формы[18]
- ^ На основе максимальной общей длины (TL) питона Tanahjampea 2,10 м для самцов и 3,35 м для самок.[19] и максимальная длина рыла южного суматринского питона до длины ствола (SVL) 4,5 м для самцов и 6,1 м для самок.[20] с SVL, скорректированными до TL путем умножения на коэффициент 1,127, полученного из средней относительной длины хвоста (0,113) африканских и индийских питонов.[21]
- ^ Для близлежащих континентальных серых лисиц[40]
- ^ Для континентальных серых лисиц в целом[41]
- ^ Нравиться Гоплитомерикс, Candiacervus Представляется необычным случаем, когда представители этого рода эволюционировали в островные виды самых разных размеров, причем не только карликовые, но и некоторые, которые можно было бы считать гигантами.[46][47]
- ^ Гоплитомерикс очевидно, довольно необычный случай, потому что представители этого рода, по-видимому, эволюционировали как в карликовые, так и в гигантские островные формы на одном и том же острове (ах).[46]
Рекомендации
- ^ Prothero, D. R .; Серено, П. К. (зима 1982 г.). «Аллометрия и палеоэкология карликовых носорогов среднего миоцена прибрежной равнины Техасского залива». Палеобиология. 8 (1): 16–30. Дои:10.1017 / S0094837300004322. JSTOR 2400564.
- ^ Перельман, П .; и другие. (2011). «Молекулярная филогения живых приматов». PLOS Genetics. 7 (3): 1–17. Дои:10.1371 / journal.pgen.1001342. ЧВК 3060065. PMID 21436896.
- ^ а б c d е Van Den Bergh, G.D .; Rokhus Due Awe; Морвуд, М. Дж .; Сутикна, Т .; Ятмико; Вахю Саптомо, Э. (май 2008 г.). "Самый молодой Стегодон остатки в Юго-Восточной Азии из археологического памятника позднего плейстоцена Лян-Буа, Флорес, Индонезия ". Четвертичный международный. 182 (1): 16–48. Bibcode:2008QuInt.182 ... 16V. Дои:10.1016 / j.quaint.2007.02.001.
- ^ а б c d Raia, P .; Мейри, С. (август 2006 г.). «Островное правило у крупных млекопитающих: палеонтология встречается с экологией». Эволюция. 60 (8): 1731–1742. Дои:10.1111 / j.0014-3820.2006.tb00516.x. PMID 17017072. S2CID 26853128.
- ^ а б Keogh, J. S .; Scott, I. A. W .; Хейс, К. (январь 2005 г.). «Быстрое и повторяющееся происхождение островного гигантизма и карликовости у австралийских тигровых змей». Эволюция. 59 (1): 226–233. Дои:10.1111 / j.0014-3820.2005.tb00909.x. PMID 15792242. S2CID 58524.
- ^ а б Evans, A.R .; и другие. (30.01.2012). «Максимальная скорость эволюции млекопитающих». PNAS. 109 (11): 4187–4190. Bibcode:2012PNAS..109.4187E. Дои:10.1073 / pnas.1120774109. ЧВК 3306709. PMID 22308461. Получено 2011-02-11.
- ^ а б Burness, G.P .; Даймонд, Дж.; Фланнери, Т. (2001-12-04). «Динозавры, драконы и карлики: эволюция максимального размера тела». Труды Национальной академии наук. 98 (25): 14518–14523. Bibcode:2001PNAS ... 9814518B. Дои:10.1073 / pnas.251548698. ISSN 0027-8424. JSTOR 3057309. ЧВК 64714. PMID 11724953.
- ^ «Остров карликовых динозавров действительно существовал, утверждают ученые». Телеграф Медиа Группа. 2010-02-22. Получено 2010-02-26.
- ^ а б c d е Бентон, М. Дж .; Csiki, Z .; Григореску, Д .; Redelstorff, R .; Sander, P.M .; Stein, K .; Вейшампель, Д. Б. (28 января 2010 г.). «Динозавры и правление острова: карликовые динозавры с острова Хацег» (PDF). Палеогеография, палеоклиматология, палеоэкология. 293 (3–4): 438–454. Bibcode:2010ППП ... 293..438Б. Дои:10.1016 / j.palaeo.2010.01.026. Архивировано из оригинал (PDF) на 2011-07-10. Получено 2017-07-30.
- ^ Дайк, Г. (2011-09-20). "Барон динозавров Трансильвании". Scientific American. 305 (4): 80–83. Bibcode:2011SciAm.305c..80D. Дои:10.1038 / scientificamerican1011-80. PMID 22106812.
- ^ Компания, Дж. (2010). «Гистология костей титанозавра. Лираинозавр астибия (Dinosauria: Sauropoda) из позднего мела Испании ». Naturwissenschaften. 98 (1): 67–78. Дои:10.1007 / s00114-010-0742-3. HDL:10251/148874. PMID 21120450. S2CID 31752413.
- ^ Карпентер, К. (2001) Бронированные динозавры. Издательство Индианского университета, 526 страниц.
- ^ а б c d Cuarón, A.D .; Мартинес-Моралес, М. А .; McFadden, K. W .; Валенсуэла, Д .; Гомппер, М. Э. (2004). «Статус карликовых хищников на острове Косумель, Мексика». Биоразнообразие и сохранение. 13 (2): 317–331. CiteSeerX 10.1.1.511.2040. Дои:10.1023 / b: bioc.0000006501.80472.cc. S2CID 25730672.
- ^ Паркер С. (1984) Вымерший эму острова Кенгуру, до сих пор неизвестный вид. Бюллетень Британского клуба орнитологов 104: 19–22.
- ^ а б Heupink, T. H .; Huynen, L .; Ламберт, Д. М. (2011). «Древняя ДНК указывает на то, что дварф и« гигантский »эму являются идентичными». PLoS ONE. 6 (4): e18728. Bibcode:2011PLoSO ... 618728H. Дои:10.1371 / journal.pone.0018728. ЧВК 3073985. PMID 21494561.
- ^ Glaw, F .; Köhler, J .; Townsend, T. M .; Венцес, М. (2012-02-14). «Соперничество с самыми маленькими рептилиями в мире: открытие миниатюрных и микроэндемичных новых видов листовых хамелеонов (Брукезия) из Северного Мадагаскара ". PLoS ONE. 7 (2): e31314. Bibcode:2012PLoSO ... 731314G. Дои:10.1371 / journal.pone.0031314. ЧВК 3279364. PMID 22348069.
- ^ де Ланг Р., Фогель Г. (2005). Змеи Сулавеси: Полевой справочник по наземным змеям Сулавеси с идентификационными ключами. Вклад Франкфурта в группу 25 естественной истории, издание Chimaira 2005. ISBN 3-930612-85-2. С. 23-27, 198-201.
- ^ Barker, D.G .; Barten, S.L .; Ehrsam, J.P .; Даддоно, Л. (2012). "Скорректированная длина двух хорошо известных гигантских питонов и установление нового рекорда максимальной длины для бирманских питонов", Питон бивиттатус" (PDF). Бюллетень Чикагского герпетологического общества. 47 (1): 1–6. Получено 2020-03-02.
- ^ а б Аулия, М .; Mausfeld, P .; Schmitz, A .; Бёме, В. (9 апреля 2002 г.). "Обзор сетчатого питона (Питон ретикулатус Schneider, 1801) с описанием нового подвида из Индонезии ». Naturwissenschaften. 89 (5): 201–213. Bibcode:2002NW ..... 89..201A. Дои:10.1007 / s00114-002-0320-4. PMID 12135085. S2CID 4368895.
- ^ Shine, R .; Harlow, P.S .; Keogh, J.S .; Боэди, Н. (1998). "Влияние пола и размера тела на пищевые привычки гигантской тропической змеи, Питон ретикулатус ". Функциональная экология. 12 (2): 248–258. Дои:10.1046 / j.1365-2435.1998.00179.x.
- ^ Sheehy, C.M .; Альберт, J.S .; Lillywhite, H.B .; Ван Дамм, Р. (2016). «Эволюция длины хвоста у змей, связанная с различной гравитационной средой». Функциональная экология. 30 (2): 244–254. Дои:10.1111/1365-2435.12472.; см. таблицу S1
- ^ Ширбер, Майкл. Surviving Extinction: Где жили шерстистые мамонты. Живая наука. Imaginova Corporation. Проверено 20 июля 2007.
- ^ Мамонты Остров Врангеля, к северу от Сибирь, больше не считаются карликами. См .: Тихонов Алексей; Ларри Агенброуд; Сергей Вартанян (2003). Сравнительный анализ популяций мамонтов на острове Врангеля и Нормандских островах. DEINSEA 9: 415–420. ISSN 0923-9308
- ^ а б c d е ж Азиз, Ф .; ван ден Берг, Г. Д. (25 сентября 1995 г.). "Карлик Стегодон из Самбунгмакана (Центральная Ява, Индонезия) ". Proc. Кон. Нед. Акад. В. Ветенш. 98 (3): 229–241. Получено 2017-07-31.
- ^ а б c d е Sondaar, P. Y .; A.A.E. ван дер Гир (2005). «Эволюция и исчезновение плиоплейстоценовых островных копытных». Международный журнал Французской четвертичной ассоциации. 2: 241–256. Получено 2017-07-31.
- ^ http://www.rhinoresourcecenter.com/pdf_files/129/1291330178.pdf
- ^ Сисванто, С., & Ноервиди, С. (2014). ИСПОЛЬЗОВАНИЕ ПРОБОСИДЕИ С САЙТА СЕМЕДО: его корреляция с биостратиграфией и прибытием человека на Яву. Беркала Аркеологи, 34 (2).
- ^ Заим Ю. (20 августа 2010 г.). «Геологические свидетельства самого раннего появления гомининов в Индонезии». В Fleagle, J. G; Shea, J. J .; Grine, F.E .; Baden, A. L .; Лики, Р. Э. (ред.). Из Африки I: первая гомининская колонизация Евразии. Springer Science & Business Media. п. 106. ISBN 978-90-481-9036-2. OCLC 668096676.
- ^ http://ro.uow.edu.au/cgi/viewcontent.cgi?article=3055&context=smhpapers
- ^ Саегуса, Х. (2008). "Карлик Стеголофодон из миоцена Японии: Пассажиры на тонущих лодках ". Четвертичный международный. 182 (1): 49–62. Bibcode:2008QuInt.182 ... 49S. Дои:10.1016 / j.quaint.2007.08.001.
- ^ «Новая группа карликовых лемуров может быть самым редким приматом в мире».
- ^ Ученый изучит морфинг Хоббита, abc.net.au
- ^ Уэйд, Л. (10 апреля 2019 г.). «Новый вид древнего человека, обнаруженный на Филиппинах». Наука. 364. Дои:10.1126 / science.aax6501.
- ^ Détroit, F .; Mijares, A. S .; Корни, Дж .; Daver, G .; Zanolli, C .; Дизон, Э .; Robles, E .; Grün, R .; Пайпер, П. Дж. (2019). "Новый вид Гомо из позднего плейстоцена Филиппин ". Природа. 568 (7751): 181–186. Bibcode:2019Натура.568..181Д. Дои:10.1038 / s41586-019-1067-9. PMID 30971845. S2CID 106411053.
- ^ Tucci, S .; и другие. (2018-08-03). «Эволюционная история и адаптация человеческой популяции пигмеев острова Флорес, Индонезия». Наука. 361 (6401): 511–516. Bibcode:2018Научный ... 361..511T. Дои:10.1126 / science.aar8486. ЧВК 6709593. PMID 30072539.
- ^ "Исследование гласит, что древние малые люди на Палау не гномы ". National Geographic News. 27 августа 2008 г.
- ^ Гиббонс, А. (2018). «Жизнь на острове может уменьшить человека». Наука. Дои:10.1126 / science.aau9750.
- ^ Мондаль, М .; Casals, F .; Xu, T .; Dall'Olio, G.M .; Pybus, M .; Netea, M. G .; Comas, D .; Laayouni, H .; Li, Q .; Majumder, P.P .; Бертранпетит, Дж. (2016). «Геномный анализ андаманцев дает представление о древней миграции людей в Азию и адаптации» (PDF). Природа Генетика. 48 (9): 1066–1070. Дои:10,1038 / нг.3621. HDL:10230/34401. PMID 27455350. S2CID 205352099.
- ^ Ладья, Л. (31 декабря 2008 г.). «Первый семинар по летописи европейских окаменелостей приматов (Сиена и Гроссето, 11-13 сентября 2008 г.) с обновленной информацией об итальянских исследованиях в палеоприматологии» (PDF). Атти Мусс. Stor. Nat. Маремма (22): 129–143.
- ^ Парфит, М .; Гроо, М. (22 апреля 2020 г.). «Воодушевляющая история об этих крошечных островных лисицах, почти уничтоженных катастрофой». NationalGeographic.com. Национальная география. Получено 2020-04-23.
- ^ Moore, C.M .; Коллинз, П.В. (1995). «Виды млекопитающих - Urocyon littoralis" (PDF). 489: 1–7. Архивировано из оригинал (PDF) 22 января 2012 г.. Получено 16 сентября 2011. Цитировать журнал требует
| журнал =(помощь) - ^ Ренема, Виллем (2007). Биогеография, время и место: распространение, барьеры и острова. Springer Science & Business Media. п. 334. ISBN 978-1-4020-6374-9. OCLC 228153573.
- ^ ван Вуур, СНГ (2005). Возвращение зубров: история, морфология и экология вымершего дикого быка. Coronet Books Incorporated. ISBN 978-954-642-235-4. OCLC 472741798.
- ^ van der Geer, A .; Lyras, G; de Vos, J .; Дермитзакис, М. (14 февраля 2011 г.). «Сардиния и Корсика». Эволюция островных млекопитающих: адаптация и вымирание плацентарных млекопитающих на островах. Джон Вили и сыновья. ISBN 978-1-4443-9128-2. OCLC 894698082.
- ^ Chiozzi, G .; Bardelli, G .; Ricci, M .; De Marchi, G .; Кардини, А. (2014). «Еще один островной карлик? Фенотипическая особенность малоизвестной Газели Земмерринга, Nanger Soemmerringii (Cetartiodactyla: Bovidae) с острова Дахлак-Кебир ". Биологический журнал Линнеевского общества. 111 (3): 603–620. Дои:10.1111 / bij.12239.
- ^ а б Mazza, P.P.A .; Росси, M.A .; Агостини, С. (2015). «Hoplitomerycidae (поздний миоцен, Италия), пример гигантизма у островных жвачных животных». Журнал эволюции млекопитающих. 22 (2): 271–277. Дои:10.1007 / s10914-014-9277-2. S2CID 16437411.
- ^ ван дер Гир, A.A.E. (2018). «Однородность разнообразия: морфология и эволюция рогов в среде, свободной от хищников». Palaeontologia Electronica (21.1.9A): 1–31. Дои:10.26879/834.
- ^ Kaifu, Y .; Fujita, M .; Yoneda, M .; Ямасаки, С. (15 февраля 2015 г.). «Плейстоценовое мореплавание и колонизация островов Рюкю, юго-запад Японии». In Kaifu, Y .; Идзухо, М .; Goebel, T .; Sato, H .; Оно, А. (ред.). Возникновение и разнообразие современного человеческого поведения в палеолите Азии. Издательство Техасского университета A&M. ISBN 978-1-62349-277-9. OCLC 985023261.
- ^ Листер, А. М. (1989-11-30). «Быстрое затмение благородных оленей на Джерси в последнее межледниковье». Природа. 342 (6249): 539–542. Bibcode:1989Натура.342..539L. Дои:10.1038 / 342539a0. PMID 2685610. S2CID 4343091.
- ^ Wilder, B.T .; Фельгер, Р. (30 сентября 2010 г.). «Карликовые гиганты, гуано и изоляция: растительность и флористическое разнообразие острова Сан-Педро-Мартир, Калифорнийский залив, Мексика» (PDF). Труды Общества естественной истории Сан-Диего. 42: 1–24, см. Стр. 9–13.. Получено 2020-01-05.
(стр. 12) Уменьшение роста растений Сан-Педро-Мартир, по-видимому, связано с отбором более коротких особей, чтобы выжить в жестоких тропических штормах, возможной корневой конкуренцией в таком густом лесу и неопределенным эффектом высоких уровней азота и фосфора из-за обилие гуано, которое может задержать рост. Генетических исследований не проводилось ...
- ^ Бернс, К. (Май 2019). Эволюция в изоляции: поиск островного синдрома у растений. Издательство Кембриджского университета. С. 174–177. Дои:10.1017/9781108379953. ISBN 978-1108379953. OCLC 1108160200.
(стр. 174-175) ... степень, в которой его карликовый рост генетически детерминирована, и объяснение того, почему островная карликовость может быть избирательно выгодной, требует дополнительных исследований.
внешняя ссылка
| Правила |
|  |
|---|---|---|
| Связанный |
| |